五倍子废渣对小麦生长和产量的影响及施肥对其的缓解效应
2018-04-10冯虹毓胡庭兴胡红玲戴薇薇廖玉华郑永强杨珊珊
冯虹毓,胡庭兴,胡红玲,戴薇薇,廖 蓉,廖玉华,郑永强,杨珊珊,李 琪,3
(1 四川农业大学 林学院,四川 成都 611130;2 厦门大学 环境与生态学院,福建 厦门 361102; 3 中国农业大学 资源与环境学院,北京 100083)
五倍子(Gallachinensis)又名文蛤、百虫仓,是瘿绵蚜科(Pemphigidae)的一些蚜虫如角倍蚜或倍蛋蚜雌虫寄生于漆树科植物盐肤木(RhuschinensisMill.)、青麸杨(RhuspotaniniiMaxim.)、红麸杨(Rhuspunjabensisvar.sinica (Diels) Rehd.et Wils.)及其同属其他植物的嫩叶或叶柄,刺伤植物组织而生成一种囊状聚生物虫瘿,经烘焙干燥后所得,其酿制品为百药煎[1-2]。五倍子广泛用于医药、食品、微电子、环保新材料、有机合成中间体及稀有金属提取、军工试验、表面技术处理等领域。在我国境内,盐肤木、青麸杨主要分布在湖北、湖南、陕西、四川、贵州、云南等省,主要混生于阔叶林之中或呈散生状态分布,这些省的五倍子产量占全国总产量的90%以上[3]。
用五倍子生产单宁酸进而提取没食子酸生产中的废弃物,尚有10%~20%的单宁酸及没食子酸、废弃活性炭和鞣花酸残留,造成资源浪费和环境污染[4]。由于我国没食子酸制备以碱水解法为主,相较于日本采取的发酵法[5-7],废水、废渣及废碳排放量大。但近年来除了对五倍子进行较大规模的加工利用外,有关其产生的废渣在环保型、循环型再利用方面研究很少,限制了倍用林的发展和保护。
我国在中成药生产废渣处理上的研究较多。如刘慧来[8]发现,药渣和化肥配合施用可促进马铃薯(SolanumtuberosumLinn.)的生长发育,使生育期缩短。何佳芳等[9]将夏枯草(PrunellavulgarisLinn.)药渣配制成育苗基质,栽培的小白菜幼苗整齐度好、壮苗指数高。裴毅等[10]探讨了没食子酸对铜绿微囊藻(Microcystisaeruginosa)的抑制效应及超微结构的影响。孙兆法等[11]将多种中药渣混合配制成复合基质,探讨了其对一品红(EuphorbiapulcherrimaWilld.ex Klotzsch.)生长量、开花期、冠幅、花径、苞片数、株高、外观品质等的影响。郑照强等[12]发明的再生活性炭法处理五倍子加工废渣效果较好,但工艺较复杂,对企业的生产和经济能力要求高。那么在企业能力有限的情况下,如果将废渣返回田地,是否会影响作物的生长和产量,是目前亟待探明的问题。
基于此,本研究拟采用盆栽试验,以小麦为受体,利用五倍子废渣模拟还田处理,分析其对农作物生长发育的影响,这对生物资源加工废渣的友好型、环保型再利用具有重要意义和参考价值,对五倍子原料林基地建设、盐肤木的保护与合理利用也具有重要的现实意义。
1 材料与方法
1.1 试验材料
1.1.1供体材料于2015年5月初在四川省峨眉山市相关企业回收五倍子加工废渣,风干捣碎,储藏备用。
1.1.2受体材料小麦品种为蜀麦969,种子由四川农业大学小麦研究所提供,全生育期约185 d[13]。选择饱满、大小均一的种粒,播种前在40 ℃水浴中温汤浸种2.5 h后取出,晾干备用。
1.1.3栽培土壤与容器本试验采用盆栽方式,栽植容器为口径27 cm、高度20 cm的白色聚乙烯塑料盆。栽植土壤为当地的农田土,其理化性质为:pH值7.85,偏碱性,有机质含量24.38 g/kg,全氮含量0.66 g/kg,全磷含量0.60 g/kg,全钾含量5.81 g/kg,碱解氮含量56 mg/kg,速效磷含量128.63 mg/kg,速效钾含量15.27 mg/kg。
1.2 试验方法
1.2.1试验设计本试验采用单因素完全随机试验设计。试验设置0(CK),10,20,30,40,50,60 g/盆 7个五倍子废渣添加水平,分别记为A0,A1,A2,A3,A4,A5和A6,每个水平设置8个重复,共计56盆。将五倍子废渣按照设计的添加量与土壤(8 kg/盆)均匀混合装盆,随后播种小麦(2015-11-02播种),每盆播种30粒并保证其在盆中均匀分布,适当浇水。在出苗后至成熟期间,采用HH2土壤水分速测仪(ML2x,GBR,英国)定期监测土壤含水量并及时浇水,保持土壤湿度在17%左右(约为该试验用土田间持水量的70%)。小麦出苗后,当观察到各处理间形态生长表现出明显差异(如叶片变黄,植株个体大小不同)时,表明添加五倍子废渣对小麦生长产生了影响,故44 d时在各处理中抽出4个重复进行施肥处理(史丹利复合肥:N 24%,P 10%,K 14%),施肥量为7.2 g/盆,分4次施入,每10 d施1次。
1.2.2测定指标及方法分别于播种后第22,44,66和88天采用匀苗方式进行4次取样,对应每盆取幼苗5,4,3,3株,最后每盆保留15株,持续观察生长直至成熟收获。
茎秆地径:用游标卡尺测量植株土壤表面处茎秆宽度(精度0.1 mm)。
株高:用直尺测量植株从土壤表面到叶片顶端的长度(精度0.1 cm)。
地上部分生物量:将植株地上部分在鼓风干燥箱内105 ℃杀青30 min,然后80 ℃烘24 h,测定最终恒定质量(精度0.000 1 g)。
过氧化物酶(POD)活性:采用愈创木酚法[14]测定。
超氧化物歧化酶(SOD)活性:采用NBT光化还原法[15]测定。
过氧化氢酶(CAT)活性:采用紫外分光光度法[14]测定。
可溶性糖(SS)含量:采用硫代巴比妥酸加热显色法[14]测定。
可溶性蛋白(SP)含量:采用考马斯亮蓝G250法[14]测定。
产量及收获指标的测定与计算:每盆单独收获,测定穗粒数、生物产量(地上部分生物量精度0.000 1 g)和籽粒产量(精度0.000 1 g),并计算收获指数(Harvest index,HI),HI=籽粒产量/生物产量[16]。
化感效应指数(Response index,RI)采用Williamson等[17]提出的评判方法,即:RI=1-C/T(当T≥C时)或RI=T/C-1(当T
1.3 数据处理
采用SPSS 22.0统计分析软件(IBM,Inc.,USA)进行单因素方差分析(One-way ANOVA),并用最小显著差数法(LSD法)进行多重比较分析,利用Sigmaplot 10.0(Systat Software,Inc.,USA)和Excel 2010(Microsoft,Inc.,USA)进行相关图表的制作。
2 结果与分析
2.1 五倍子废渣对小麦形态指标及生物量的影响
由表1可知,播种后22~88 d,随着废渣施入量的增加,小麦的株高呈下降趋势。当废渣分解22 d时,各剂量废渣处理对小麦株高的抑制作用总体不显著;44 d时,中剂量(A3、A4,下同)以及高剂量(A5、A6,下同)处理的小麦株高均受到显著抑制(P<0.05);66 d时,中高剂量处理小麦受到的抑制作用逐渐增强;而低剂量废渣(A1、A2,下同)始终与A0差异不显著(P>0.05);88 d时,低剂量废渣对小麦株高的作用由之前的抑制转为促进,而其他处理虽然仍表现为显著抑制,但化感效应指数有所降低。
由表1还可知,在同一测定时间,随着废渣施入量的增加,小麦地径始终呈下降趋势。第1次匀苗(播种后22 d)取样时小麦幼苗比较纤细,难以准确量取茎秆地径,故未量测。播种后44~66 d除A1外,88 d除A1-A3外的其他各处理地茎均显著低于A0(P<0.05)。
由表1可见,通过施肥,当废渣分解到66 d时,小麦株高受到的抑制作用明显降低;88 d时,这种抑制作用进一步降低。地径的变化趋势与株高相似。表明施肥处理可相应缓解五倍子废渣对小麦生长产生的抑制效应,且高剂量处理表现尤为明显。
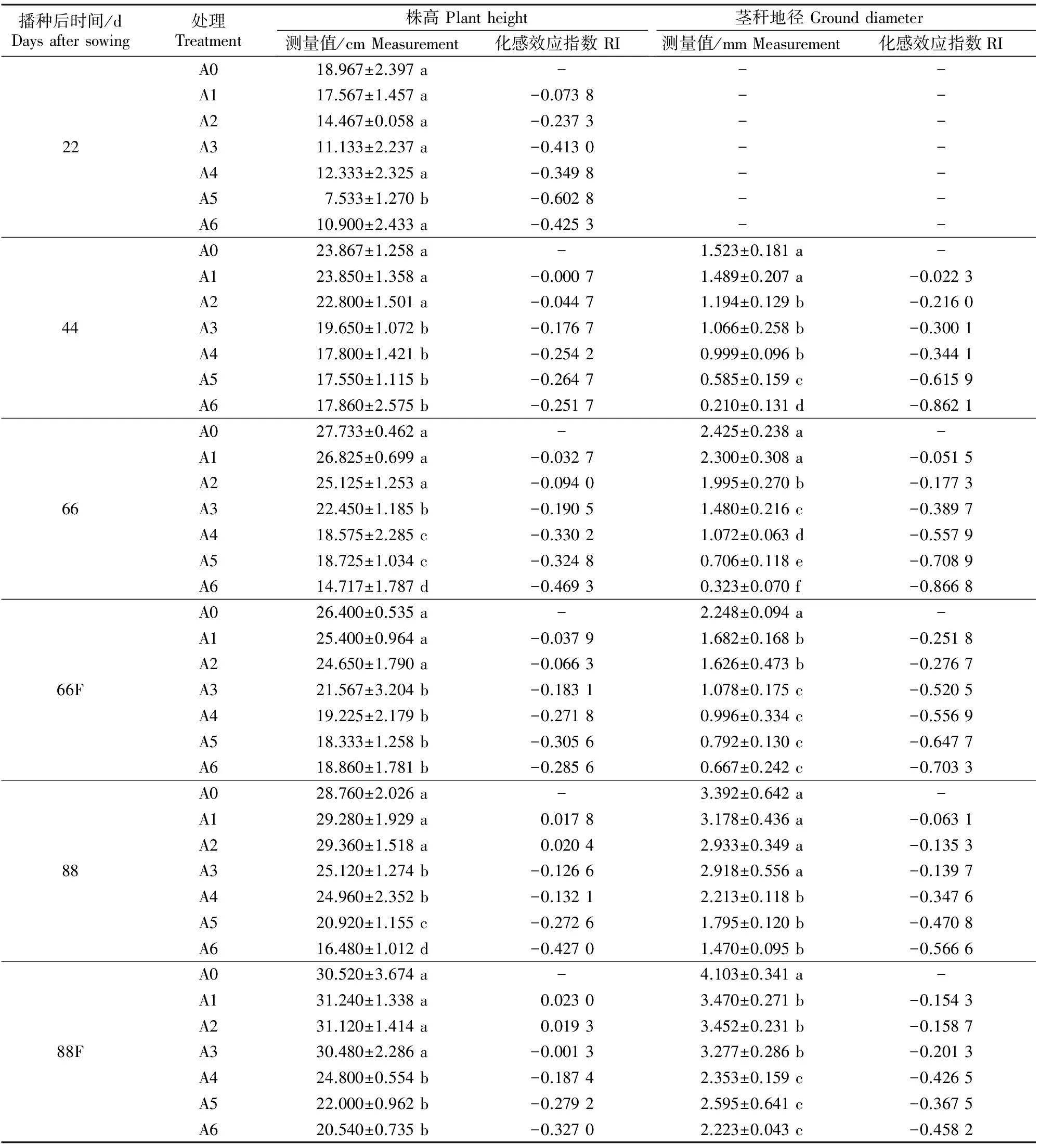
表1 五倍子废渣对小麦株高、茎秆地径的影响及施肥的缓解效应Table 1 Effects of processing residue of Galla chinensis on plant height and ground diameter of wheat and alleviating effect of fertilization
注:同一测定时间同列数据后标不同小写字母表示处理间差异达显著水平(P<0.05);66F和88F分别代表66和88 d时施肥处理。
Note:Different lowercase letters in each column indicate significant difference between treatments at same time (P<0.05);66F and 88F indicate in fertilization group at 66 d and 88 d,respectively.
由图1可知,在同一测定时间,随废渣添加量的增加,小麦地上部分生物量明显降低,且当废渣分解44 d和66 d 时,各处理地上部分生物量始终显著低于A0(P<0.05);88 d时,低剂量处理的地上部分生物量则逐渐接近A0,其他各剂量处理仍显著低于A0处理(P<0.05)。通过施肥,废渣分解66 d时,A3-A6各处理间地上部生物量无显著差异,但仍显著低于A0处理(P<0.05);88 d时,各处理间均无显著差异(P>0.05)。同时由图1还可知,通过施肥处理后,高剂量处理表现出的缓解程度明显高于低、中剂量处理。

图柱上方不同小写字母表示同一测定时间下各处理差异显著(P<0.05);66F和88F分别代表66和88 d时施肥处理。下图同Different lowercase letters on the top of square column and at the same time indicate significant difference at the level of 0.05;66F and 88F indicates fertilization group at 66 d and 88 d,respectively.The same below图1 五倍子废渣对小麦地上部分生物量的影响及施肥的缓解效应Fig.1 Effects of processing residue of Galla chinensis on aboveground biomass of wheat and alleviating effect of fertilization
2.2 五倍子废渣对小麦抗氧化酶活性的影响
五倍子废渣对小麦抗氧化酶活性的影响及施肥的缓解效应见图2。
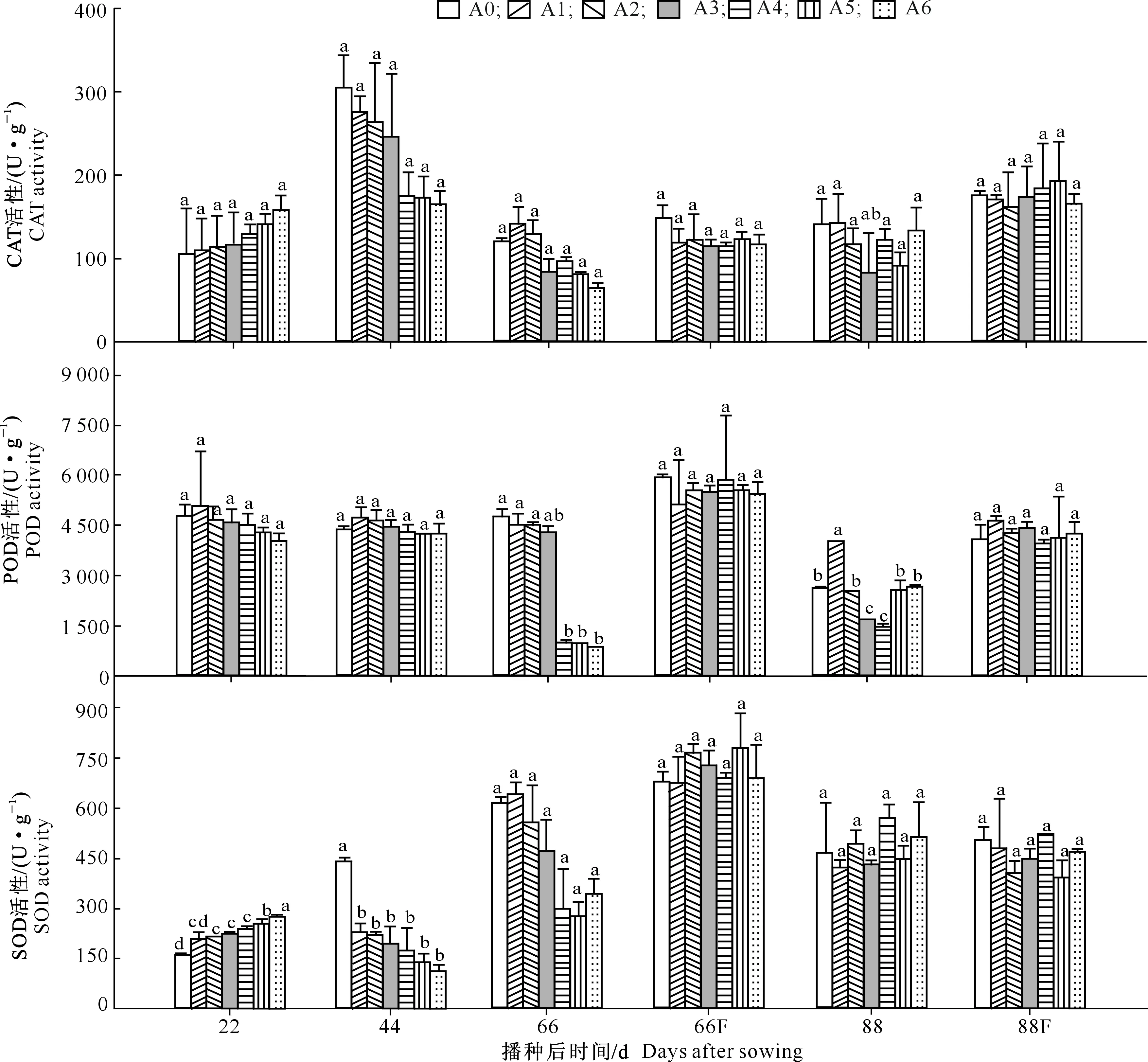
图2 五倍子废渣对小麦抗氧化酶活性的影响及施肥的缓解效应Fig.2 Effects of processing residue of Galla chinensis on antioxidant enzymes activities of wheat and alleviating effect of fertilization
CAT是生物氧化过程一系列抗氧化酶的终端产物[18],能够清除植物在逆境条件下产生的H2O2。由图2可知,当废渣分解44 d 时,小麦叶片CAT活性随着废渣添加量的增加呈现降低趋势;66 d时,其抑制作用虽然明显,但总体上有所降低;88 d时各处理的CAT活性逐渐恢复至正常水平,其抑制作用总体上已不明显。
POD能利用H2O2氧化供氢体,清除过剩的自由基,提高植物的抗逆性。其活性高低能够反映植物的生长发育及体内代谢情况,同时也是植物抗性强弱的标志之一[19-20]。由图2可知,在废渣分解22和44 d时,随着废渣添加量的增加,小麦叶片POD活性逐渐减弱,但处理间差异不显著;66 d时,A4-A6处理废渣对POD活性的抑制作用明显增强,其活性与A0差异达显著水平(P<0.05),而A1~A3的POD活性与A0差异仍不显著;88 d时,各处理间的差异没有明显的规律性。
SOD对于清除氧自由基,防止氧自由基破坏细胞的组成、结构和功能,保护细胞免受氧化损伤具有十分重要的作用。由图2可知,当废渣分解22 d时,随着废渣添加量的增加,小麦叶片的SOD活性逐渐增强,且处理间差异显著(P<0.05);44 d时,随着废渣添加量的增加,各处理的SOD活性呈下降趋势,且与A0间差异显著(P<0.05);66 d时,SOD活性与44 d时的趋势相似,但各处理间无显著差异(P>0.05);88 d时各处理的SOD活性逐渐恢复至正常水平,废渣的抑制作用总体上已不明显。
由图2还可知,通过施肥,五倍子废渣对小麦抗氧化酶活性的影响得到一定的缓解。当废渣分解66和88 d时,CAT、POD和SOD 3种酶活性在各处理间均无显著差异(P>0.05)。结果表明,在小麦幼苗生长过程中,施肥对五倍子废渣的抗氧化酶抑制效应发挥了一定的缓解作用。
2.3 五倍子废渣对小麦渗透调节物质的影响
由图3可知,当废渣分解22 d时,各处理小麦叶片可溶性蛋白含量总体上差异不显著(P>0.05);44 d后,随着废渣添加量的增加,可溶性蛋白含量均呈下降趋势,除A1外的其他处理均与A0差异显著(P<0.05),且表现出明显的梯度式抑制效应;88 d时,各处理的抑制作用减弱,可溶性蛋白含量均接近A0,且无显著性差异(P>0.05)。
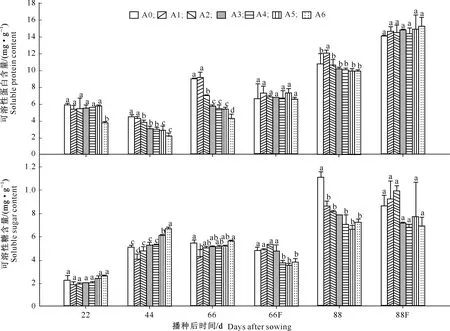
图3 五倍子废渣对小麦渗透调节物质含量的影响及施肥的缓解效应Fig.3 Effects of processing residue of Galla chinensis on osmotic adjustment substance contents of wheat and alleviating effect of fertilization
由图3可知,可溶性糖含量的变化趋势与可溶性蛋白含量基本相反。当废渣分解44 d时,各处理可溶性糖含量与A0的差异逐渐增大(P<0.05),但随着废渣添加量的增加,可溶性糖含量逐渐增大,梯度效应明显;66 d时,各处理的可溶性糖含量总体无显著差异;88 d时,各处理可溶性糖含量均显著低于A0,并随添加量增加呈现下降趋势,但添加废渣处理间无显著差异(P>0.05)。
由图3还可知,经过施肥处理后,当废渣分解66 d时,各处理可溶性蛋白含量无显著差异(P>0.05),但部分中高剂量处理的可溶性糖含量显著低于A0(P<0.05);88 d时,各处理间可溶性糖和可溶性蛋白含量均无显著差异(P>0.05)。结果表明,通过施肥处理后,五倍子废渣对叶片可溶性蛋白和可溶性糖含量的影响得到一定的缓解。
2.4 五倍子废渣对小麦产量及其构成因子的影响
由图4可知,随着废渣添加量的增加,未施肥组小麦的收获指数(HI)呈现出逐渐降低的趋势,A1-A3处理的HI在0.40~0.41,而A4-A6处理的HI在同一收获时期则处于0.34~0.41。小麦的生物产量以及籽粒产量均随着废渣添加量的增加呈逐渐降低的趋势,干物质分配也呈现相同趋势。除A1外,其他各处理较A0均显著降低(P<0.05)。
由图4可知,通过施肥,各处理小麦HI均在0.40~0.55。虽然籽粒产量和生物产量仍随着废渣添加量的增加而逐渐降低,但各处理间的籽粒产量无显著差异(P>0.05);除A6处理外,其他剂量处理间的生物产量也无显著差异。
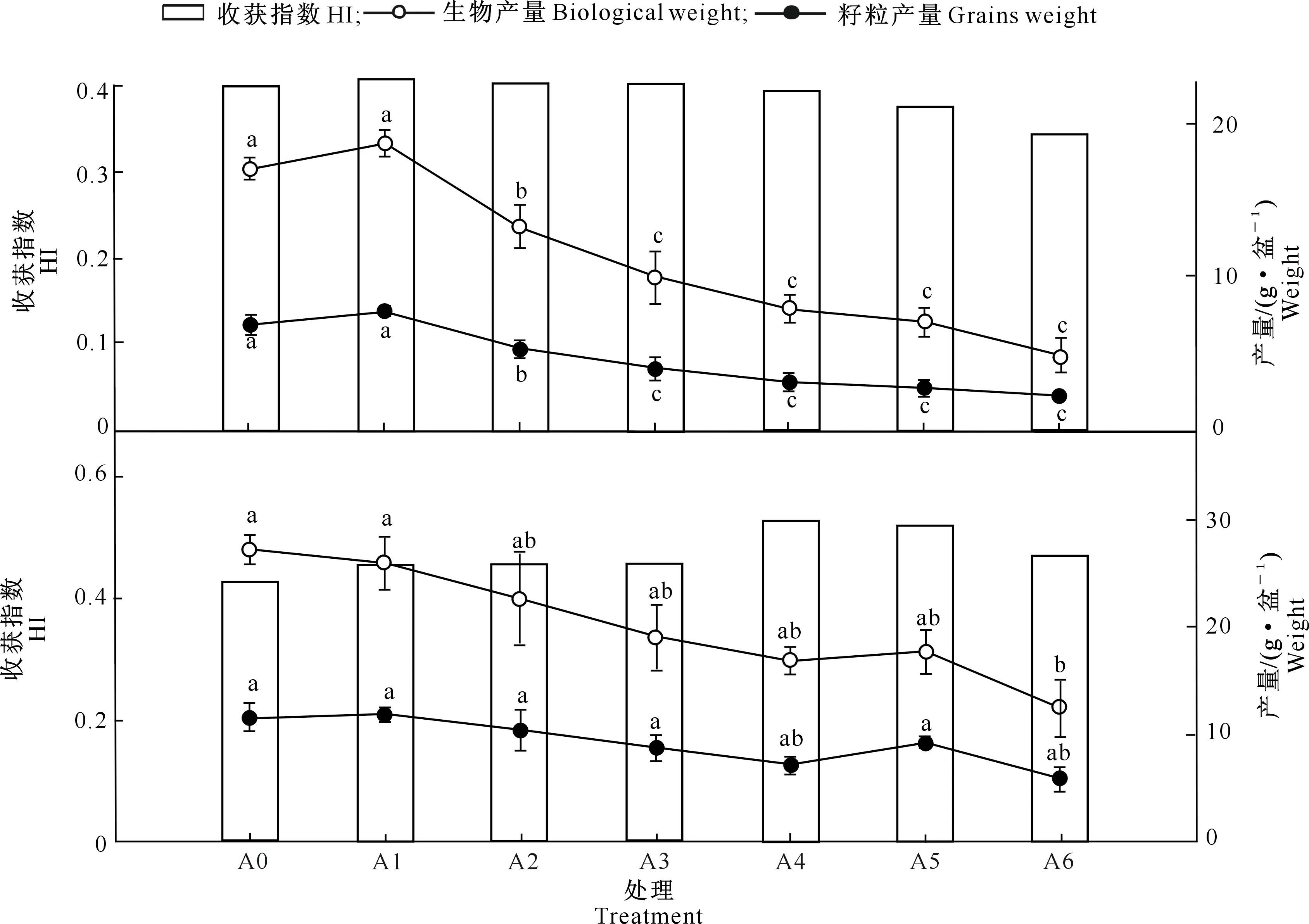
图4 五倍子废渣对未施肥组(上)和施肥组(下)小麦收获质量的影响及施肥的缓解效应Fig.4 Effects of processing residue Galla chinensis on harvest quality of wheat without fertilization group(up),with fertilization group (down) and alleviating effect of fertilization
由图5可知,小麦穗粒数的增减趋势与生物产量和籽粒产量相同,均随着废渣添加量的增加而降低,但各处理间千粒质量降低趋势不明显。这可能是因为在小麦生殖生长过程中,分配给每颗籽粒营养物质的机会是均等的(除穗顶端有几粒在生长后期缺乏养分等原因偏小之外),而废渣分解对这种分配模式的影响并不明显,只是对穗粒数量产生了显著的抑制作用。千粒质量是产量构成中最稳定的因素[21],影响小麦千粒质量的因子有很多,如干旱、物候、氮素过多、小雨等均会导致小麦“走面”现象的发生。通过图5并结合对应的数据统计分析可知,小麦的穗长、穗粒数与收获期的株高存在极显著正相关关系(P<0.01),其相关系数(r)分别为0.981 6和0.946 4;通过图4和图5并结合对应的数据统计分析可知,生物产量、穗粒数、穗长、收获期株高与籽粒产量之间也存在极显著正相关关系(P<0.01),其相关系数(r)分别为0.984 3,0.975 9,0.923 4,0.912 9。这表明在小麦营养生长期只要采取措施减轻废渣的负效应,促进植株个体的生长,就能在一定程度上提高受影响小麦的产量,其中及时施肥则是很有效的措施之一。

图5 五倍子废渣对小麦生殖生长的影响及施肥的缓解效应Fig.5 Effects of processing residue of Galla chinensis on reproductive growth of wheat and alleviating effect of fertilization
由图5可知,虽然小麦的穗长、株高和穗粒数总体上仍随着废渣添加量的增加而降低,但施肥明显缓解了这种降低的趋势。如果将施肥时间提前(比如提前到植株生长表现出受抑制状态初期(表1,A5处理22 d时),其缓解效应可能会更好。另外,各处理小麦的千粒质量均趋于正常水平,其间无显著差异,进一步表明籽粒获得营养物质的机会是均等的。
综合上述结果可知,施氮处理在一定程度上缓解了五倍子废渣对小麦各产量构成因素的抑制效应。
3 讨 论
3.1 五倍子废渣对小麦形态指标及生物量的影响
本研究结果显示,当废渣分解22 d时各处理间的形态指标(株高、地径)总体上无显著差异,而各处理间的地上部分生物量则没有显现出明显的规律性;44和66 d时,各处理间的各项指标均已表现出明显差异;88 d时,低剂量处理的株高超过高剂量处理近60%,地径超过近95%,地上部分生物量则超过413%;且随着废渣施入量的增加,小麦形态指标的抑制效应逐渐增强。这与黄建贝等[22]、杨珊珊等[23]的研究结果相似。究其原因是:一方面施加的五倍子废渣中残留较多的活性炭和木素[24],故可能土壤内碳氮比率随着废渣施入量的增加而增加,导致土壤微生物群体量增加,过高的微生物群体量会出现与植株竞争养分的现象[25],使植株营养生长受到抑制;另一方面废渣的施入会使植株受到化感物质的影响[24],其生长会随着土壤中化感物质的积累而受到一定程度的抑制,影响土壤酶、土壤微生物的代谢以及土壤呼吸,使营养物质不能及时被植物吸收,导致植株不能正常进行营养生长。
3.2 五倍子废渣对小麦抗氧化保护酶系统的影响
氧是一切生物必不可少的生存条件,但是高浓度的氧会引起机体氧中毒如损坏DNA等,氧中毒是由于氧在机体内大量地被转化为活性氧(reactive oxygen species,ROS),造成活性氧过度积累所致[26-28]。同时植物体会应激地启动有效的保护酶系统并对自由基进行清除,形成一定的减缓活性氧伤害的保护机制[20,26]。一般认为,CAT、POD、SOD是植物酶促反应体系的保护酶,它们协同作用,可以防御活性氧对细胞的伤害,在保护自身免受OH-毒害过程中发挥着重要作用[29-30]。
本研究结果表明,随着废渣分解,各处理的CAT和SOD活性均呈先升高后降低的趋势,这与王娟等[31]的研究结果“在长时间胁迫作用下细胞保护酶活性出现一定程度下降”,以及夏民旋等[32]的研究结果“在胁迫条件下SOD活性呈先上升后下降的趋势”相似。而POD活性则与CAT和SOD活性的变化趋势略有不同,这可能与POD对供氢体要求呈广泛性,而SOD和CAT功能则为专一性有关。随着废渣添加量的增加,POD活性呈逐渐降低的趋势,这与张敏等[33]的研究结果“普通小麦POD活性随着盐胁迫浓度的增加而降低”一致。同时高永生等[34]研究发现,POD活性随着盐浓度的升高表现出先升后降的趋势,本研究也得出了相似的结论。
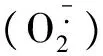
3.3 五倍子废渣对小麦渗透调节物质的影响
渗透调节物质如可溶性糖(SS)和可溶性蛋白(SP)对植物生长具有一定的积极作用。在本研究中,SS含量随着废渣施入量的增加出现先增加后降低的现象,这与王兰兰等[37]发现的“可溶性糖积累量随胁迫程度增加出现先上升后下降的现象”一致,SP含量随着废渣施入量的增加而逐渐降低,仅A1处理的SP含量高于A0,这与王兰兰等[37]、靖姣姣等[38]发现的“小麦在盐胁迫下可溶性蛋白含量高于对照”的结论不一致,可能原因是:(1)胁迫物质与环境不同,废渣的抑制作用导致SS与SP相互转化;(2)废渣的抑制作用使得植物体内渗透调节机制受到干扰,从而降低了植物的渗透调节能力;(3)废渣中的某些物质影响了小麦根系对矿物质元素的吸收,导致蛋白质合成受到一定的抑制作用。由于加工后五倍子废渣中的成分比较复杂,因此对SP和SS的影响机制以及其具体组分的变化有待进一步研究。
3.4 五倍子废渣对小麦产量及其构成因素的影响
本研究表明,随着废渣添加量的增加,小麦各产量构成因素呈降低的趋势。产量构成因素均与籽粒产量之间存在不同程度的相关关系。生物产量、穗粒数、穗长以及收获期的株高与籽粒产量的相关性非常高,表明通过添加废渣,以上4种因素对小麦产量的影响非常大。可能原因是:(1)高剂量五倍子废渣影响小麦叶片叶绿素的合成及影响作物碳源的积累速度,使其营养生长受到抑制从而影响了生殖生长。(2)随着五倍子废渣添加量的增加,作物体内ROS逐渐增加,其产生的ROS对作物本身影响比较大,在生殖生长早期,花药中可能ROS积累异常,影响花药的发育及花粉的成熟[39],导致雄性不育影响部分产量构成因素。Nguyen等[40]在水稻水分胁迫研究中发现,花药中细胞程序性死亡与氧胁迫的相互作用会导致花粉不育。(3)由于五倍子废渣的抑制作用,小麦叶片电解质外渗,部分游离氨基酸流失,影响养分的积累,导致在生殖生长过程中小麦生殖器官发育不良,产生花粉不育或败育的现象。本研究结果与麦博儒等[41]研究得出的“在模拟酸雨胁迫下小麦有效穗数、每穗粒数、千粒质量、结实率等均呈下降趋势”的结论相似。同时高剂量处理的小麦产量大幅度降低,A4-A6的HI分别较A0下降1.57%,6.18%和14.01%,且均低于0.4,这与Unkovich等[42]“水分胁迫显著影响小麦收获指数”的结论相似。
3.5 施肥对五倍子废渣作用的缓解效应
本研究表明,施肥能明显削弱五倍子废渣分解对小麦生长产生的抑制作用,当废渣分解到44 d时,对小麦进行追肥处理,66~88 d时,各处理间的抗氧化酶活性、渗透调节物质等各项指标总体上无显著差异,88 d时,A4~A6地上部分生物量明显提高,收获时A3~A6的产量构成因素(株高、穗长、穗粒数和千粒质量)都有明显提高,表明施肥对废渣的抑制效应有一定的缓解作用。可能原因是,一方面施肥增加了土壤中植物所需的营养元素,在一定程度上促进了植物的生长;另一方面是土壤中氮素的增加,为土壤微生物提供了充足的氮源,加速了废渣的分解,释放了更多的养分,进一步促进了植物的生长。杨珊珊等[23]研究植物所受的胁迫作用时,也通过施肥探讨了其缓解作用,得到了与本研究相似的结果。
4 结 论
在种植小麦的土壤中添加五倍子废渣,抑制了小麦的生长,其抑制作用随添加量的增加而呈现先增强后减弱的趋势;高剂量添加处理对小麦收获产量有较强的抑制作用。根据本试验中的处理剂量测算,当废渣添加量达到5 240 kg/hm2以上时,其抑制作用达到显著水平,因此五倍子废渣还田量在5 240 kg/hm2以下时,不会影响小麦的生长,通过施肥还可缓解五倍子废渣产生的抑制效应。
[参考文献]
[1]武汉大学生物系.本草纲目简编 [M].武汉:湖北人民出版社,1978:355-356.
Botany, Wuhan University.Compendium of materia medica [M].Wuhan:Hubei People’s Press,1978:355-356.
[2]张雅丽.五倍子没食子酸的制备抑菌作用及机理研究 [D].西安:陕西师范大学,2013.
Zhang Y L.The extraction preparation of gallic acid fromGallachinensistannin and its antimicrobial activities and related mechanisms [D].Xi’an:Shaanxi Normal University,2013.
[3]赵拉.五倍子制备没食子酸的研究进展 [J].化学与生物工程,2008(5):5-7.
Zhao L.Research advance on preparation of gallic acid from Chinese nutgall [J].Chemistry & Bioengineering,2008(5):5-7.
[4]盛达成.利用五倍子单宁酸生产废弃物制备鞣花酸研究 [D].长沙:湖南农业大学,2013.
Sheng D C.Study on manufacturing technology of ellagic acid from the wastes of processing tannin acid in Chinese gall [D].Changsha:Hunan Agricultural University,2013.
[5]Keny S J,Kumbhar A G,Thinaharan C.Gallic acid as a corrosion inhibitor of carbon steel in chemical decontamination formulation [J].Cormsion Science,2008,50(2):411-419.
[6]Kim Y J.Antimelanogenic and antioxidant properties of gallic acid [J].Biological & Pharmaceutical Bulletin,2007,30(6): 1052-1055.
[7]Beltran F J,Garcia-Araya J F,Giraldez.Gallic acid water ozonation using activated carbon [J].Applied Catalysis B Environmental,2006,63(3/4):249-259.
[8]刘慧来.药渣与化肥配施对马铃薯产量与品质的影响 [J].黑龙江农业科学,2015(9):48-50.
Liu H L.Effect of herb residue and chemical fertilizer on yield and quality of potato [J].Heilongjiang Agricultural Science,2015(9):48-50.
[9]何佳芳,陈龙,苟久兰,等.不同药渣基质配比对小白菜种苗质量的影响研究 [J].种子,2015,34(2):94-96.
He J F,Chen L,Gou J L,et al.Effect of different drug residue ratio to Chinese cabbage seedling quality [J].Seed,2015,34(2):94-96.
[10]裴毅,杜宪,聂江力,等.没食子酸对铜绿微囊藻的化感作用 [J].植物研究,2014,34(6):840-844.
Pei Y,Du X,Nie J L,et al.Allelopathy of gallic acid onMicrocystisaeruginosa[J].Bulletin of Botanical Research,2014,34(6):840-844.
[11]孙兆法,张淑霞,宋朝玉,等.中药渣和泥炭基质对一品红生长和盆花品质的影响 [J].天津农业科学,2008,14(6):40-45.
Sun Z F,Zhang S X,Song C Y,et al.Effect of Chinese medicine residue and peat substrate on growth and visual quality of Poinsettia [J].Tianjin Agricultural Sciences,2008,14(6):40-45.
[12]郑照强,夏洪应,彭金辉,等.响应曲面法优化废弃五倍子药渣制取活性炭的研究 [J].环境污染与防治,2013,35(3):5-9.
Zheng Z Q,Xia H Y,Peng J H,et al.Process optimization for the preparation of activate carbon from abandoned dregs of gallumt using response surface methodology [J].Environmental Pollution & Control,2013,35(3):5-9.
[13]刘登才,郑有良,兰秀锦,等.四川农业大学小麦研究所新品种:蜀麦969(2013年四川审定)[EB/OL].(2014-10-08)[2017-02-10].http://xms.sicau.edu.cn/?p=25&a=view&r=334.
Liu D C,Zheng Y L,Lan X J,et al.New varieties from Triticeae Research Institute, Sichuan Agricultural University: Shumai 969(Sichuan validation,2013) [EB/OL].(2014-10-08) [2017-02-10].http://xms.sicau.edu.cn/?p=25&a=view&r=334.
[14]熊庆娥.植物生理学实验教程 [M].成都:四川科学技术出版社,2003:72-85.
Xiong Q E.Plant physiology experiment [M].Chengdu: Sichuan Science & Technology Press,2003:72-85.
[15]Giannopolitis C N,Roes S K.Superoxide dismutases: I. Occurrence in higher plants [J].Plant Physiology,1977,59:309-314.
[16]许为钢,胡琳.小麦收获指数的改良 [J].国外农学:麦类作物,1994(6):51-53.
Xu W G,Hu L.Improvement of wheat harvest index [J].Foreign Agronomy: Triticeae Crops,1994(6):51-53.
[17]Williamson G B,Rich A D.Bioassays for allelopathy: measuring treatment responses with independent controls [J].Journal of Chemical Ecology,1998,14(1):181-182.
[18]黄化刚,李廷轩,杨肖娥,等.植物对铅胁迫的耐性及其解毒机制研究进展 [J].应用生态学报,2009,20(3):696-704.
Huang H G,Li T X,Yang X E,et al.Research advances in plant lead to lerance and detoxification mechanise [J].Chinese Journal of Applied Ecology,2009,20(3):696-704.
[19]王琳,段璋玲.Hg2+胁迫对小麦幼苗生长及其过氧化物酶、淀粉酶活性的影响 [J].安徽农业科学,2011,39(5):2604-2607.
Wang L,Duan Z L.Effects of mercury stress on wheat growth POD and amylase activity [J].Journal of Anhui Agricultural Sciences,2011,39(5):2604-2607.
[20]刘丽君,尹田夫,薛津,等.渗透胁迫对大豆幼苗过氧化物酶活性及脯氨酸含量的影响 [J].大豆科学,1987,6(3):221-224.
Liu L J,Yin T F,Xue J,et al.Studies on drought resistance of soybean seedling [J].Soybean Science,1987,6(3):221-224.
[21]潘洁,姜东,戴廷波,等.不同生态环境与播种期下小麦籽粒品质变异规律的研究 [J].植物生态学报,2005,29(3):467-473.
Pan J,Jiang D,Dai T B,et al.Variation in wheat grain quality grown under different climatic conditions with different sowing dates [J].Acta Phytoecologica Sinica,2005,29(3):467-473.
[22]黄建贝,胡庭兴,吴张磊,等.核桃凋落叶分解对小麦生长及生理特性的影响 [J].生态学报,2014,34(23):6855-6863.
Huang J B,Hu T X,Wu Z L,et al.Effects of decomposing leaf litter ofJuglansregiaon growth and physiological characteristics ofTriticumaestivum[J].Acta Ecologica Sinica,2014,34(23):6855-6863.
[23]杨珊珊,王茜,胡红玲,等.银木凋落叶分解对玉米生长的影响及施肥的缓解作用 [J].应用与环境生物学报,2015,21(4):770-776.
Yang S S,Wang Q,Hu H L,et al.Allelopathy ofCinnamomumseptentrionaleleaf litter on maize growth and relieving effect of fertilization [J].Chinese Journal of Applied and Environmental Biology,2015,21(4):770-776.
[24]蒋晓梅.五倍子加工剩余物对几种蔬菜生长的影响 [D].成都:四川农业大学,2016.
Jiang X M.Effect of Chinese gallnut processing residue on the growth of three vegetables [D].Chengdu:Sichuan Agricultural University,2016.
[25]王利利,董民,张璐,等.不同碳氮比有机肥对有机农业土壤微生物生物量的影响 [J].中国生态农业学报,2013,21(9):1073-1077.
Wang L L,Dong M,Zhang L,et al.Effects of organic manures with different carbon-to-nitrogen ratios on soil microbial biomass of organic agriculture [J].Chinese Journal of Eco-Agriculture,2013,21(9):1073-1077.
[26]Ezatollah E,Fariborz S,Farid S,et al.The effect of salt stress on antioxidant enzymes’ activity and lipid peroxidation on the wheat seedling [J].Notulae Botanicae Horti Agrobotanici Cluj-Napoca,2007,35(1):48-56.
[27]赵天宏,孙加伟,付宇.逆境胁迫下植物活性氧代谢及外源调控机理的研究进展 [J].作物杂志,2008(3):10-13.
Zhao T H,Sun J W,Fu Y.Advances of research on metabolism of plant reactive oxygen species and exogenous regulation under abiotic stresses [J].Crops,2008(3):10-13.
[28]王海波,黄雪梅,张昭其.植物逆境胁迫中活性氧和钙信号的关系 [J].北方园艺,2010(22):189-194.
Wang H B,Huang X M,Zhang Z Q.Relationship between reactive oxygen species and calcium signalling in plant stress [J].Northern Horticulture,2010(22):189-194.
[29]Breusegem F V,Vranova E,Dat J F,et al.The role of active oxygen species in plant signal transduction [J].Plant Sci,2001,161:405-414.
[30]克热木·伊力,袁琳,齐曼·尤努斯,等.盐胁迫对阿月浑子SOD、CAT、POD活性的影响 [J].新疆农业科学,2004,41(3):129-134.
Karim·Ali,Yuan L,Qiman·Yunus,et al.Effects of salt tolerance on activity of SOD,CAT,POD of pistachio [J].Xinjiang Agricultural Sciences,2004,41(3):129-134.
[31]王娟,李德全.逆境条件下植物体内渗透调节物质的积累与活性氧代谢 [J].植物学通报,2001,18(4):459-465.
Wang J,Li D Q.The accumulation of plant osmoticum and activated oxygen metabolism under stress [J].Chinese Bulletin of Botany,2001,18(4):459-465.
[32]夏民旋,王维,袁瑞,等.超氧化物歧化酶与植物抗逆性 [J].分子植物育种,2015,13(11):2633-2646.
Xia M X,Wang W,Yuan R,et al.Superoxide dismutase and its research in plant stress-tolerance [J].Molecular Plant Breeding,2015,13(11):2633-2646.
[33]张敏,蔡瑞国,韩金玲,等.盐胁迫对小麦萌发期幼苗抗氧化酶系统的影响 [J].河北科技师范学院学报,2010,24(3):15-19.
Zhang M,Cai R G,Han J L,et al.Effects of salt stress on the activities of antioxidant enzymes in wheat seeding [J].Journal of Hebei Normal University of Science & Technology,2010,24(3):15-19.
[34]高永生,陈集双.盐胁迫下镧对小麦幼苗叶片抗氧化系统活性的影响 [J].中国稀土学报,2005,23(4):490-495.
Gao Y S,Chen J S.Effects of La3+on antioxidant system in wheat seedling leaves under salt stress [J].Journal of the Chinese Rare Earth Society,2005,23(4):490-495.
[35]马旭俊,朱大海.植物超氧化物歧化酶(SOD)的研究进展 [J].遗传,2003,25(2):225-231.
Ma X J,Zhu D H.Functional roles of the plant superoxide dismutase [J].Hereditas,2003,25(2):225-231.
[36]李广敏,唐连顺,商振清,等.渗透胁迫对玉米幼苗保护酶系统的影响及其与抗旱性的关系 [J].河北农业大学学报,1994,17(2):1-5.
Li G M,Tang L S,Shang Z Q,et al.Effect of osmotic stress on protective enzyme systems in maize seedlings and their relationship to drought resistance [J].Journal of Agricultural University of Hebei,1994,17(2):1-5.
[37]王兰兰,刘新,刘贺楠,等.小麦幼苗对盐胁迫和水分胁迫的生理反应对比 [J].沈阳师范大学学报(自然科学版),2013,31(1):120-123.
Wang L L,Liu X,Liu H N,et al.Physiological responses of west seedling to water and salt stress [J].Journal of Shen-yang Normal University (Natural Science Edition),2013,31(1):120-123.
[38]靖姣姣,张颖,白志英,等.盐胁迫对小麦代换系渗透调节物质的影响及染色体效应 [J].植物遗传资源学报,2015,16(4):743-750.
Jing J J,Zhang Y,Bai Z Y,et al.The effects of salt stress on osmoregulation substance and chromosome of wheat substitution lines [J].Journal of Plant Genetic Resources,2015,16(4):743-750.
[39]彭巧慧,刘孟雨,姜净卫,等.生殖生长早期水分胁迫降低作物穗粒数的研究进展 [J].河北农业科学,2013,17(1):12-15,27.
Peng Q H,Liu M Y,Jiang J W,et al.Research progress of water stress leading to reduced grain bumber of crops during early reproductive stage [J].Journal of Hebei Agricultural Sciences,2013,17(1):12-15,27.
[40]Nguyen G N,Hailstones D L,Wilkes M,et al.Drought-induced oxidative conditions in rice anthers leading to a programmed cell death and pollen abortion [J].Journal of Agronomy and Crop Science,2009,195:157-164.
[41]麦博儒,郑有飞,梁骏,等.模拟酸雨对小麦叶片同化物、生长和产量的影响 [J].应用生态学报,2008,19(10):2227-2233.
Mai B R,Zheng Y F,Liang J,et al.Effects of simulated acid rain on leaf photosynthate, growth, and yield of wheat [J].Chinese Journal of Applied Ecology,2008,19(10):2227-2233.
[42]Unkovich M,Baldock J,Forbes M.Variability in harvest index of grain crops and potential significance for carbon accounting: examples from Australian agriculture [J].Advances in Agronomy,2010,105:173-219.
