低氧微环境对非小细胞肺癌细胞系A549细胞增殖和侵袭的影响
2018-04-04,,,,,
, ,, ,,
[1.陆军军医大学(第三军医大学)第一附属医院老年科,重庆 400038;2.陆军军医大学(第三军医大学)第一附属医院全科医学科,重庆 400038]
肺癌是最常见的恶性肿瘤,也是我国高发的恶性肿瘤。在2017年出版的世界卫生组织公布的《全球癌症报告》中,全球范围内的肺癌发病率和病死率迅速增长,位居恶性肿瘤的首位[1]。在所有类型肺癌中非小细胞型肺癌(non-small cell lung cancer,NSCLC)约占80%,其5年生存率仅为13%[2-3]。尽管在过去几十年间随着我国医药卫生事业的发展,新辅助化疗和个体化综合治疗的推广应用,肺癌的早期预防和治疗已经取得明显疗效,但是非小细胞肺癌的预后仍然很差,仍然为最常见的和病死率最高的癌症[4-5]。NSCLC主要致死原因之一是残余肺癌肿瘤细胞的异常增殖和远处转移,因此深入研究肺癌的异常增殖和远处侵袭转移的分子作用机制对于改善肺癌的临床治疗和患者预后具有重要的临床意义。在实体瘤迅速生长的过程中,短时间内肿瘤细胞的快速增殖和肿瘤细胞周围血管生长滞后二者间形成一对矛盾体,由于无法为肿瘤细胞提供足够的营养和氧气,导致肿瘤局部组织中形成缺血缺氧微环境[6-8]。低氧微环境能够进一步对肿瘤细胞的增殖、血管生成以及侵袭转移等产生影响[9-11]。研究表明,Wnt/β-catenin信号通路与肿瘤细胞的增殖、分化以及侵袭转移密切相关[12-13]。但有关Wnt/β-catenin信号通路对缺氧条件下的非小细胞肺癌A549细胞增殖活性和侵袭转移的作用,目前尚未见报道。本研究旨在通过研究体外低氧微环境对非小细胞肺癌A549细胞增殖和侵袭的影响,并探讨Wnt/β-catenin信号通路在这一过程中的可能作用机制。
1 材料与方法
1.1 材料
人非小细胞肺癌细胞系A549购自中国科学院上海生命科学研究院生物化学与细胞生物学研究所;TRIzol RNA提取试剂盒购自美国Invitrogen公司;HIF-1α及β-catenin引物均委托北京天一辉远生物科技公司设计合成;多克隆兔抗人HIF-1α及β-catenin购自美国Santa Cruz公司,单克隆兔抗人β-actin购自美国Cell Signaling Technology公司,辣根酶标记羊抗兔IgG购自武汉博士德生物科技公司;中分子量Protein marker 购自Thermo Fisher公司;ECL 超敏化学发光试剂盒购自武汉碧云天生物科技公司;胎牛血清购自美国GIBCO公司;含酚红RPMI-1640培养基购自美国HYCLONE公司;细胞裂解液(RIPA)及BCA蛋白定量试剂盒均购自武汉碧云天生物科技公司;6孔、96孔培养板、25 cm2塑料培养瓶以及24孔Transwell小室(孔径8 μm)均购自美国Corning Costar公司;侵袭实验所用基质胶(Matrigel)购自美国BD公司;Co170R-230-0200三气细胞培养箱(用于模拟缺氧细胞培养环境)购自德国Eppendorf公司;倒置相差显微镜购自日本Olympus公司;垂直电泳槽、转移电泳槽均购自北京六一电子仪器公司。
1.2 方法
1.2.1细胞培养非小细胞肺癌细胞A549在细胞培养箱(37 ℃、5%CO2、饱和湿度条件)下用含10%小牛血清、100 u/mL青霉素、100 μg/mL庆大霉素、含酚红RPMI-1640培养基中培养。用0.25%胰酶消化传代。细胞培养至融合70%~80%后,用无血清培养基饥饿过夜,再置入低氧培养箱中培养24 h,随后对A549细胞进行相应指标的检测。常氧培养条件为37 ℃、5%CO2、20%O2;低氧培养条件为1%O2。
1.2.2CCK-8测定不同培养条件下的生长曲线取对数生长期的第3代肺癌A549细胞系常规消化后,按1×104/mL的细胞密度接种于96孔板中,每孔200 μL,并设空白孔(只加入200 μL培养基),共接种2块板。分别置于低氧培养箱及常氧培养箱中培养,分别于3、6、12、24 h共4个时间点取5个细胞孔及1个空白孔行CCK-8检测,每孔加入20 μL CCK-8试剂,孵育2 h后取出在酶标仪450 nm处测定其吸光值(A),以时间(h)为横坐标,A为纵坐标绘制生长曲线。
1.2.3RNA提取和Real-time PCR检测RT-PCR检测Wnt/β-catenin的表达,将已经过处理的各组细胞用冰上预冷PBS洗涤3次,每孔中加入1 mL Trizol RNA提取液,按照产品说明书操作步骤提取细胞总RNA,紫外分光光度法测定A260/A280比值,测算RNA 浓度,重复3次。按照逆转录试剂盒将提取的RNA逆转录为cDNA后进行PCR 循环扩增。PCR所用引物由上海擎科生物科技公司上海分公司合成,引物序列如下:β-actin上游引物5’-ATGGGGAAGGTGAAGGTCG-3’,下游引物5’-GGGTCATTGATGGCAACAATATC-3’;HIF-1α上游引物5’-CGUACUGCACUCCAAAUATT’,下游引物5’-UAUUUGGAGUGCAGUAGCGTT-3’;β-catenin上游引物5’-GCTGCTGTTTTGTTCCGAATGT-3’,下游引物5’-GCCATTGGCTCTGTTCTGAAGA-3’。实时荧光定量PCR:反应体系20 μL,mixture 10 μL,探针引物1 μL,cDNA 2 μL。反应条件均为:95 ℃,3 min 预变性;94 ℃,30 s;48 ℃,30 s,72 ℃,1 min,扩增35个循环;72 ℃延伸10 min。PCR 产物进行1.5%琼脂糖凝胶电泳,凝胶图像扫描系统成像并进行灰度扫描,重复3 次。
1.2.4Western blot检测非小细胞肺癌细胞系A549经过常氧及缺氧培养之后,将6孔板中的贴壁细胞用冰上预冷的PBS洗涤3次后加入RIPA细胞蛋白裂解液,并使用细胞刮将细胞裂解液收集转移到1.5 mL EP管中,冰上静置裂解30 min,随后于12 000 rpm离心10 min,弃去细胞沉淀,取上清液-20 ℃储存。采用BCA 法测定蛋白浓度,加入5×loading buffer 将蛋白混匀后,置于95 ℃金属水浴锅中进行蛋白变性。配制12%SDS-PAGE,每孔加入30 μg蛋白样品上样,80 V电泳积聚蛋白,100 V电压使蛋白分离并使用湿转法电转至PVDF膜。转膜结束采用5%脱脂牛奶封闭1 h。用多克隆兔抗人的HIF-1α(1∶800)β-catenin、(1∶1 000)、单克隆小鼠抗人的β-actin(1∶3 000)分别与膜接触4 ℃孵育过夜后,用TBST在脱色摇床下洗膜3次,每次5 min,在加辣根酶标记羊抗小鼠或抗兔IgG,室温下孵育1 h后,用TBST洗膜3次,每次5 min,加入ECL化学发光显示剂在室温下反应1 min后,进行曝光。
1.2.5Transwell细胞侵袭实验检测细胞侵袭能力实验前所有试剂及器材均于冰上预冷,将Transwell小室置于6孔板内,将Transwell小室内膜均匀涂抹0.2 μg/μL的Matrigel 胶(matrigel与无血清培养基之比为1∶250 μL,37 ℃孵育20 min,使胶凝固,并放入培养箱中4 h凝固备用。次日将处于对数生长期肺癌A549细胞系在无血清DMEM培养液饥饿培养24 h后,使用0.25%EDTA胰蛋白酶消化,接着无血清DMEM培养基悬浮成单细胞悬液,将细胞密度调整至至4×105/mL,台盼蓝染色检测细胞活力95%以上。每个侵袭小室上室中加入无血清细胞悬液200 μL,在侵袭小室的下室加入含10%小牛血清的细胞培养液DMEM,每孔600 μL,分别置于常氧和低氧培养箱中培养24 h。培养结束后取出小室,用PBS分3次洗涤去除培养基。使用湿棉签轻柔擦去小室上层未穿过细胞后置于4%多聚甲醛中固定20 min。随后将小室置于室温下晾干并进一步使用结晶紫染色20 min。染色完后在倒置显微镜下观察穿过膜的细胞数,计数中间及四周5个高倍(400倍)镜下视野细胞数,计算平均数。
1.3 统计学方法

2 结果
2.1 低氧对肺癌A549细胞增殖能力的影响
非小细胞肺癌细胞系A549经过常氧和低氧培养24 h后采用CCK8法进行细胞增殖能力检测。结果显示:与常氧组相比,在低氧环境下培养的非小细胞肺癌细胞系A549的增殖能力显著增强,而且随着低氧培养时间的延长增殖效应越显著,差异具有统计学意义(P<0.05),见图1。

图1 低氧促进非小细胞肺癌细胞系的增殖
2.2 低氧对肺癌A549细胞侵袭能力的影响
非小细胞肺癌细胞系A549经过常氧和低氧培养24 h后采用Transwell侵袭实验对细胞侵袭能力进行检测。结果显示,与常氧对照组相比,在低氧环境下培养24 h后的非小细胞肺癌细胞系A549的侵袭能力显著增强,差异具有统计学意义(P<0.05),见图2。
2.3 低氧对Wnt/β-catenin信号通路的影响
在常氧和低氧环境下培养非小细胞肺癌细胞系A549 24 h后,分别采用RT-PCR和Western blot检测β-catenin的mRNA和蛋白表达。结果显示,与常氧组相比,低氧组A549的β-catenin mRNA和蛋白表达水平显著升高,差异均有统计学意义(P<0.05),表明低氧能够激活Wnt/β-catenin信号通路(图3)。
2.4 靶向沉默β-catenin对Wnt/β-catenin信号通路的影响
转染β-catenin特异性siRNA及β-catenin-NC于非小细胞肺癌细胞系A549 48 h后,分别采用RT-PCR和Western blot检测β-catenin mRNA和蛋白表达。结果显示与β-catenin NC组和空白对照组相比,β-catenin siRNA组的β-catenin明显下调,差异具有统计学意义(P<0.05),见图4。
2.5 β-catenin siRNA转染对低氧介导的非小细胞肺癌细胞增殖能力
对分别进行常氧培养和低氧培养的非小细胞肺癌细胞系A549进行细胞增殖能力检测。CCK8结果显示,与常氧组相比,在低氧环境下培养的A549细胞系随着培养时间的延长其增殖能力显著增强,差异具有统计学意义(P<0.05);而转染了β-catenin siRNA的细胞其增殖能力显著降低,差异具有统计学意义(P<0.05),见图5。
2.6 β-catenin siRNA转染对低氧介导的非小细胞肺癌细胞侵袭能力的影响
在Transwell小室迁移实验中,我们发现低氧处理24 h后,与常氧对照组相比低氧转染NC处理组中A549细胞的侵袭能力显著增强,差异具有统计学意义(P<0.05);而转染β-catenin siRNA组的细胞侵袭能力被显著抑制,差异具有统计学意义(P<0.05),见图6。
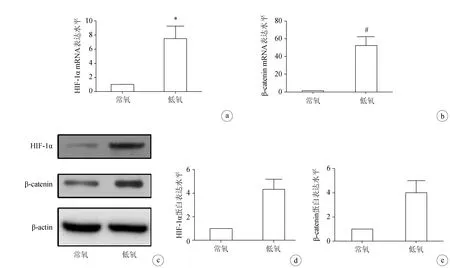
a:RT-PCR检测HIF-1α水平;b:RT-PCR检测β-catenin mRNA水平;c:Westtern blot检测HIF-1α和β-catenin蛋白表达水平;d、e:Westtern blot结果定量分析*:P<0.01;#:P<0.001
图3低氧激活非小细胞肺癌细胞系A549的Wnt/β-catenin信号通路

a:β-catenin siRNA转染荧光图(× 200);b:RT-PCR检测β-catenin mRNA表达;c:Western blot检测β-catenin 蛋白表达;d:Western blot结果定量分析*:P<0.001;#:P<0.01
图4β-cateninsiRNA在非小细胞肺癌细胞系A549中有效沉默β-catenin

图5 β-catenin siRNA抑制低氧诱导的细胞增殖
3 讨论
肺癌是目前危害人类健康最严重的恶性肿瘤之一,其发病率和病死率位居全球第一,探讨其发生机制以及寻找有效的治疗手段一直是受到广泛关注的研究热点之一[14-15]。在正常机体中,氧浓度的变化对组织细胞的代谢、增殖、分化具有重要的调节作用[16]。为了适应低氧代谢应激条件,肿瘤细胞能够通过激活一系列信号调控机制,如促进周边新生血管生成、增强细胞新陈代谢、迁移等,促进了细胞的异常增殖、转移、耐药等[17-18]。低氧环境下这一系列生物学事件的发生不仅改变了肿瘤细胞的生物学特性,引起肿瘤细胞的基因组不稳定性,导致肿瘤发生恶性转化甚至远处侵袭转移。作为最常见的实体瘤,肺癌病灶生长过程中随着肿瘤细胞的快速异常增殖,使得局部血供无法进一步支持肿瘤细胞的快速生长,不可避免地经历局部缺血缺氧微环境。而在这一过程中适应环境生存下来的肿瘤细胞的增殖又会继续加剧微环境的缺氧状况。在这样的缺血缺氧条件下,肿瘤细胞往往会通过自分泌或旁分泌作用改变自身周围生存和发展的环境,促进肿瘤继续生长和发展。肿瘤细胞本身的一些特征也会随之改变,例如具有更强的局部浸润和远处侵袭能力,使其更有利于脱离目前的恶劣环境,转移到新的部位重新定植。
Wnt/β-catenin 信号通路广泛参与生物体内细胞信息调控,其在细胞增殖、组织修复、胚胎发育及肿瘤(包括肺癌)的发生发展过程中发挥重要作用[19]。β-catenin是细胞内的一种多功能蛋白,其作为Wnt信号通路中的核心调控蛋白主要参与细胞之间粘附的调控。正常情况下,正常成熟的组织细胞中,胞浆内β-catenin与细胞膜上的E-cadherin相互结合起到促进细胞间粘附的作用。此时,胞浆内游离状态的β-catenin浓度维持在较低水平,无法进入胞核激活其下游靶基因。当细胞受到各种因素的刺激,使得Wnt信号通路激活时,Wnt配体与细胞膜表面的Frizzled/LRP受体结合从而磷酸化激活下游分子Dsh,使β-catenin从APC/Ax-in/GSK-3β组成的复合物中解离出来,使其在细胞内不断蓄积,并随后进入细胞核与TCF/LEF复合体结合,最终激活其下游靶基因的转录,从而发挥对细胞增殖、调亡、运动、分化等一系列生物学事件的调控[20-21]。研究表明,β-catenin与多种不同的肿瘤的发生发展密切相关,主要包括肝癌、胃癌以及肺癌等[22-24],这些研究均显示β-catenin在这些肿瘤组织中呈现高表达。在肝癌细胞中,Wnt/β-catenin通过增强低氧诱导的上皮间质转化促进细胞侵袭转移,同时β-catenin表达水平与肝癌术后存活时间呈负相关[25-26]。在结肠癌细胞中,低氧通过激活Nur77-β-catenin环路促进细胞的生长和侵袭[27]。然而,目前尚未有文献报道Wnt/β-catenin信号通路在低氧微环境下调控非小细胞肺癌细胞增殖和侵袭能力中的调控机制。
在本实验中,我们首先验证了体外低氧微环境对非小细胞肺癌细胞系A549的增殖与侵袭能力的影响。结果表明,与常氧对照组相比,A549细胞经过低氧环境下培养24 h后,其增殖速率显著高于常氧对照组,同时细胞侵袭能力的显著增强。接下来我们进一步研究低氧环境下这一现象的分子机制。研究发现,与常氧对照组相比,低氧环境下培养24 h后A549中β-catenin信号通路显著激活,提示其可能参与低氧环境下肺癌的异常增殖和侵袭过程。为了明确Wnt/β-catenin信号通路在其中的作用机制。我们采用特异性β-catenin siRNA来阻断其表达,并进一步检测其对细胞增殖和侵袭的影响。研究发现,在靶向沉默Wnt/β-catenin信号通路之后,低氧介导的细胞增殖和侵袭现象被阻断,表明Wnt/β-catenin信号通路在这一过程中起着关键作用。基于以上研究,我们推测低氧通过激活Wnt/β-catenin信号通路来调控下游靶基因转录激活,从而促进非小细胞肺癌细胞的异常增殖和远处侵袭转移,最终促进肺癌的发生发展。

a:Tanswell侵袭实验检测细胞侵袭能力(结晶紫染色 ×200);b:统计分析 *:P<0.001
综上所述,本文初步证实在肺癌A549细胞系中低氧微环境能够通过激活Wnt/β-catenin信号通路来促进肿瘤细胞的异常增殖和远处侵袭转移,这将有助于进一步认识肺癌的发病机制。因此,深入研究Wnt/β-catenin信号通路在肺癌发生、发展中的作用及其调节机制,通过靶向阻断Wnt/β-catenin信号通路相关分子,从而抑制肿瘤细胞的异常增殖和侵袭转移,这将为肺癌的特异性治疗提供新的思路和依据,具有广泛的临床意义和良好的应用前景。
[参考文献]
[1] Hong QY,Wu GM,Qian GS,et al.Prevention and management of lung cancer in China[J].Cancer,2015,121 (Suppl 17):3080-3088.doi:10.1002/cncr.29584.
[2] Pastorkova Z,Skarda J,Andel J.The role of microRNA in metastatic processes of non-small cell lung carcinoma[J].Biochim Biophys Acta,2016,160(3):343-357.doi:10.5507/bp.2016.021.
[3] Kumarakulasinghe NB,van Zanwijk N,Soo RA.Molecular targeted therapy in the treatment of advanced stage non-small cell lung cancer(NSCLC)[J].Respirology(Carlton,Vic),2015,20(3):370-378.doi:10.1111/resp.12490.
[4] Rafei H,El-Bahesh E,Finianos A,et al.Immune-based therapies for non-small cell lung cancer[J].Anticancer Res,2017,37(2):377-387.doi:10.21873/anticanres.11330.
[5] Durm G,Hanna N.Second-Line chemotherapy and beyond for non-small cell lung cancer[J].Hematol Oncol Clin North Am,2017,31(1):71-81.doi:10.1016/j.hoc.2016.08.002.
[6] Parks K,Cormerais Y,Marchiq I,et al.Hypoxia optimises tumour growth by controlling nutrient import and acidic metabolite export[J].Mol Aspects Med,2016:3-14.doi:10.1016/j.mam.2015.12.001.
[7] Harris BH,Barberis A,West CM,et al.Gene expression signatures as biomarkers of tumour hypoxia[J].Clin Oncol(R Coll Radiol),2015,27(10):547-560.doi:10.1016/j.clon.2015.07.004.
[8] Gilkes DM,Semenza GL,Wirtz D.Hypoxia and the extracellular matrix:drivers of tumour metastasis[J].Nat Rev Cancer,2014,14(6):430-439.doi:10.1038/nrc3726.
[9] Hubbi ME,Semenza GL.Regulation of cell proliferation by hypoxia-inducible factors[J].Am J Physiol Cell Physiol,2015,309(12):775-782.doi:10.1152/ajpcell.00279.2015.
[10] Madaneckip P,Kapoor N,Bebok Z,et al.Regulation of angiogenesis by hypoxia:the role of microRNA[J].Cell Mol Biol Lett,2013,18(1):47-57.doi:10.2478/s11658-012-0037-0.
[11] Yuen A,Díaz B.The impact of hypoxia in pancreatic cancer invasion and metastasis[J].Hypoxia(Auckl),2014,2:91-106.doi:10.2147/HP.S52636.
[12] Vilchez V,Turcios L,Marti F,et al.Targeting Wnt/beta-catenin pathway in hepatocellular carcinoma treatment[J].World J Gastroenterol,2016,22(2):823-832.doi:10.3748/wjg.v22.i2.823.
[13] Yang Y,Yang JJ,Tao H,et al.New perspectives on beta-catenin control of cell fate and proliferation in colon cancer[J].Food Chem Toxicol,2014,74:14-19.doi:10.1016/j.fct.2014.08.013.
[14] Yand J,Chen J,He J,et al.Wnt signaling as potential therapeutic target in lung cancer[J].Expert Opin Ther Targets,2016,20(8):999-1015.2016,20(8):999-1015.doi:10.1517/14728222.2016.1154945.
[15] Sihoe AD,Van Schil P.Non-small cell lung cancer:when to offer sublobar resection[J].Lung cancer(Amsterdam,Netherlands),2014,86(2):115-120.doi:10.1016/j.lungcan.2014.09.004.
[16] Eskandani M,Vandghanooni S,Barar J,et al.Cell physiology regulation by hypoxia inducible factor-1:targeting oxygen-related nanomachineries of hypoxic cells[J].Int J Biol Macromol,2017,99:46-62.doi:10.1016/j.ijbiomac.2016.10.113.
[17] Marchiq I,Pouysségur J.Hypoxia,cancer metabolism and the therapeutic benefit of targeting lactate/H(+) symporters[J].J Mol Med(Berl),2016,94(2):155-171.doi:10.1007/s00109-015-1307-x.
[18] Manoochehri Khoshinani H,Afshar S,Najafi R.Hypoxia:a double-edged sword in cancer therapy[J].Cancer Invest,2016,34(10):536-545.doi:10.1080/07357907.2016.1245317.
[19] Yao H,Ashihara E,Maekawa T.Targeting the Wnt/beta-catenin signaling pathway in human cancers[J].Expert Opin Ther Targets,2011,15(7):873-887.doi:10.1517/14728222.2011.577418.
[20] Zhang K,Zhang J,Han L,et al.Wnt/beta-catenin signaling in glioma[J].J Neuroimmune Pharmacol,2012,7(4):740-749.doi:10.1007/s11481-012-9359-y.
[21] Guo Y,XiaoL,Sun L,et al.Wnt/beta-catenin signaling:a promising new target for fibrosis diseases[J].Physiol Res,2012,61(4):337-346.
[22] Monga SP.Role of Wnt/beta-catenin signaling in liver metabolism and cancer[J].Int J Biochem Cell Biol,2011,43(7):1021-1029.doi:10.1016/j.biocel.2009.09.001.
[23] Cai J,Feng D,Hu L,et al.FAT4 functions as a tumour suppressor in gastric cancer by modulating Wnt/beta-catenin signalling[J].Br J Cancer,2015,113(12):1720-1729.doi:10.1038/bjc.2015.367.
[24] Chen X,Song X,Yue W,et al.Fibulin-5 inhibits Wnt/beta-catenin signaling in lung cancer[J].Oncotarget,2015,6(17):15022-15034.doi:10.18632/oncotarget.3609.
[25] Zhang Q,Bai X,Chen W,et al.Wnt/beta-catenin signaling enhances hypoxia-induced epithelial-mesenchymal transition in hepatocellular carcinoma via crosstalk with hif-1alpha signaling[J].Carcinogenesis,2013,34(5):962-973.doi:10.1093/carcin/bgt027.
[26] Liu L,Zhu X D,Wang WQ,et al.Activation of beta-catenin by hypoxia in hepatocellular carcinoma contributes to enhanced metastatic potential and poor prognosis[J].Clin Cancer Res,2010,16(10):2740-2750.doi:10.1158/1078-0432.CCR-09-2610.
[27] To SK,Zeng WJ,Zeng JZ,et al.Hypoxia triggers a Nur77-beta-catenin feed-forward loop to promote the invasive growth of colon cancer cells[J].Br J Cancer,2014,110(4):935-945.doi:10.1038/bjc.2013.816.
