电针对吗啡成瘾大鼠中枢多巴胺受体的影响
2018-03-22梁艳曾亮高晓悦张晴来祯
梁艳,曾亮,高晓悦,张晴,来祯
电针对吗啡成瘾大鼠中枢多巴胺受体的影响
梁艳,曾亮,高晓悦,张晴,来祯
(上海中医药大学附属岳阳中西医结合医院,上海 200437)
观察吗啡成瘾大鼠中脑腹侧被盖区(VTA)、伏隔核(NAc)、前额皮层(PFC)内多巴胺(DA)受体变化及电针的调节作用。30只雄性Wistar大鼠随机分为对照组、模型组和电针组,每组10只。采用吗啡自身给药成瘾大鼠模型,电针组选取双侧T5、L2夹脊穴,电针连续治疗4 d。Western Blotting法观察各组大鼠脑VTA、NAc、PFC内D1、D2受体含量的变化。治疗后,相对于模型组,电针组大鼠的吗啡摄入量明显减少(<0.05)。模型组大鼠脑VTA内多巴胺D2受体减少(<0.05);NAc内多巴胺D1受体增加(<0.05),D2受体减少(<0.05);PFC内多巴胺D1受体减少(<0.05),D2受体增加(<0.05)。电针治疗能调节上述脑区发生变化的D1、D2受体含量,使之趋于正常。电针治疗可以在一定程度上抑制成瘾大鼠对吗啡的渴求;吗啡成瘾后多巴胺投射通路中的多巴胺受体发生了一定程度的适应性改变,电针治疗可以调节DA受体的异常表达,对吗啡成瘾大鼠的多巴胺受体起到保护作用。
电针;吗啡依赖;行为,成瘾;多巴胺受体
药物渴求的中枢神经基础是从中脑腹侧被盖区到前脑的多巴胺投射通路,是阿片类药物引起精神依赖的轴心部位,也是脑内奖赏通路中药物作用最敏感部位,同时也是毒品依赖难以戒除的主要原因。多巴胺(DA)及其受体系统在药物奖赏机制中起重要作用。D1受体可能更多地参与奖赏介导的动机调控,而D2受体可能更多地参与药物的强化和药物依赖的获得。研究证实,药物成瘾是通过抑制多巴胺重吸收或促进多巴胺的释放增加突触间隙多巴胺的浓度,高于生理水平的多巴胺作用于相应的DA受体,产生神经适应性和成瘾行为。另一方面,DA受体也是多种药物作用的靶点,药物通过对其进行拮抗或激动来发挥作用[1]。本研究观察了电针对成瘾大鼠相关脑区中枢多巴胺受体的影响,现报道如下。
1 材料与方法
1.1 实验动物
30只Wistar大鼠,雄性,体重约250 g,SPF级,购自中国科学院上海实验动物中心。环境温度(24±2)℃,明/暗周期12 h,自由饮水,术后单笼饲养,行为学实验期间每日限食20 g。
1.2 模型制作
参照文献[2-4]建立大鼠吗啡自身给药模型。吗啡注射液(东北制药集团沈阳第一制药有限公司,批号141004-1,10 mg/mL)。
实验预备期,大鼠开始实验前适应环境3~5 d。之后采用FR1程序(固定比率为1,大鼠1次触鼻即可获得1次奖赏)进行蔗糖训练,每日训练2 h,使动物建立触鼻与奖赏之间的关系。选取蔗糖触鼻行为训练稳定的动物行颈静脉插管手术。
手术及恢复期,腹腔注射戊巴比妥钠55 mg/kg麻醉,同时给予阿托品0.1 mL。大鼠麻醉后绑定,剪毛,以75%乙醇局部消毒,剪开右侧颈部皮肤,分离颈外静脉,结扎远心端,在近心端剪V型缺口,将静脉插管缓慢插入血管至心房上方并结扎,插管游离端,用背心固定,塞入静脉插管塞封口,缝合创口。术后每日2次经静脉插管给予0.2 mL青霉素肝素钠溶液(青霉素40万u/mL,肝素钠50 u/mL),用于抗感染和抗凝。
吗啡自身给药训练,采用FR1程序,吗啡每次注射量为0.75 mg/kg,每日2 h,实验结束后动物放回饲养笼。
半量实验,自身给药训练最后1 d使用半量程序(FR2,固定比率为2,大鼠2次触鼻即可获得1次奖赏),观察2 h。如动物的吗啡摄入量保持恒定,说明吗啡自身给药训练(造模)成功。
1.3 动物分组及处理
对照组,与其他组动物相同环境饲养,不予吗啡造模及治疗干预措施。模型组和电针组进行吗啡造模。电针组,造模成功后给予电针治疗,取双侧T5、L2夹脊,连接G6805-2电针仪,连续波,频率2 Hz,留针20 min,每日1次,连续4 d。穴位选择参照《实验针灸学》《实验动物针灸手册》的动物穴位图谱,以比较解剖和骨度分寸法为依据模拟定位。T5夹脊穴位于背部,平第5胸椎棘突下,脊柱旁开0.5寸,左右各一穴。L2夹脊穴位于背部,平第2腰椎棘突下,脊柱旁开0.5寸,左右各一穴。
1.4 观察指标
大鼠行为学指标,蔗糖溶液给予次数,吗啡摄入量。
采用Western Blot方法检测大鼠中脑腹侧被盖区(VTA)、伏隔核(NAc)、前额皮层(PFC)区的D1、D2受体蛋白表达。
1.5 统计学方法
采用SPSS16.0统计软件进行数据分析。对符合正态分布计量资料采用均数±标准差进行统计描述,组间比较采用或独立样本检验、或采用方法进行两两比较;组内比较采用配对检验;对不符合正态分布的计量资料采用非参数检验。以<0.05为差异有统计学意义。
2 结果
2.1 电针对吗啡成瘾大鼠吗啡摄入量的影响
2.1.1 两组蔗糖训练结果比较
蔗糖训练期内每组动物连续5 d获得的0.2 mL 5%蔗糖溶液次数比较均无差异(>0.05),说明各组动物的蔗糖训练行为在造模前具有可比性。详见表1。
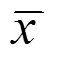
表1 两组蔗糖训练结果比较 (±s,次)
2.1.2 两组治疗前吗啡摄入量的比较
重复测量方差分析显示,两组间大鼠吗啡摄入量比较无明显差异(=1.669,>0.05),组别与时间之间无交互作用(=1.569,>0.05),两组内不同时间大鼠吗啡摄入量有差异(=2.159,<0.05),吗啡成瘾造模过程中存在时间差异,吗啡摄取量随时间递增客观上呈现增多的趋势,不影响模型稳定。两组大鼠吗啡自身给药9 d后的平均吗啡摄入量比较无明显差异(=1.075,>0.05)。说明各组大鼠的吗啡摄入量在治疗前具有可比性。详见表2。
表2 两组治疗前吗啡摄入量的比较 (±s,mg)
2.1.3 两组半量实验吗啡摄入量比较
模型组和电针组大鼠自身给药1~9 d FR1程序下平均吗啡摄入量比较,差异无统计学意义(>0.05)。两组大鼠自身给药的第10天,与F1程序比较,两组在FR2程序下吗啡摄入量基本不变(>0.05)。说明动物在变换实验程序后对吗啡的渴求没有降低,提示使用FR2程序后动物对吗啡的渴求行为是一种渴望得到药物的表现。详见表3。
表3 两组半量实验吗啡摄入量比较 (±s,mg)
2.1.4 两组治疗后吗啡摄入量比较
两组治疗后FR1程序下吗啡摄入量比较,差异有统计学意义(<0.05)。详见表4。
表4 两组治疗后吗啡摄入量比较 (±s,mg)
注:与模型组比较1)<0.05
2.2 电针对吗啡成瘾大鼠中脑腹侧被盖区(VTA)D1、D2受体含量的影响
在吗啡成瘾大鼠VTA的D1受体含量方面,各组比较无明显统计学差异(>0.05)。说明吗啡造模及电针治疗对大鼠VTA的D1受体含量影响不明显。在吗啡成瘾大鼠VTA的D2受体含量方面,模型组及电针组D2受体含量较对照组明显减少,差异具有统计学意义 (<0.05);与模型组比较,电针组D2受体含量明显增加,差异有统计学意义(<0.05)。说明吗啡造模可以使大鼠VTA的D2受体明显减少;电针治疗能使其含量增加。详见图1、表5。

图1 电针干预后VTA多巴胺D1、D2受体表达比较
表5 3组VTA D1、D2受体含量比较 (±s)
注:与对照组比较1)<0.05;与模型组比较2)<0.05
2.3 电针对吗啡成瘾大鼠伏隔核(NAc)D1、D2受体含量的影响
吗啡成瘾大鼠NAc的D1受体含量比较发现,模型组、电针组D1受体含量较对照组明显增加,差异具有统计学意义(<0.05);与模型组比较,电针组D1受体含量明显减少,组间比较差异有统计学意义(<0.05)。说明吗啡造模可以使大鼠NAc的D1受体含量增加,电针治疗可以减少NAc的D1受体的含量。吗啡成瘾大鼠NAc的多巴胺D2受体含量比较发现,模型组、电针组D2受体含量较对照组明显减少,差异具有统计学意义(<0.05);与模型组比较,电针治疗后D2受体含量增加,组间比较差异无统计学意义(>0.05)。说明吗啡自身给药模型可以使大鼠NAc的D2受体含量减少,电针治疗后NAc的D2受体含量有增加的趋势。详见图2、表6。

图2 电针干预后NAc多巴胺D1、D2受体表达
表6 3组NAc D1、D2受体含量比较 (±s)
注:与对照组比较1)<0.05;与模型组比较2)<0.05
2.4 电针对吗啡成瘾大鼠前额皮层(PFC)D1、D2受体含量的影响
在吗啡成瘾大鼠PFC的D1受体含量方面,模型组、电针组与对照组比较有明显差异(<0.05);模型组与电针组比较无明显统计学差异(>0.05)。说明吗啡造模可以使大鼠PFC的D1受体含量明显减少;电针治疗对吗啡成瘾大鼠PFC的D1受体含量影响不明显。在吗啡成瘾大鼠PFC的多巴胺D2受体含量方面,模型组、电针组与对照组比较有明显差异(<0.05);与模型组比较,电针组D2受体含量明显减少,组间差异有统计学意义(<0.05);说明吗啡造模可以使大鼠PFC的D2受体含量增加,电针治疗能使其含量减少。详见图3、表7。
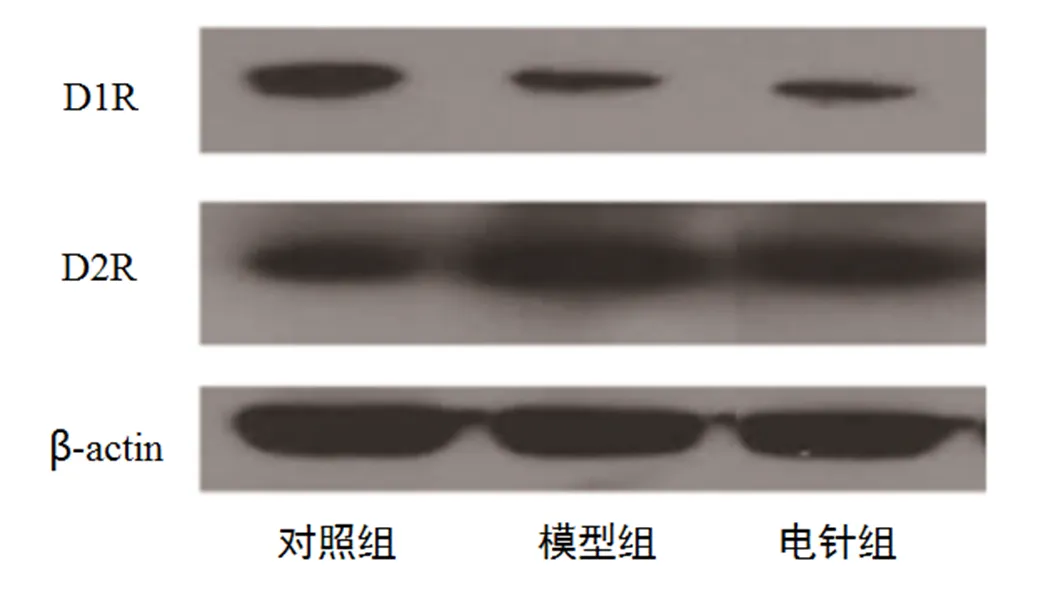
图3 电针干预后PFC多巴胺D1、D2受体表达
表7 3组PFC D1、D2受体含量比较 (±s)
注:与对照组比较1)<0.05;与模型组比较2)<0.05
3 讨论
3.1 选穴依据
临床研究时对99例海洛因成瘾者的脊柱压痛点进行了统计分析,发现压痛点集中出现在第5~7胸椎棘突下的人数占总调查人数的72.7%,出现在第2腰椎棘突下的人数占总调查人数23.4%。并且压痛点的疼痛程度随躯体症状的缓解而逐渐减弱[5]。华佗夹脊穴位于脊柱两侧旁开0.5寸,邻近背俞穴,刺之可调节脏腑气血功能;T5夹脊穴正是位于督脉神道附近,具有泄热清心、安神定志的作用。L2夹脊穴邻近肾俞穴,肾俞亦属背俞穴,二穴相配对脏腑、经络、气血津液的失调都有很好的调整作用。刺之可泄热清心、安神定志、培补元气、温肾化浊。《素问·长刺节论》中认为:“治寒热。深专者,刺大藏,迫藏刺背,背俞也。”海洛因依赖者,往往病程长而顽固,病理多为本虚标实,多侵及五脏,取背俞穴往往能取得较好的疗效。现代研究[6]表明,肾俞等穴对垂体、肾上腺功能有促进作用,能有效抑制吗啡戒断症状,针刺肾俞等穴,对长期使用海洛因所致的肾虚诸症都有较好的调整作用。有研究者[7]利用韩氏穴位神经刺激器针刺双侧足三里和三阴交,发现针刺对海洛因成瘾引起的大鼠细胞凋亡有抑制作用,并下调大鼠PAG内caspase-3的表达。
T5、L2夹脊穴对脏腑、经络、气血津液的失调都有很好的调整作用。根据辨证取穴理论和近年来各种报道的用穴频数,并考虑到动物实验操作中的难易程度,我们选择双侧T5、L2夹脊穴为治疗位点;T5、L2夹脊正是督脉神道、至阳穴附近,两穴有泄热清心、安神定志、疏肝理气的作用,可调节戒毒者的阴阳平衡,达到改善阿片类药物戒断者焦虑和抑郁情绪的作用[8]。
3.2 电针对吗啡成瘾大鼠的影响
强迫性反复用药是药物成瘾的主要特征,无论实验动物或者临床患者在脱毒后仍然存在对药物的渴求。本次实验研究结果显示,经过蔗糖、吗啡诱导已形成稳定吗啡自身给药行为的大鼠,在停药戒断后,大鼠对吗啡的渴求并没有完全消失。这一实验结果与报道的研究结果是一致的,电针对线索诱导的大鼠海洛因觅药行为有明显抑制作用,这可能与扣带前皮质、伏隔核核区、伏隔核壳区CREB的表达增强及腹侧被盖区抑制有关[9]。也表明大鼠对药物强化效应所引起的极度欣快的记忆非常深刻。
吗啡戒断期间给予电针治疗后,大鼠再次返回实验笼的行为数据显示,电针组动物的吗啡摄入量显著低于模型组(<0.05),提示吗啡戒断时给予电针治疗可以显著抑制成瘾大鼠对吗啡的渴求行为。这一研究结果与其他相关研究机构通过电针结合迷走神经刺激可减少条件线索诱导的大鼠海洛因觅药行为,增加其活动度的研究结果一致[10];也与通过条件性位置偏爱模型所观察到的结果具有一致性,电针能抑制或消除大鼠吗啡CPP表达[11-12],说明电针治疗有抑制动物渴求行为的作用。但吗啡戒断时电针治疗并没有使动物的觅药行为消失,而是对动物的觅药行为起到了一定程度的抑制,而这种抑制效应能维持多久,还有待进一步实验研究来观察和验证。
3.3 电针对吗啡成瘾大鼠相关脑区多巴胺受体的影响及机理探讨
多巴胺(DA)能神经元主要分布在纹状体、VTA及黑质等脑区,形成脑内两个DA能环路,其中一个为中脑边缘DA系统(MLDS),是参与奖赏效应的重要作用系统;MLDS神经元胞体位于VTA,属于A10群胞体,其上行路径经内侧前脑束投射至NAc、嗅结节、额叶皮质的颗粒区、前扣带回及边缘叶等的皮质,有些纤维可远至嗅球、杏仁核和梨状区皮层质,甚至投射至更为广泛的大脑皮质;到边缘系统的投射是情绪和情感的表达中枢,到前额叶皮层的投射参与认知、意识活动;这两者均与药物成瘾和精神分裂症有关[13]。中脑-边缘多巴胺系统、NAc的形态和功能改变是产生阿片类药物成瘾的重要神经基础。DA受体各亚型选择性地在中脑边缘多巴胺系统高度表达,同样提示DA在阿片类药物成瘾中发挥重要的作用。多巴胺也被一些学者认为是渴求效应产生的最终共同神经递质,甚至在应激诱发成瘾行为和促发复吸中糖皮质激素也通过影响DA系统发挥奖赏效应的强化作用。在急性给药的情况下,绝大多数成瘾药物通过激活MLDS中NAc和PFC的DA神经通路,实现药物的奖赏和强化作用;有证据表明NAc、PFC是产生吗啡强化效果的最终部位[14]。损伤NAc可以显著降低可卡因、苯丙胺、吗啡和尼古丁等的自身给药。
近半个世纪的研究结果已经形成共识,脑内奖赏或强化系统的主要结构——中脑-边缘多巴胺神经系统,是阿片类药物引起精神依赖的轴心部位,而且是阿片强化效应的最后通路,中脑-边缘系统和NAc的形态和功能改变是产生阿片类药物成瘾的重要神经基础[15-17]。DA受体各亚型选择性地在中脑边缘多巴胺系统高度表达,同样提示DA在阿片类药物成瘾中发挥重要的作用[18]。DA通过D1型受体参与吗啡引起的运动增强效应,但从作用机理上看,只是对抗吗啡的运动增强效应,没有其它的特异始动作用[19];同时D1型DA受体拮抗剂,可减少吗啡成瘾大鼠对环境刺激的反应,即降低了该大鼠感受欣快的反应,增加了大鼠的觅药动机,又抑制了该大鼠的行为活动,但D2型DA受体则不同,虽降低了该大鼠的觅药动机,却不影响该大鼠的行为活动,可见不同DA受体亚型在吗啡依赖形成过程中发挥不同的作用[20]。本研究小组其他研究结果显示电针治疗虽然降低了动物对吗啡的渴求和摄入量,但动物对吗啡的觅药行为依然存在[19]。这种变化和结果很可能与D1、D2受体的作用存在一定的关系。本次研究结果显示吗啡造模可以使大鼠脑VTA、NAc内D1受体含量减少,D2受体含量增加;使PFC内D1受体含量减少,D2受体含量增加;而电针治疗则可使各脑区异常的D2受体趋于正常,对NAc内D1受体含量的变化有调节作用,说明电针治疗可以调节与吗啡成瘾相关脑区D1、D2受体含量的变化,使之趋向于正常,并且这种作用更明显地表现在对各脑区D2受体的调节上,可以猜测电针对吗啡成瘾大鼠的干预作用有可能通过对上述脑区D2受体的调节而呈现的。但由于本次研究的样本量较小,这种猜测还有待后续研究结果来进一步证实。
本次实验结果显示电针效应存在一定的累积性,同时也进一步证明了本研究小组前期通过吗啡和海洛因自身给药模型观察到电针对大鼠渴求的抑制效应与刺激量有关。研究说明电针具有调节吗啡成瘾大鼠DA受体的作用,戒断期内连续电针的刺激并没有使DA受体恢复至正常水平,这种调节效应在今后的一段时间内能维持多久,还有待进一步的临床和实验研究来提供更为有力的临床和实验室证据。
[1] 周媛,邢淑华.左旋四氢巴马汀对苯丙胺条件性位置偏爱效应的影响[J].徐州医学院学报,2002,19(2):105- 107.
[2] WEEKS JR. Experimental morphine addiction: method for automatic intravenous injections in unrestrained rats[J]., 1962,138(3537):143-144.
[3] Hall BJ, Pearson LS, Terry AV Jr,. The use- dependent, nicotinic antagonist BTMPS reduces the adverse consequences of morphine self-administration in rats in an abstinence model of drug seeking[J]., 2011,61(4):798-806.
[4] Feltenstein MW, Do PH, See RE. Repeated aripiprazole administration attenuates cocaine seeking in a rat model of relapse[J].(), 2009,207(3): 401-411.
[5] 胡军,陆英,辛玉虎,等.海洛因依赖者脊柱阳性反应点观察[J].中国针灸,2000,20(10):611-613.
[6] 刘慧英,潘书贵.电针对海洛因成瘾大鼠学习记忆及海马CREB表达的影响[J].癫痫与神经电生理学杂志, 2011,20(5):35-38.
[7] 刘胜,周文华,刘惠芬,等.针灸“肾俞”穴对吗啡依赖大鼠戒断反应和成瘾脑区神经元活动的影响[J].中国针灸,2004,24(11):805-808.
[8] 梁艳,黄寅,程波,等.电针内关、神门穴对海洛因依赖者稽延性戒断症状疗效观察[J].上海针灸杂志,2011, 30(12):804-806.
[9] 王浩然,高祥荣,张开镐,等.药物成瘾及成瘾记忆的研究现状[J].生理科学进展,2003,34(3):202-206.
[10] 刘宜军,周文华,张富强,等.迷走神经刺激结合电针对条件线索诱导觅药行为的干预作用及其分子生物学机制[J].河南中医,2008,28(11):38-41.
[11] 史向党,罗非,崔彩莲,等.电针抑制大鼠吗啡条件性位置偏爱[J].北京大学学报(医学版),2003,35(3):248- 251.
[12] 李莎莎,康林,吴宾,等.电针对吗啡依赖大鼠条件性位置偏爱效应的作用研究[J].四川大学学报(医学版), 2004,35(6):824-827.
[13] 王学铭.精神与精神病的生物化学[M].北京:人民卫生出版社,2002:265.
[14] Zhu ZC, HU J, XU P. Effect of acupuncture on contents of tyrosine hydroxylase and glial fibrillary acidic protein in ventral tegmental area of heroin self-administrating rats [J]., 2006,4(1):7-12.
[15] 王惠玲,赵晏.阿片类药物依赖与中脑—边缘多巴胺系统回路[J].中国药物依赖性杂志,2003,12(1):2-5.
[16]Saika F, Kiguchi N, Wakida N,.Upregulation of CCL7 and CCL2 in reward system mediated through dopamine D1 receptor signaling underlies metham- phetamine-induced place preference in mice[J].,2017,665:33-37.
[17] Volkow ND, Wise RA, Baler R. The dopamine motive system: implications for drug and food addiction[J]., 2017,18(12):741-752.
[18] Beaulieu JM, Gainetdinov RR. The physiology, signaling, and pharmacology of dopamine receptors[J]., 2011,63(1):182-217.
[19] 邓宏勇,胡军,梁艳.电针对吗啡成瘾大鼠戒断-复吸行为影响的初步观察[J].上海针灸杂志,2004,23(8): 31-34.
[20] Rodriguez-Arias M, Broseta I, Aguilar MA,. Lack of specific effects of selective D(1) and D(2) dopamine antagonists vs. risperidone on morphine-induced hyper- activity[J]., 2000,66(1):189-197.
Effect of Electroacupuncture on Central Dopamine Receptor in Morphine-dependent Rats
,,-,,.
,,200437,
To observe the change of dopamine receptor in ventral tegmental area (VTA), nucleus accumbens (NAc) and prefrontal cortex (PFC) in morphine-dependent rats, and the regulation effect of electroacupuncture (EA).Thirty male SD rats were randomized into a control group, a model group and an EA group, 10 rats each. Morphine-dependent rat models were induced by morphine self-administration. In the EA group, bilateral Jiaji points (EX-B2) of T5and L2were selected, and the EA intervention lasted 4 d. Western blotting method was adopted to observe the change of the contents of dopamine receptor D1 and D2 in VTA, NAc and PFC of the rats.After the intervention, compared to the model group, the morphine intake was reduced significantly in the EA group (<0.05); the level of D2 declined significantly in VTA of rats in the model group (<0.05); the level of D1 increased significantly and D2 declined significantly in NAc of rats in the model group (<0.05); the level of D1 declined significantly and D2 increased significantly in PFC of rats in the model group (<0.05); EA produced regulation effect on the altered contents of D1 and D2 in the cerebral areas mentioned above, approaching the normal level.EA can inhibit the hunger of addiction rats for morphine to some extent; the contents of dopamine receptors in dopamine projection pathway will take adaptive changes after morphine addiction, while EA can regulate the abnormal expressions of dopamine receptors, producing a protective effect on dopamine receptors of morphine- dependent rats.
Electroacupuncture; Morphine dependence; Behavior, Addiction; Dopamine receptor
1005-0957(2018)13-0324-06
R2-03
A
10.13460/j.issn.1005-0957.2018.13.0324
2017-09-16
国家自然科学基金青年基金项目(81403473);上海市卫生和计划生育委员会科研课题资助项目(20134292)
梁艳(1976―),女,副主任医师,博士,Email:dr.liangy@163.com
