施肥对闽楠幼树PSⅡ叶绿素荧光快速光响应曲线的影响
2018-03-19杨佳伟文仕知何功秀杨丽丽
杨佳伟,文仕知,何功秀,兰 竹,杨丽丽
(中南林业科技大学 林学院,湖南 长沙 410004)
闽楠Phoebe bournei (Hemsl.) Yang又称楠木,樟科楠属,是南方珍贵用材树种和优良庭院观赏树,属国家二级保护珍稀濒危树种[1],主要分布于福建、江西、湖南、湖北和贵州等地区的常绿阔叶林[2]。目前,闽楠已被列入国家林业局重点发展的南方珍贵用材树种之一,并在育苗和栽培方面进行一系列研究[3],目前的主要研究集中在种源选择[4]、种子休眠与萌发[5]、抗逆性筛选[6]、种群结构和空间布局[7]、生物量结构[8]、育苗造林[9-10]以及光合、叶绿素荧光生理特性[11]。
植物光合作用是指在叶绿体类囊体中叶绿素等光合色素分子吸收外界光能,同时将光能转化为化学能,形成ATP和NADPH的光反应过程[12-13]。光合作用中植物将吸收的光能转化为体内电能过程,部分光能作用于光化学反应,部分光能以热耗散形式消耗,还有部分光能则由于电子门开启,从光系统Ⅱ以荧光的形式散发,即叶绿素荧光是植物吸收的小部分光重新以光的形式发射出来[14]。与净光合速率、胞间CO2浓度等一些“表观性”的气体交换指标相比,叶绿素荧光参数在探测逆境对光合作用影响方面具有独特的作用,能够反映光合系统“内在性”特性[15]。有研究认为,Cd2+等重金属元素对植物光系统Ⅱ的活性有抑制作用[16],还能降低光合色素含量[17];李泽等人[18]的研究表明,根外追肥对油桐幼苗的光合促进作用显著;另有研究表明,闽楠幼树在不同光环境下生物量比例没有显著差异,从而得出结论闽楠是通过生物量积累来适应不同光环境,而非改变生物量比例[19];石元豹等[20]对宁夏枸杞关于CO2浓度的研究显示,CO2浓度倍增将提高枸杞光能转化率;张伟[5]、刘宝[21]等学者研究了不同种源间的叶绿素荧光参数,结果显示种源间光合荧光不同程度存在差异性,光合利用能力各不相同;不同光照强度等级对有柄石韦的相对导电率、相对叶绿素含量、光合荧光参数和植株形态有显著影响[22]。
本研究以4年生闽楠幼树为试验对象,设置一个空白对照样地,4个施肥处理样地,对闽楠叶绿素荧光参数和快速光响应曲线进行田间活体测定,比较施肥对林木荧光参数的影响和各样地的快速光响应曲线差异,以寻求高效的施肥量,为闽楠人工培育提供基础数据。
1 材料与方法
1.1 试验材料与处理
本研究以4年生闽楠幼树作为施肥对象,施用有机复合肥,含有机质28.88%,N11.81%,有效 P2O53.06%,K2O 3.37%,Pb 60.00 mg/kg,Cr 40.00 mg/kg,As 1.64 mg/kg,Hg 0.016 mg/kg。
试验样地位于攸县黄丰桥林场,地理坐标为113°41′0″E,27°18′25″N,设置一个空白样地,样地编号1,四个施肥试验样地,分别按照150、300、450、600 g/株肥量进行施肥处理样地依次编号为2,3,4,5。各样地林木种植约90株,采用水平带状整地后实生苗种植。
样地在2014年秋季追肥,2014年12月和2015年6月进行数据采集。各样地根据等同坡位选取3株样地固定平均木进行叶绿素荧光参数测定。
1.2 荧光参数的测定
每株样木选取树冠外缘向阳的成熟叶片,采用德国WANL公司的MINI-PAM便携式荧光仪进行叶绿素荧光动力学参数和光响应曲线的测定。测定植株最小荧光Fo、最大荧光Fm、PSⅡ的最大量子产量Fv/Fm是需要使用暗适应叶夹对叶片进行暗适应处理三十分钟,后用荧光仪测定光响应曲线参数。植株快速光响应曲线的测定,对光适应样品进行连续光量子能量梯度的光照射(0、60、150、250、400、600、800、1 200、1 600 μmol·m-2s-1),各梯度光强照射10 s后仪器发射饱和脉冲[6],从而测定光量子产量Yield、相对电子传递速率ETR、荧光淬灭系数等荧光参数。
1.3 数据处理
使用Excel2013进行数据处理、制图,所有参数值均为三个重复数据取平均值;SPSS进行方差分析,检验差异显著性;快速光响应曲线拟合使用Statistica 10.0软件进行拟合。
2 结果与分析
2.1 施肥对闽楠幼树Fo、Fm、Fv/Fm、Fv/Fo的影响
Fo的测定是植株经过充分暗适应之后,打开一个只激发本底荧光的微弱调制测量光而不引起任何光合作用时测量光得到的荧光值,一定程度上与叶片色素含量线性相关。
在暗适应条件下下叶片初始荧光Fo,是光系统Ⅱ所有反应中心均处于开放状态时得到的最小荧光值(图1)。随着施肥量的增大,冬季Fo值先减小,在450 g/株之后又增大,单株林木施肥量600 g/株的时候其基本初始荧光Fo值与样地1未施肥林木Fo值平齐。随着肥效的深入,到夏季闽楠幼树荧光Fo值相比于冬季数值明显变小,各样地对比则是随施肥量的增加先增大然后减小,节点是300 g/株(P>0.05)。
Fm值的变化趋势和Fo相似,在冬季随着施肥量的增加先减小后增大,在600 g/株的时候显著升高,达到最大值;到夏季生长旺盛期随施肥量的增加先增大后减小,300 g/株施肥量出现峰值(P>0.05)。其中未施肥样地和单株林木施肥量150 g/株的监测数据在两次监测中差异都不显著。
PSⅡ光化学量子效率(Fv/Fm),在冬季的测量数据,差异不显著,而夏季的测量数据显示单株林木施肥量600 g/株的样地5数据稍低于其他4种处理,两个时间段的测量值总体水平都在0.8±0.03(P>0.05)。Fv/Fm值是反应光系统Ⅱ反应中心利用所捕获激发能的情况,测定数据说明施肥量的差异对于光系统Ⅱ反应中心的开放比例没有显著影响,光合电子链的传递速率没有因为施肥量的差异而有所显著不同。
Fv/Fo值的测定显示,在冬季施肥样地的数值高于未作施肥处理的样地1,且四个施肥样地的差异不显著,而夏季数据表明样地5的600 g/株施肥量其Fv/Fo最低。Fv/Fo值是反映PSⅡ活性的一个指标,从测定的数据看来,冬季施肥样地的光系统Ⅱ活性略高于未作施肥处理的对照样地,而夏季的高温天气对于闽楠幼树来说,适量的施肥可以促进PSⅡ反应中心活性,当施肥量超过450 g/株时反而会一定程度上对PSⅡ活性形成抑制。
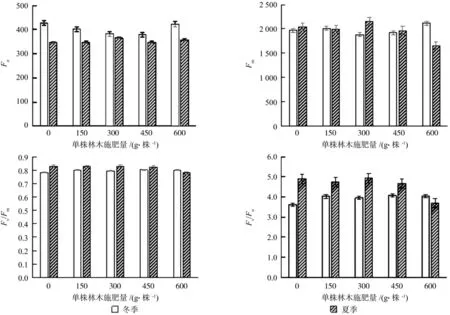
图1 施肥对闽楠幼树Fo、Fm、Fv/Fm、Fv/Fo的影响Fig. 1 Responses of Fo, Fm, Fv/Fm, Fv/Fo of Phoebe bournei to fertilization
2.2 施肥对闽楠幼树Yield、ETR的影响
冬季植株的PSⅡ光量子产量Yield随光强的增加平缓降低,直到1 600 μmol·m-2s-1时光量子产量Yield达到最小值,各处理样均是同一变化趋势(图2)。同时期测量的植株叶片PSⅡ相对光合电子传递速率 ETR随着光强的增加逐渐增大,在1 600 μmol·m-2s-1光照强度的时候达到最大值,各处理的变化趋势一致(图3)。冬季闽楠的Yield值变化趋势相近,但是总体来看样地5的施肥处理样值高于其他样地,且差异较为显著;而ETR随光强的变化表明样地5的600 g/株施肥量的相对电子传递速率高于其他处理。
到夏季PSⅡ光量子产量Yield随光强增加而降低,在光照初期,Yield值有一个显著降低,而后降低趋势较为平缓,直到最大光照强度时为最低值(图2)。相对光合电子传递速率 ETR的变化趋势同冬季相差不大,随着光照强度的增加而增大,直到光照强度800 μmol·m-2s-1之后其增长幅度逐渐趋于平缓,在1 400 μmol·m-2s-1时达到最大值(图3)。夏季闽楠的Yield值变化趋势相近,但是总体来看样地1未作施肥处理的样地值明显低于施肥样地,且差异较为显著,在弱光照条件下(<400 μmol·m-2s-1)样地3施肥量水平下 Yield值高于其他样地,在较强光照条件下样地2的施肥量处理Yield值较高;而ETR随光强的变化表明样地1未施肥处理的相对电子传递速率明显低于其他施肥处理。
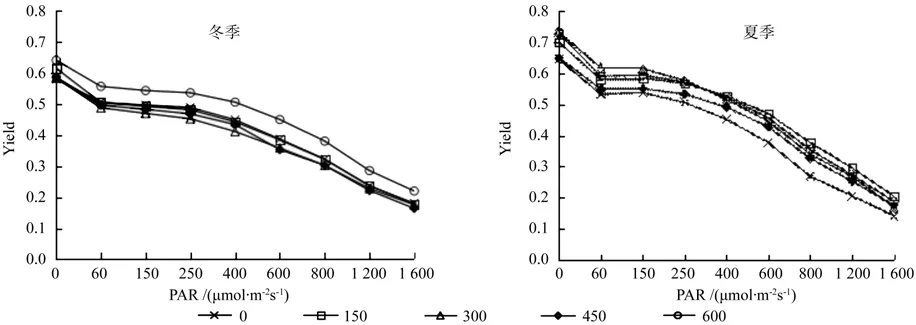
图2 施肥对闽楠Yield快速光响应的影响Fig. 2 Rapid light-response curve of Yield about Phoebe bournei to fertilization
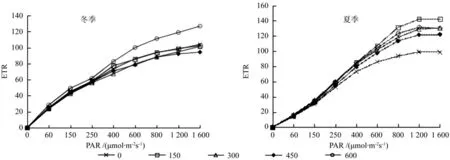
图3 施肥对闽楠ETR快速光响应的影响Fig. 3 Rapid light-response curve of ETR about Phoebe bournei to fertilization
2.3 施肥对闽楠幼树qP、qN、NPQ的影响
在光照下光合作用进行时,只有部分电子门处于关闭状态,实时荧光F比最大荧光Fm要低,这是荧光淬灭。现有的研究显示,植物吸收的光能被用于三个方面,光合作用、叶绿素荧光、热,即荧光产量的下降是由光合作用的增加或热耗散增加而引起[24]。由光合作用增加引起的荧光淬灭称为光化学淬灭,既是qP。而由于热耗散的增加引起的荧光淬灭称之为非光化学淬灭,即为qN、NPQ。
qP值是反映PSⅡ反应中心的开放程度,冬季的监测数据随着光照强度的增大,qP值以平缓线形的趋势变小,在光照强度为1 600 μmol·m-2s-1时达到最小值;在弱光照下未施肥样地数据qP值趋于各样地的均值水准,随光照强度增大,其值的减小程度强于其他施肥样地,整体数值低于其他样地,样地3和样地4施肥标准的qP值较为突出。夏季的测定数据随光强增加而降低,降低趋势较冬季更为明显,也是在最大光强时达到最小值,样地1的未施肥处理也和冬季趋势一致整体数值低于其他样地,样地2的150 g/株和样地4的450 g/株施肥标准qP值比较突出(图4)。
qN值是反映植物耗散过剩光能为热的能力,即光保护能力。如图5所示,冬季qN值的整体趋势是随光强增加而增大,在低光照强度下(<400 μmol·m-2s-1)时增大较为平缓,超过 400 μmol·m-2s-1光强时qN值增大速率显著提高,样地4和样地3的样本热耗散能力相对强于其他处理样本。夏季qN值的整体变化趋势同冬季一致,只是在低光照条件下施肥样地的值增长较样地1未施肥的样本相对缓慢,当光强增大到250 μmol·m-2s-1时施肥样地的qN值随光强增加迅速增大,但整体来看还是未施肥样地的qN值较为明显,而施肥量最大的样地5整体值偏小(P>0.05)。由于冬夏季的日光照强度差异,两个时期测量的qN的最大值不相同,表明闽楠幼树在不同季节,其叶片耗散热量的能力有所差异,冬季中度施肥的样本热耗散能力强,而夏季高温季节则是未施肥的空白样地热耗散能力高。
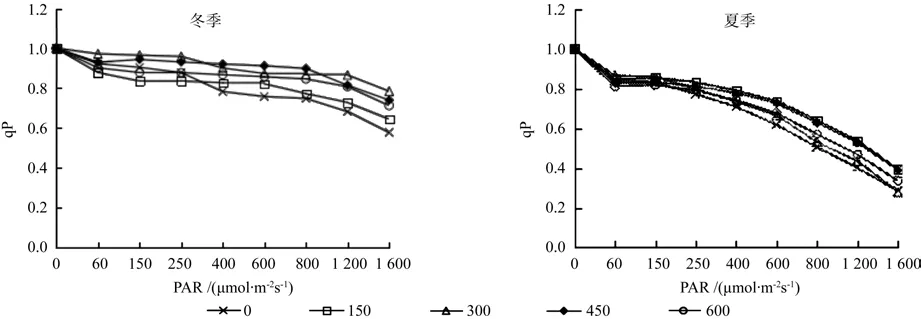
图4 施肥对闽楠qP快速光响应的影响Fig. 4 Rapid light-response curve of qP about Phoebe bournei to fertilization

图5 施肥对闽楠qN快速光响应的影响Fig. 5 Rapid light-response curve of qN about Phoebe bournei to fertilization
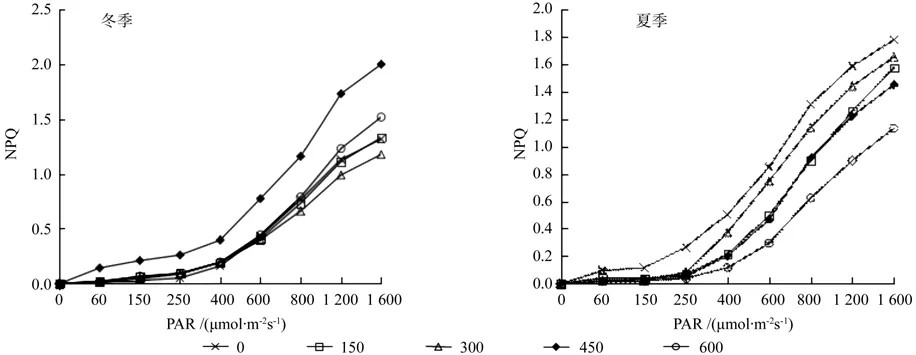
图6 施肥对闽楠NPQ快速光响应的影响Fig. 6 Rapid light-response curve of NPQ about Phoebe bournei to fertilization
NPQ值是非光化学淬灭的另一种表达方式,同样是作为反映植物耗散过剩光能的能力指标,是更注重于由天线系统中的热耗散引起的非光化学淬灭。两个季度的测量数据趋势和快速光响应的qN值趋势一致,冬季是样地4的样本数据高于其他样地,夏季依然是样地1未作施肥处理的样本数值较高,其次是样地3和样地5的施肥处理(图6)。
2.4 施肥对高温季节闽楠幼树快速光曲线的影响
光响应曲线是指植物光合速率随PAR变化而变化的曲线,光合放氧速率(Pn)、相对电子传递速率(ETR)、CO2固定速率等都可用于光响应曲线的绘制[25]。快速光响应曲线(快速光曲线)则是运用调制叶绿素荧光技术,通过各梯度较短光照时间测量得出的光响应曲线。
图7是拟合的高温季节不同施肥样地闽楠幼树的快速光响应曲线,拟合公式为1980年Platt提出的 P=Pm·(1-e-a·PAR/Pm)·e-b·PAR/Pm[26]。式中 Pm 是最大光合速率;a为初始斜率,反映植物的光能利用效率;b为光抑制参数;lk=Pm/a表示半饱和光强,反映样品对强光的忍受能力。
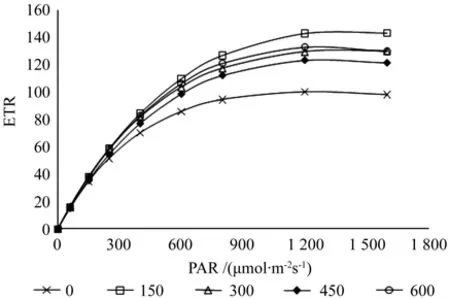
图7 拟合高温季节不同施肥量闽楠幼树快速光响应曲线Fig. 7 Fitting the rapid light curves about different fertilization of Phoebe bournei saplings in hot season
从图7中可以看出,各施肥样本的最大光合速率Pm都是出现在光照强度1 200~1 400 μmol·m-2s-1之间,同王振兴等[11]的研究中光饱和点基本一致,Pm值由大到小分别是150 g/株,600 g/株,300 g/株,450 g/株,空白样地;样本的初始斜率a值由大到小依次为300 g/株,600 g/株,150 g/株,空白样地,450 g/株,值分别为0.293>0.282>0.281>0.274>0.267,其值大小间接反映样本植株的光能利用效率。分析不同施肥量的闽楠拟合曲线可知,样地1的未施肥处理和样地2、3、4的施肥处理其光合速率趋势相同,而样地5的600 g/株施肥量样本光合速率在光合有效辐射1 300 μmol·m-2s-1附近开始有一个明显地降低趋势,说明过多的施肥在高温季节高光强时易对植株产生光合胁迫。
3 讨 论
施肥是常规的营林措施,但是不同作物其经营方式不同,施肥重点不一,施肥量亦有所差异,寻求闽楠幼树的合理施肥量是本试验重点。叶绿素荧光参数的变化与光合作用各反应阶段都有一定相关性,作物的生长过程中的各种胁迫往往能通过叶绿素荧光诱导动力学参数反应出来。初始荧光Fo与PSⅡ反应中心活性相关,本试验中两个季节的监测数据变化趋势相反,冬季闽楠幼树未施肥和大量施肥的样本Fo值较高说明PSⅡ反应中心相较于中度施肥样本活性较弱,夏季则是相反。Fm值是PSⅡ反应中心关闭时的荧光值,值越大说明反应中心关闭程度越高[27],冬季600 g/株的施肥样地Fm高于其他处理,夏季则是300 g/株施肥样地Fm较高,而600 g/株的施肥样地Fm最低,说明大量施肥会增大高温季节的闽楠幼树PSⅡ反应中心开放程度。Fv/Fm是植株暗适应之后开放的PSⅡ反应中心捕获激发能的效率,能反应植物受胁迫和受伤害的程度,本试验中,各施肥处理样地的Fv/Fm在同一观测时期内无明显差异,样地3的300 g/株施肥量在夏季略高于其他样地,而样地5的600 g/株施肥处理值明显偏低,与整体变化趋势不符,说明高温季节大量施肥会使植株光系统组织受到一定胁迫。
Yield值冬季测量数据显示闽楠幼树的随光强的增大平缓降低,样地5的600 g/株施肥处理值明显高于其他样地,而夏季测量结果是样地1未施肥样地的各光强对应光量子产量低于其他施肥样地;PSⅡ相对光合电子传递速率 ETR随光强的变化表明大量施肥在高温季节对闽楠幼树的相对电子传递速率促进作用不显著,ETR在1 300 μmol·m-2s-1光强时饱和,反应中心部分关闭或失活,过剩光能只能通过非光化学淬灭形式耗散;光化学荧光淬灭系数qP与非光学荧光淬灭系数qN、NPQ,说明夏季高温季节施肥量的大小会影响闽楠幼树叶片的热耗散能力,施肥量越大,叶片耗散过量光能的压力越大。
综上所述,中度施肥导致闽楠幼树Fv/Fm、Fv/Fo值升高,大量施肥在高温季节则降低。施肥对冬季生长缓慢期的闽楠PSⅡ反应中心开放程度有促进作用,高温季节闽楠PSⅡ反应中心开放程度显著提升,但随着光强的增加,大量施肥样本过剩能量以非光化学荧光淬灭形式耗散,会导致PSⅡ反应中心受损,易变现出光抑制现象,建议秋季施肥以促进冬季PSⅡ反应中心开放程度,以增进冬季的物质累积。
本文的研究只是针对不同施肥处理后闽楠在不同生长期的光合荧光特性,研究结果内容相对单一,研究结论尚不能指导生产。下一步研究方向会将闽楠施肥光合参数结合林木生长量、生态化学计量数据作相关性分析,寻找闽楠生长的限制性因子,以达到有侧重点地施肥,促进林木生长。
[1]何功秀,文仕知,邵明晓,等. 湖南永顺闽楠人工林生态系统碳贮量及其分布特征[J]. 水土保持学报,2014, 28(5):159-163.
[2]曾祥谓. 我国多功能森林经营中的珍贵树种问题研究[D]. 北京:中国林业科学研究院,2010.
[3]刘 军,姜景民,陈益泰,等. 闽楠种子轻基质容器育苗及优良家系选择[J]. 西北林学院学报,2011, 26(6):70-73.
[4]吴际友,黄明军,陈明皋,等. 闽楠种源苗期生长差异与早期选择研究[J]. 中南林业科技大学学报,2015, 35(11):1-4.
[5]张 伟. 闽楠种子萌发技术与幼苗荧光特性的研究[D]. 长沙:中南林业科技大学,2013.
[6]朱亚红,孙存举,朱铭强,等. 河北杨与沙棘的光合生理特性及叶绿素荧光参数比较[J]. 中南林业科技大学学报,2015,35(2): 26-31.
[7]葛永金,王军峰,方 伟,等. 闽楠地理分布格局及其气候特征研究[J]. 江西农业大学学报,2012, 34(4):749-753.
[8]邵明晓. 不同林龄闽楠人工林生物量与碳密度研究[D]. 长沙:中南林业科技大学,2014.
[9]葛永金. 闽楠栽培生理生态基础研究[D]. 北京:北京林业大学,2014.
[10]申 展. 闽楠无性繁殖技术研究[D]. 长沙:中南林业科技大学,2013.
[11]王振兴,朱锦懋,王 健,等. 闽楠幼树光合特性及生物量分配对光环境的响应[J]. 生态学报,2012, 32(12):3841-3848.
[12]Salgado-Luarte C, Gianoli E. Herbivory may modify functional responses to shade in seedlings of a light-demanding tree species.Functional Ecology, 2011, 25(3):492-499.
[13]曹 刚. 不同LED光质对黄瓜和结球甘蓝苗期生长、光合特性及内源激素的影响[D]. 兰州:甘肃农业大学,2013.
[14]许楠. 黑龙江省桑树氮肥运筹及光合调控机理的研究[D]. 哈尔滨:东北林业大学,2013.
[15]郝岩松. 模拟酸雨对墨西哥柏幼苗光合生理特性的影响[D].南京:南京林业大学,2013.
[16]Bernier M, Popovic R, Carpentier R. Mercury inhibition at the donor side of photosystemⅡis reversed by chloride[J]. FEBS Letters, 1993, 321(1):19-23.
[17]Baszyńki T, Wajda L, Król M, et al. Photosynthetic activities of cadmium-treated tomato plants[J]. Physiologia Plantarum, 1980,48(3): 365-370.
[18]李 泽,谭晓风,卢 锟,等. 根外追肥对油桐幼苗生长、光合作用 及叶绿素荧光参数的影响[J]. 中南林业科技大学学报,2016, 36(2):40-44.
[19]Valladares F, Niinemets. Shade tolerance a key plant feature of complex nature and consequences[J]. Annual Review of Ecology,Evolution, and Systematics, 2008, 39(1): 237-257.
[20]石元豹,曹 兵. CO2浓度倍增对宁夏枸杞叶绿素荧光参数的影响[J]. 经济林研究,2015, 33(3):108-111.
[21]刘 宝,陈存及,林达定,等. 21个闽楠种源叶片光合色素含量及叶绿素荧光参数分析[J]. 江西农业大学学报,2014, 36(1):115-121.
[22]杨柳青,杨 帆,杨 琳. 不同光照条件对有柄石韦的形态和生理特性的影响[J]. 经济林研究,2016, 34(2):109-113.
[23]SchreibeiU, GademannR, RalphPJ, et al. Assessment of photosynthetic performance of Prochloron in Lissoclinum patella in hospite by chlorophyll fluorescence measurements[J].Plant Cell and Physiology, 1997, 38(8): 945-951.
[24]周丽娜. 基于叶绿素荧光光谱分析的稻叶瘟病害识别与预警[D]. 长春: 吉林大学,2014.
[25]郭春燕,李晋川,岳建英,等. 两种高质牧草不同生育期光合生理日变化及光响应特征[J]. 生态学报,2013, 33(6):1751-1761.
[26]P.H.C. Eilers, J.C.H. Peeters. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton[J]. Ecological Modelling, 1988, 42(3): 199-215.
[27]高培培,章 艺,吴玉环,等. 外源水杨酸对铝胁迫下栝楼光合特性及耐铝性的影响[J]. 水土保持学报,2012, 26(6):268-273.
