广西4个油茶品种苗期对干旱胁迫的生理响应
2018-03-19洪文泓黄永芳龚海光韦雪芬蓝来娇
董 斌 ,洪文泓 ,黄永芳 ,薛 立 ,龚海光 ,韦雪芬 ,蓝来娇
(1.华南农业大学 林学与风景园林学院,广东 广州 510642;2.广东农工商职业技术学院,广东广州 510507;3. 中国科学院华南植物园,广东 广州 510650)
油茶Camellia oleifera,为山茶科山茶属植物,是我国栽培历史悠久,种植面积最广泛的木本油料作物。油茶树具有耐贫瘠,耐干旱的生长特性,适宜丘陵山地的生长环境,在中国南方低山丘陵地区广泛种植。但是,由于丘陵山地普遍灌溉条件受限,尤其是油茶主产区雨旱季节性明显的亚热带季风性气候,旱季供水严重不足导致油茶生长受限。而此时正值油茶花芽发育、果实膨大、油脂转化的关键时期,干旱缺水影响油茶树体生长,导致枝叶焦枯,光合作用受抑制,果实大量脱落,油脂合成降低,严重影响产量[1]。因此,研究油茶抗旱性对于提高茶油产量具有重要意义。
针对油茶抗旱性生理的研究已见不少报道。已有学者对长林3号,湘林5号,湘林52号,闽43号和凤阳1号等油茶优良无性系一、二年生苗开展抗旱生理的研究[2-6],周招娣等[7]研究了4种不同砧木对湘林11号一年生嫁接苗抗旱性的影响;霍佩佩等[8]和冯士令等[9]分别针对长林系列8个和9个种优良无性系二年生苗开展了抗旱性综合评价,通过多指标对抗旱性进行排序。
总体而言,油茶品种间抗旱性的差异较大,对部分油茶品种的抗旱性研究少见报道,且已有研究多以一年生或二年生幼苗为试验材料。此外,采取多指标综合评价油茶抗旱性的研究更少开展。本研究采用隶属函数分析法,从多个生理生化指标入手对产自广西且栽培面积较广的4个油茶品种幼苗抗旱性进行综合评价,以期筛选出抗旱能力较强的品种,为油茶的苗木繁育,抗旱育种以及向干旱地区推广提供参考依据。
1 材料与方法
1.1 供试苗木
试验材料为2015年4月产自广东南亚热带作物研究所的桂无1号(桂1),桂无4号(桂4),岑软2号(岑2),岑软3号(岑3)共4个引自广西的优良无性系一年生油茶嫁接苗,砧木为岑软2号、3号油茶子一代实生苗。将油茶苗统一移植入12 cm×8 cm的育苗袋中,基质为黄壤土,土肥,光照条件保持一致,随机摆放在华南农业大学经济林研究中心苗圃地。在圃地统一培育6个月后,挑选长势基本相同,无病虫害的健壮植株进行PEG干旱胁迫试验,幼苗生长基本情况见表1。
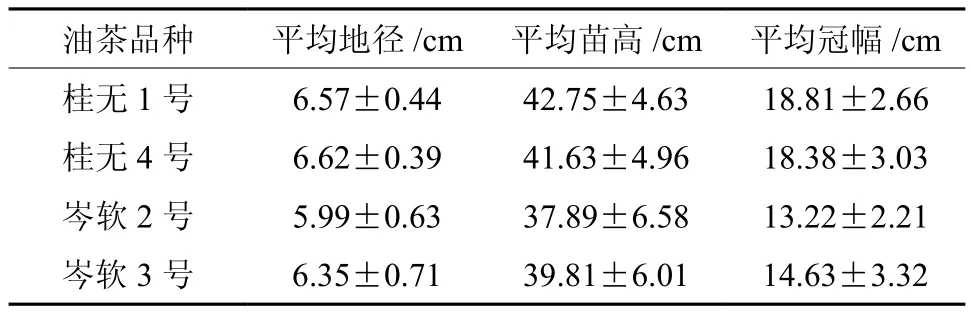
表 1 4种油茶品种幼苗的基本情况(平均值±标准差)Table 1 General characteristics of four seedlings (mean±SD)
1.2 实验方法
1.2.1 不同干旱胁迫程度处理
在新鲜的1/2Hoagland营养液里加入PEG6000配制成不同浓度的胁迫溶液,分别为轻度胁迫10%(100 g/L PEG)、中度胁迫20%(200 g/L PEG)、重度胁迫30%(300 g/L PEG)。将苗的根部浸入胁迫溶液中进行不同程度的干旱胁迫处理。
1.2.2 不同干旱胁迫时间的处理
各胁迫强度均设4个时间处理,分别在12,24,36,48 h时采集叶片,其中12、24、36 h为持续干旱胁迫,48 h为模拟复水处理,即在胁迫36 h后将苗放置在1/2Hoagland营养液中继续培养12 h。以在1/2Hoagland营养液中缓苗2 d后的幼苗为对照(0 h),测定各项生理指标。
1.2.3 幼苗处理
选取大小长势基本相同的油茶幼苗,剥落泥土并洗净根部。将幼苗随机分为4组,用海绵板固定于3个装有5 L去离子水的塑料盆中。水培苗先在1/2Hoagland营养液中恢复2d后移入用营养液配制的具不同渗透势的PEG6000溶液中,进行模拟干旱胁迫和后续模拟复水。实验均在BIC-250智能型人工气候室进行,温度为28±2 ℃,相对湿度为75%~85%,给予光照强度2 500 lx,16 h白天和8 h黑暗的光周期处理,早晚各通气1次。幼苗处理后,选择生长良好、大小均一的叶片为试材,在8:00和20:00采样,每处理设3次重复,随机采自顶芽第3~8片完全展开叶,用去离子水清净吸干,混合后进行生理指标测定。每个品种均按此方法进行操作。
1.3 生理指标测定
叶片相对含水量按照饱和含水量法测定;叶片相对电导率用电导法测定;过氧化物酶(POD)活性用愈创木酚法测定;叶绿素含量测定用分光光度法测定;丙二醛(MDA)含量用硫代巴比妥酸(TBA)比色法;可溶性蛋白含量采用考马斯亮蓝G-250 染色法;可溶性糖按照蒽酮比色法测定。以上指标和相对含水量均参照陈建勋等的方法测定[10],叶绿素含量用丙酮提取法测定[11]。每指标3次重复。
1.4 数据统计
采用Microsoft Excel 2016、SPSS 17.0 软件进行数据处理与制表,多重比较和隶属函数分析。
采用隶属函数法对4个不同油茶品种的抗旱性进行综合评价。根据不同指标的均值,用下列公式计算不同指标的隶属函数值[12]。
隶属函数计算公式:
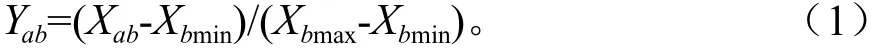
反隶属函数计算公式:
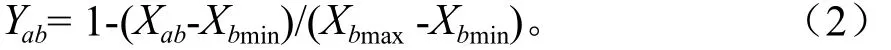
式中:Yab表示a品种b指标的隶属函数值;Xab表示a品种的b 指标的均值;Xbmax、Xbmin分别表示所有品种b指标平均值的最大值和最小值。若b指标与抗旱性呈正相关,用式(1);若b 指标与抗旱性呈负相关,用式(2)。最后把每个品种各指标的抗旱隶属值累加,并求平均数,平均数越大表示抗性越强。
2 结果与分析
2.1 干旱胁迫对相对含水量的影响
4个油茶品种在轻度、中度和重度干旱胁迫下,叶片相对含水量均呈下降趋势。其中,桂4和岑2干旱胁迫后各阶段相对含水量均显著低于对照(P<0.05),桂1和岑3除轻度胁迫12 h与对照差异不显著外,其余均显著低于对照(P<0.05)。总体而言,桂4各阶段叶片相对含水量下降幅度要明显大于其余3个品种,岑3相对含水量下降幅度较少。复水12 h后,除桂4轻度胁迫和岑3中度胁迫显著上升外,其余品种各处理均变化不显著(P<0.05)。
2.2 干旱胁迫对相对电导率的影响
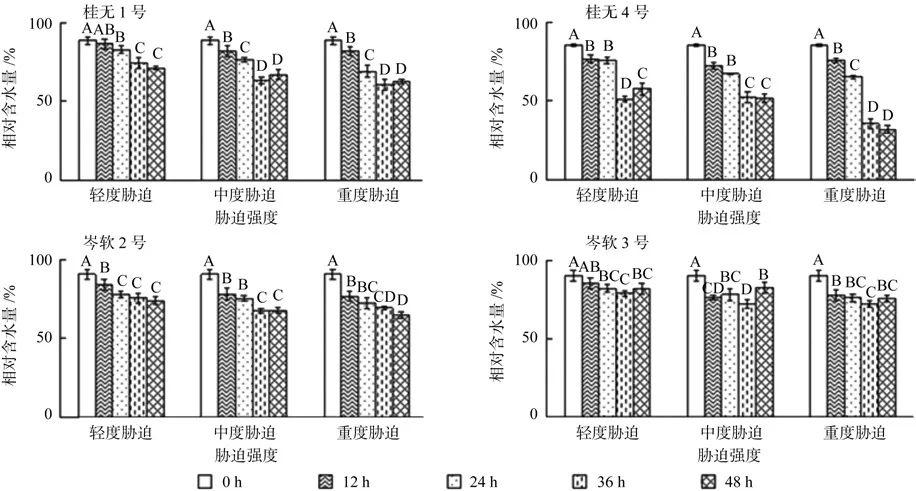
图1 不同干旱胁迫持续时间对各幼苗相对含水量的影响(不同字母表示差异性显著)Fig. 1 Effect of drought stress on the relative water content in leaves of seedlings
4个油茶品种幼苗叶片在各胁迫处理下相对电导率均显著大于对照(P<0.05)(图2)。其中桂4各胁迫强度在胁迫初期12 h相对电导率即快速升高,桂1在24 h,岑2和岑3在36 h才出现电导率快速升高现象;复水12 h后,桂4和岑2轻度胁迫,桂1中度胁迫,岑3中度和重度胁迫电导率显著下降(P<0.05),其余处理变化不显著或显著上升。总体而言,岑3在整个实验过程中,电导率变化幅度最小。
2.3 干旱胁迫对叶绿素含量的影响
从图3可见,桂1和桂4各胁迫强度36 h叶绿素含量均显著低于对照(P<0.05),岑2和岑3各胁迫强度36 h叶绿素含量则显著高于对照(P<0.05)。4个油茶品种胁迫处理前24 h叶绿素含量变化较为波动,其中桂1和岑2波动上升,桂4基本持平,岑3波动下降。复水12 h后,岑2中度、重度胁迫和岑3各胁迫强度叶绿素含量显著下降,其余品种和胁迫强度变化不显著(P<0.05)。
2.4 干旱胁迫对POD活性的影响
从图4可见,干旱胁迫导致各油茶品种POD活性均不同程度增加,其中桂1和桂4的总体增幅较小,岑2和岑3的总体增幅较大。在干旱胁迫下,桂1各胁迫强度POD活性均呈现先减少,后增加,再减少的波动变化趋势;桂4在轻度和中度胁迫12 h 的POD活性虽高于其余时间点,但增幅较少;岑2和岑3轻度和重度胁迫12 h的POD活性即大幅度显著增加(P<0.05)。复水12 h后,除岑2重度胁迫和岑3各胁迫强度POD活性显著降低外,其余品种及胁迫强度均增加。
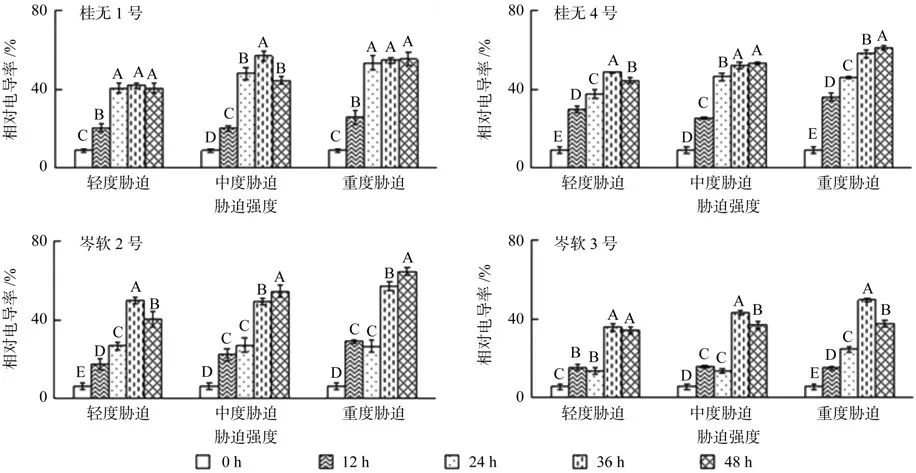
图2 不同干旱胁迫持续时间对各幼苗相对电导率的影响(不同字母表示差异性显著)Fig. 2 Effect of drought stress on the relative electrical conductivity in leaves of seedlings
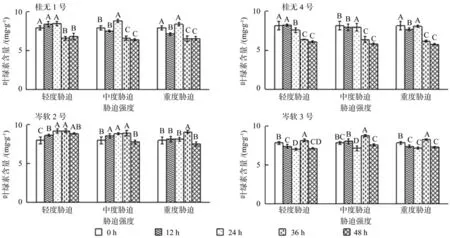
图3 不同干旱胁迫持续时间对各幼苗叶绿素含量的影响(不同字母表示差异性显著)Fig. 3 Effect of drought stress on the chlorophyll content in leaves of seedlings
2.5 干旱胁迫对丙二醛含量的影响
干旱胁迫处理后4个油茶品种幼苗叶片MDA含量均增幅显著(P<0.05),随着胁迫时间的延长总体呈现上升趋势(图5)。其中桂1,桂4和岑2在初始胁迫12 h的MDA含量已大幅度增加,尤其是桂1和桂4中度和重度胁迫已与峰值差异不显著(P<0.05);岑3的叶片MDA含量随胁迫时间的延长逐步增加。复水12 h后,桂1和岑3的轻度胁迫叶片MDA含量显著下降(P<0.05),其余品种和处理变化不显著。
2.6 干旱胁迫对可溶性蛋白含量的影响
如图6所示,干旱胁迫环境条件下,4个油茶品种叶片可溶性蛋白含量总体呈上升趋势,其中桂1和岑3整体增幅较显著,尤其是岑3反应较迅速,其可溶性蛋白含量在胁迫12 h即有反应,且中度和重度胁迫12 h即增幅显著(P<0.05),其余品种各胁迫强度大多在24 h才显著增加,12 h时变化幅度很小。复水12 h后,除桂1、桂2重度胁迫,岑3各胁迫强度可溶性蛋白显著下降外,其余品种和处理均差异不显著(P<0.05)。
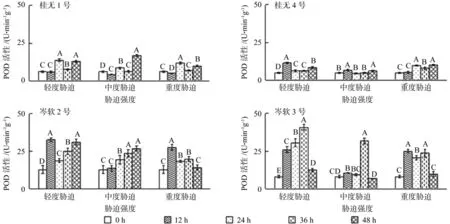
图4 不同干旱胁迫持续时间对各幼苗POD活性的影响(不同字母表示差异性显著)Fig.4 Effect of drought stress on the activity of POD in leaves of seedlings
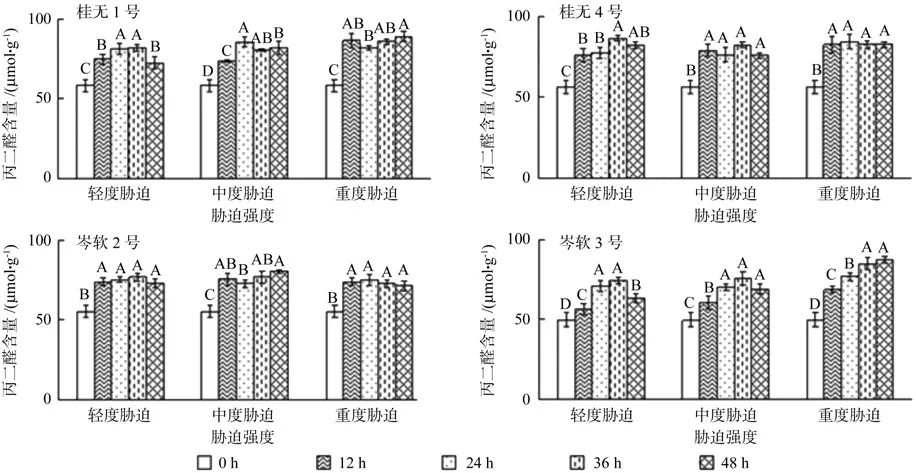
图5 不同干旱胁迫持续时间对各幼苗丙二醛含量的影响(不同字母表示差异性显著)Fig.5 Effect of drought stress on the content of MDA in leaves of seedlings
2.7 干旱胁迫对可溶性糖含量的影响
干旱胁迫环境条件下,4个油茶品种叶片可溶性糖含量均有变化,其中岑3的变化幅度最大(图7)。除桂1重度胁迫和桂4轻度胁迫可溶性糖含量增幅较明显外,该两个品种的其余处理均变化幅度较小,有的甚至出现下降;岑2各胁迫强度总体呈上升趋势,在24 h或36 h才有较明显增幅;岑3在各胁迫强度在12 h叶片可溶性糖含量即显著增加(P<0.05),中度和重度胁迫均出现双峰。复水12 h后,除桂1轻度和中度胁迫,岑2中度胁迫叶片可溶性糖含量显著升高外,其余品种各胁迫强度均下降。
2.8 幼苗抗旱性的综合评价
隶属函数综合评价的评价结果准确,可信度高,能综合评价植物的抗旱能力。为了研究影响4个油茶品种抗旱性的主要因素,作者采用隶属函数进行综合评价,叶片相对含水量、叶片相对电导率、叶绿素含量、POD活性、丙二醛含量、可溶性糖含量、可溶性蛋白含量这7种生理指标抗旱隶属函数分析结果见表2。
对4个油茶品种的各指标的隶属函数值进行累加计算并求平均数,并进行品种间抗旱性强弱比较。其中隶属函数值大于0.5的为岑2和岑3,即抗旱性较强;而岑2和岑3均隶属函数值小于0.5,则说明抗旱性较弱。结果表明,4个油茶品种的抗旱性顺序为岑3>岑2>桂1>桂4。
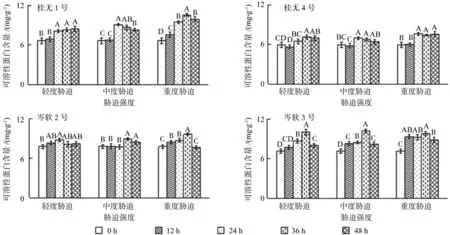
图6 不同干旱胁迫持续时间对各幼苗可溶性蛋白含量的影响(不同字母表示差异性显著)Fig.6 Effect of drought stress on the contents of soluble protein in leaves of seedlings
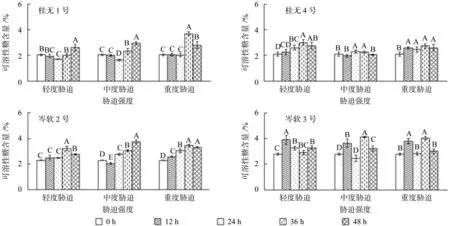
图7 不同干旱胁迫持续时间对各幼苗可溶性糖含量的影响(不同字母表示差异性显著)Fig.7 Effect of drought stress on the contents of soluble sugar in leaves of seedlings
3 结论与讨论
3.1 结 论
干旱胁迫会引起植物生理指标的变化,研究结果显示,在干旱胁迫的环境条件下,4个油茶品种幼苗的叶片相对含水量、叶片相对电导率、叶绿素含量、POD活性、丙二醛含量、可溶性蛋白含量、可溶性糖含量这7种生理指标均发生显著变化。其中,叶片相对含水量、叶绿素含量、POD活性、可溶性糖含量和可溶性蛋白含量这5个抗旱性正相关指标显示岑2和岑3的抗旱性较强,而叶片相对电导率、丙二醛含量这2个抗旱性负相关指标显示桂1和桂4的抗旱性较弱。复水处理后,岑2和岑3相关生理指标具有一定恢复能力,表明其抗旱性较强。
植物的抗旱性是一个复杂的综合性状,它受多种因素共同影响,多个抗旱相关生理指标的综合评价能够较全面反映植物的综合抗性。隶属函数对4个油茶品种幼苗的7 个生理指标进行综合评定显示,抗旱性排序为岑软3号 >岑软2号 >桂无1号 >桂无4号。
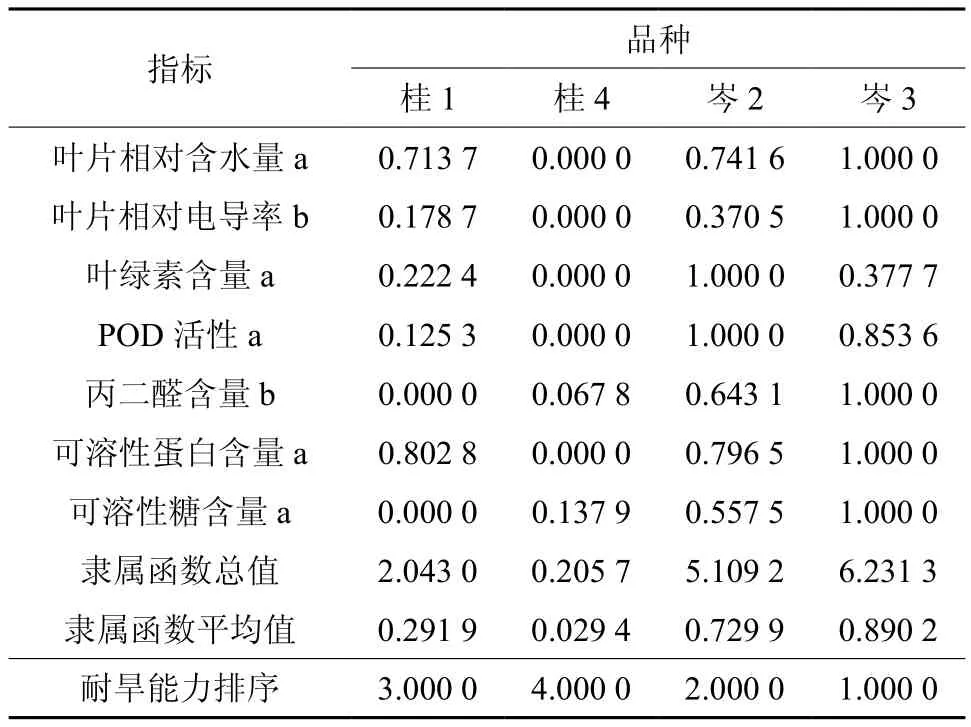
表 2 4个油茶品种抗旱性综合评定指数与排序†Table 2 Subordinate function and comprehensive judgment on drought resistance index
3.2 讨 论
水分是活细胞必要组分和重要代谢活动的物质,抗旱性强的树种在干旱环境下具有更强的持水能力,能减少体内水分丧失,维持正常生理活动[13]。干旱胁迫后4个油茶品种叶片的相对含水量均呈下降趋势,其中桂4降幅度最大,轻度胁迫的降幅已较其余3个品种重度胁迫更大,表明其抗旱能力较弱。而岑2和岑3,尤其是岑3的叶片相对含水量降幅最小,表明其抗旱能力较强。莫言玲等对西瓜12个基因型的抗旱性研究也表明,抗旱性强的基因型叶片相对含水量降幅小,抗旱性弱的基因型降幅大[14]。复水后,岑3叶片相对含水量上升,表明其在干旱胁迫后仍具一定恢复能力。
植物细胞在干旱条件下遭受氧化胁迫,质膜被破坏导致电解质外渗。本实验中,随着胁迫强度加深及时间延长,4个油茶品种叶片相对电导率整体呈上升趋势,这与前人研究结果相似[15-16]。在各胁迫强度下,桂4在12 h,桂1在24 h,岑2和岑3在36 h叶片相对电导率分别出现显著增幅,显示品种间抗旱性具较大差异,桂4抗旱性最弱,岑2和岑3抗旱性较强。与其余品种比较,岑3在不同胁迫强度下电导率变化幅度均较小,表明其抗旱性最强。复水后,桂4中度胁迫,岑2轻度胁迫,岑3各胁迫强度均有下降,表明其细胞膜具有一定功能恢复能力。
干旱胁迫能引起叶片叶绿素含量降低[17],进而导致光合作用下降。抗旱能力强的植物,能在相同干旱胁迫条件下保持较高的叶绿素含量。实验结果表明,随着干旱胁迫的持续,桂1和桂4叶绿素含量呈波动下降趋势,36 h已显著低于对照,可能是干旱胁迫引起了叶绿素分解。而岑2和岑3的36 h叶绿素含量均维持在较高水平且显著高于对照,可能干旱胁迫导致含水量下降出现绿叶素浓缩,也可能与植物对环境因子的补偿和超补偿效应有关[18]。复水后,岑2和岑3的叶绿素含量恢复或接近对照水平,与两个品种叶片含水量维持在较高水平密切相关。
POD是植物体内重要的抗氧化保护酶,逆境胁迫下其活性的增强能有效清除活性氧[19],保护细胞膜的完整性,对维持植物正常的生理活动具有重要意义[20]。本研究结果显示,岑2和岑3在轻度和重度胁迫12 h时POD活性已显著提升,表明POD的应激保护机制能快速启动,保护植株减少受干旱胁迫的损伤,抗旱性较强有利于植物抵御干旱胁迫。而桂1和桂4两个品种的POD活性增幅较小,显示其POD活性应变能力较差,植物抗旱性较弱。复水后,岑3各胁迫强度的POD活性回落至对照水平,表明该品种具有较强的调节能力。
MDA是膜脂过氧化作用的产物之一,其含量高低是细胞膜脂过氧化作用和植物受逆境胁迫损伤程度的重要指标[21]。随着干旱胁迫的加剧,4个油茶品种的MDA含量在胁迫初期即显著增加,表示干旱胁迫已引起了植株的膜脂过氧化反应,对其造成了损伤。桂1,桂4和岑2在胁迫初期MDA含量即大幅度增加,显示其损伤严重,抗旱性较弱;而岑3在轻度和中度胁迫12 h时,MDA含量增幅较小且明显低于其余3个品种,表明其受胁迫损伤较少,抗旱性较强。复水后,桂1和岑3轻度胁迫的MDA含量显著下降,表明其自身修复机制已启动,具有较好的恢复能力。
干旱胁迫导致植物体内可溶性蛋白含量增加,降低细胞的渗透势,从而增强其抗旱性[22],前人研究也证明可溶性蛋白的积累能力与植物抗旱性呈正相关[23]。4个油茶品种的可溶性蛋白含量随干旱胁迫时间的延长,总体呈现稳步升高趋势,这是植物对干旱胁迫的保护反应。但岑3可溶性蛋白增长较快且幅度明显,表明其快速启动相关渗透调节机制,抗旱性较强;而桂4可溶性蛋白增幅均最小,表明其调节能力较差,抗旱性较弱。复水处理后,4个油茶品种可溶性蛋白含量普遍下降,有可能是植物在非胁迫环境下对受损蛋白的自我修复或合成新蛋白。
可溶性糖是植物主要的渗透调节剂,能有效提高细胞的渗透浓度,降低细胞渗透势,维持体内细胞一定的膨压,抵御水分亏缺或促进吸水[24]。干旱胁迫期间,4个油茶品种叶片可溶性糖含量波动增加,这与武燕奇等的研究结果一致[25]。其中,岑3在12 h可溶性糖含量即显著升高,表明其能迅速调动可溶性糖合成生理,降低水势以维持高含水量,抗旱性较强;而其余品种增幅较小或增长较慢,很可能是其可溶性糖调控能力较弱,或者启动较慢,表明其抗旱性较弱。复水后,岑2轻度胁迫,桂4中度胁迫,桂1重度胁迫,以及岑3中度和重度胁迫含水量均显著下降,表明其具有一定调节能力。
在本研究中,4种油茶幼苗7个生理指标在不同干旱胁迫下发生变化,能够反映出不同品种间抗旱性的差异,且隶属函数对生理指标的综合评价能对4个油茶品种抗旱性差异进行科学排序。但是,与当前对林木抗旱性开展的大多数研究类似,本研究以1~2年生幼苗为试验材料,幼树与成年树的抗旱特性可能存在差异,是否表现一致还未经验证。后续对不同树龄苗木抗旱性的持续追踪和研究,能更深入、更全面地了解油茶品种的抗性,是今后研究开展的方向。
[1]钟飞霞,王瑞辉,李 婷,等. 土壤水分对油茶果实主要经济指标的影响[J]. 经济林研究,2015,33(4):32-37.
[2]张规富,孙航远. PEG6000模拟干旱对油茶幼苗形态结构的影响[J]. 湖北农业科学,2016,55(11):2829-2833.
[3]袁德梽. 水分胁迫对油茶容器苗生理特性的影响[J]. 山东农业大学学报(自然科学版),2015,46(4):509-513.
[4]许彦明,陈永忠,陈 勇,等.持续干旱对油茶叶片及根系生理生化指标的影响[J]. 湖南林业科技,2016,43(4):7-11.
[5]刁兆龙,陈 辉,冯金玲,等. 水分胁迫对油茶苗生理生化特性的影响[J]. 安徽农业大学学报,2014,41(4):642-646.
[6]张诚诚,曹志华,胡娟娟,等. 水分胁迫对油茶容器苗生理生化特性的影响[J]. 安徽农业大学学报,2013,40(4):623-626.
[7]周招娣,叶 航,马锦林,等. 油茶湘林11号无性系嫁接苗的抗旱性研究[J]. 中南林业科技大学学报,2015,35(2):54-58.
[8]霍佩佩,李小燕,林 萍,等.干旱胁迫对油茶优良无性系渗透调节物质和叶片水分状况的影响[J]. 内蒙古农业大学学报(自然科学版),2012,33(4):54-58.
[9]冯士令,程浩然,李 旭,等. 长林无性系油茶抗旱性的综合评价[J]. 广西植物,2016,36(6):735-740.
[10]陈建勋,王晓峰. 植物生理学实验指导(第二版)[M]. 广州:华南理工大学出版社,2006.
[11]Jiang Y W, Huang B R. Protein alterations in tall fescue in response to drought stress and abscisic acid[J]. Crop Science,2002, 42(1): 202-207.
[12]梁丹妮, 郭兴燕,兰 剑. 6份沿阶草种质对干旱胁迫的生理响应[J].草业科学,2016,33(2):184-191.
[13]Hayano-Kanashiro C, Calderón-Vázquez C, Ibarra-Laclette E, et al. Analysis of gene expression and physiological responses in three Mexican maize landraces under drought stress and recovery irrigation[J]. PLoS One, 2009, 4(10): e7531.
[14]莫言玲,郑俊鶱,杨瑞平,等. 不同西瓜基因型对干旱胁迫的生理响应及其抗旱性评价[J]. 应用生态学报,2016, 27(6):1942-1952.
[15]唐洪辉,魏 丹,赵 庆,等. 干旱胁迫对宫粉羊蹄甲生理指标的影响[J]. 中南林业科技大学学报,2017, 37(4): 7-13.
[16]Liu H, Wang Y, Zhou X, et al. Overexpression of a harpinencoding gene popW from Ralstonia solanacearum primed antioxidant defenses with enhanced drought tolerance in tobacco plants[J]. Plant cell reports, 2016, 35(6): 1333-1344.
[17]Kyparissis A, Petropoulou Y, Manetas Y. Summer survival of leaves in a soft-leaved shrub (Phlomis fruticosa L., Labiatae) under Mediterranean field conditions: avoidance of photoinhibitory damage through decreased chlorophyll contents[J]. Journal of Experimental Botany, 1995, 46(12): 1825-1831.
[18]冯慧芳,薛 立,任向荣,等. 4 种阔叶幼苗对PEG模拟干旱的生理响应[J]. 生态学报, 2011, 31(2): 371-382.
[19]Anjum S A, Xie X, Wang L, et al. Morphological, physiological and biochemical responses of plants to drought stress[J]. African Journal of Agricultural Research, 2011, 6(9): 2026-2032.
[20]李 娜,罗俊杰,张仁陟,等. 持续模拟干旱胁迫对胡麻萌发特性影响及品种抗旱性评价研究[J]. 核农学报,2016,30(2):379-387.
[21]Catola S, Marino G, Emiliani G, et al. Physiological and metabolomic analysis of Punica granatum (L.) under drought stress[J]. Planta, 2016, 243(2): 441-449.
[22]邵怡若,许建新,薛 立,等. 5种绿化树种幼苗对干旱胁迫和复水的生理响应[J]. 生态科学, 2013,32(4): 420-428.
[23]李 君,周守标,王春景,等. 野生和栽培马蹄金抗旱性比较及其抗旱机制初探[J]. 植物生态学报, 2007,31(3): 521-527.
[24]梁文斌,聂东伶,吴思政,等. 水分胁迫对短梗大参生理生化特征的影响[J]. 经济林研究,2016,34(3):99-104.
[25]武燕奇,郭素娟. 10个板栗砧木品种 (系) 抗旱性综合评价[J].东北农业大学学报, 2016, 47(10): 9-16.
