安息香属3个树种苗木抗寒生理差异研究
2018-03-19高振洲李洁宁喻方圆
高振洲 ,李洁宁 ,喻方圆
(南京林业大学 a.林学院;b.南方现代林业协同创新中心,江苏 南京 210037)
安息香属植物是安息香科中比较特别的一个属,因其树干受到外部伤害后会流出树脂样物质而得名,具有重要的药用、观赏和经济价值[1]。红皮树Styrax suberifolius是名贵家具、室内装修、胶合板等优良用材,同时也是造纸的好材料;其种子油可制肥皂和油漆,是优良的能源树种之一[2];其树形雅致,可作为园林观赏树种。东京野茉莉Styrax tonkinensis生长速度快,成材期短,具有材质轻软,木纤维长的特点,是制浆、造纸等短周期工业原料林的首选树种之一[3];同时它还是一种应用前景非常广阔的生物柴油原料。此外,东京野茉莉的花朵白色成串,清香美丽,在园林绿化应用方面极具开发价值[4-5]。东京野茉莉也是良好的保持水土的树种,对改善环境和重塑良好人工生态系统方面具有重要意义[6]。垂珠花Styrax dasyanthus是一种具有较高利用价值的观花、观果、药用及油料资源型芳香植物,具有很大的综合开发利用价值及广阔的园林应用前景。
上述3个树种在我国长江以南地区均有分布,但由于起源不同,其抗寒能力存在差异。然而,关于安息香属不同树种抗寒性差异的研究还鲜有报道。Lenahan和Graves对2个美洲安息香S. americanus居群的光合特性进行了对比研究,发现美洲安息香对温度的变化有极强的适应能力,并认为该种能成功跨域引种,之后他们选取了三个 S. americanus 居群样本探讨升温期茎干的耐寒性情况,最后得出伊利诺伊州居群的种植资源可运用于较寒冷的地区[7]。
目前,对安息香科的长果安息香的研究较多[8-10]。而抗寒性研究方面,苹果叶片解剖结构与抗寒性的关系[11],扁桃花蕾抗寒性的研究等已有报道[12],而安息香属抗寒性的研究还未见报道。为了充分开发利用安息香属植物资源,确定东京野茉莉、垂珠花和红皮树三个树种的适宜引种栽培区域,本文对东京野茉莉、垂珠花和红皮树这3个树种苗木在自然越冬条件下的抗寒性进行了研究,测定了自然越冬过程中苗木体内的可溶性糖,可溶性蛋白质,脯氨酸和游离氨基酸的含量以及相对电导率的变化,分析上述生理指标与自然低温的关系,探讨三个树种苗木抗寒性之间存在的差异,旨在为东京野茉莉、垂珠花和红皮树的引种驯化提供理论和实践依据。
1 材料和方法
1.1 试验地概况及供试材料
试验地设在南京市六合区马集镇中华茉莉谷苗圃基地。该基地地处北纬 32°11′~ 32°27′,东经 118°34′~ 119°03′,全区属北亚热带季风温湿气候区,气候温和,雨量充沛,光照充足,四季分明,常年平均气温15.1 ℃,一般年份冬季极端最低气温在-8℃~-10℃,年平均降水量1 000 mm左右,年平均日照时数为2 199.4 h以上。
试验材料为东京野茉莉(江西吉水种源),垂珠花(湖南衡阳种源)和红皮树(湖南衡阳种源)的一年生苗木。所有苗木在南京市六合区马集镇中华茉莉谷苗圃基地播种培育,采用相同的管理条件。
2015年12月至2016年2月期间,每月15日采集枝条1次,分别采集3个树种同一朝向上的一年生苗木的侧枝,每个树种3个重复,每个重复10株,3个树种共90株。采集的枝条放在自封袋中带回实验室,用自来水和去离子水洗涤擦干后,先随机取部分样品测定相对电导率,然后将每树种每重复的所有枝条混合磨碎,从中随机取相应重量的样品分别测定可溶性糖含量、可溶性蛋白质含量、脯氨酸含量及游离氨基酸含量,每个指标重复3次。
1.2 生理指标的测定
可溶性糖含量的测定采用蒽酮比色法[13]。可溶性蛋白含量的测定采用考马斯亮蓝法[14]。游离氨基酸含量的测定采用茚三酮比色法[15]。脯氨酸含量的测定采用酸性茚三酮比色法[16]。
相对电导率使用DDS-307型电导仪进行测量,以去离子水电导率为对照,然后按下式计算相对电导率(Y):
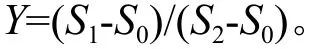
式中:Y为相对电导率;S1为初电导值;S2为终电导值;S0为去离子水电导值。
1.3 统计分析方法
用Excel软件汇总整理数据并制作图表,使用SPSS19.0对东京野茉莉、垂珠花和红皮树的可溶性糖含量、可溶性蛋白质含量、游离氨基酸含量、脯氨酸含量及相对电导率的数据进行单因素方差分析,计算标准误和进行多重比较。多重比较的显著性用p值表示,当p<0.01时,存在极显著差异;当p<0.05时,存在显著差异;当p>0.05时,差异不显著。
2 结果与分析
2.1 可溶性糖含量的比较
从图1可以看出,越冬过程中随着时间的推移红皮树枝条内的可溶性糖含量呈现先下降后平缓上升的趋势;东京野茉莉枝条内的可溶性糖含量随着时间的变化呈现下降的趋势;垂珠花枝条内的可溶性糖含量随着时间的变化呈现下降趋势。其中,红皮树枝条内可溶性糖含量在12月份最高,为34.710 mg/g,在1月份含量最低,为20.385 mg/g,下降了约41.3%;东京野茉莉枝条内可溶性糖含量在12月份最高,为93.690 mg/g,在2月份含量最低,为21.773 mg/g,约下降了76.8%;垂珠花枝条内可溶性糖含量在12月份最高,为58.450 mg/g,在2月份含量最低,为36.402 mg/g,下降了约37.7%。在自然越冬结束后,2月份,垂珠花枝条内可溶性糖含量均高于东京野茉莉和红皮树,分别高约67.2%、66.6%。根据方差分析可知,12月份(p=0.011),1月份(p=0.024)3种树种枝条内可溶性糖含量差异显著,2月份(p=0.076)可溶性糖含量差异不显著;多重比较结果如图1所示。
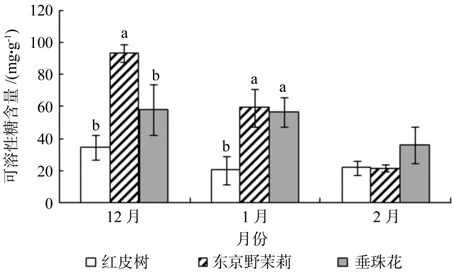
图1 越冬过程中3个树种枝条内可溶性糖含量的变化Fig.1 Changes of soluble sugar content detected in seedling twigs of Styrax suberifolius, Styrax tonkinensis and Styrax dasyanthus during the period of natural overwintering
2.2 可溶性蛋白质含量的比较
从图2可以看出,越冬过程中红皮树、东京野茉莉和垂珠花枝条内可溶性蛋白质含量随着时间的变化其变化趋势一致,均呈现上升趋势,但红皮树变化平缓。红皮树枝条内可溶性蛋白质含量在12月份最低,为2.041 mg/g,在2月份最高,为2.209 mg/g,增幅约为8.2%;东京野茉莉枝条内可溶性蛋白质含量在12月份最低,为3.185 mg/g,在2月份最高,为5.754 mg/g,增幅约为80.7%;垂珠花枝条内可溶性蛋白质含量在12月份最低,为4.910 mg/g,在2月份最高,为7.382 mg/g,增幅约为50.3%。在越冬期间枝条内的可溶性蛋白质含量:垂珠花>东京野茉莉>红皮树。根据方差分析可知,12月份(p=0.001),1月份(p=0.000),2月份(p=0.000)3种树种枝条内可溶性蛋白质差异极显著;多重比较结果如图2所示。
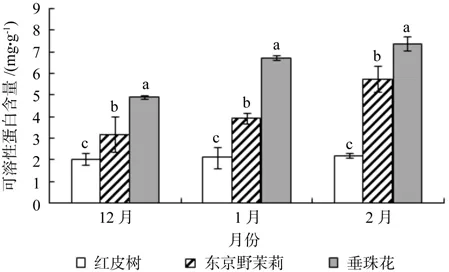
图2 越冬过程中3个树种枝条内可溶性蛋白质含量的变化Fig.2 Changes of soluble protein content detected in seedling twigs of Styrax suberifolius, Styrax tonkinensis and Styrax dasyanthus during the period of natural overwintering
2.3 脯氨酸含量的比较
从图3可以看出,越冬过程中红皮树枝条内脯氨酸含量随着时间的变化呈现下降趋势,其含量在12月份最高,为0.423 mg/g,在2月份最低,为0.155 mg/g,约下降了63.4%;东京野茉莉枝条内脯氨酸含量随着时间的变化呈现先上升后下降的趋势,但12月份与1月份脯氨酸含量差异不大,枝条内脯氨酸含量在1月份最高,为0.077 mg/g,在2月份含量最低,为0.042 mg/g,约下降了45.5%;垂珠花枝条内脯氨酸含量随着时间的变化呈现先上升后下降的趋势,但1月份与2月份脯氨酸含量差异不大,枝条内脯氨酸含量在12月份最低,为0.036 mg/g,在1月份最高,为0.079 mg/g,增幅约为119.5%。在越冬过程中红皮树枝条内脯氨酸含量高于东京野茉莉和垂珠花。由方差分析可知,12月份(p=0.000),1月份(p=0.005),2月份(p=0.000)3种树种枝条内脯氨酸差异极显著;多重比较结果如图3所示。
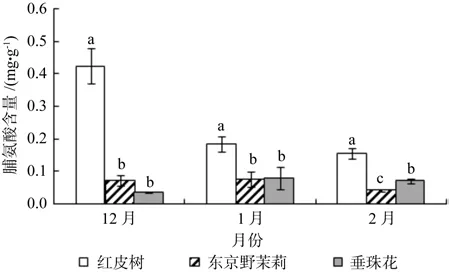
图3 越冬过程中3个树种枝条内脯氨酸含量的变化Fig.3 Changes of proline content detected in seedling twigs of Styrax suberifolius, Styrax tonkinensis and Styrax dasyanthus during the period of natural overwintering
2.4 游离氨基酸含量的比较
从图4中可以看出,越冬过程中红皮树和东京野茉莉枝条内游离氨基酸含量随着时间的变化其变化趋势一致,均呈现上升趋势。红皮树枝条内游离氨基酸含量在12月份最低,为3.194 mg/g,在2月份最高,为11.554 mg/g,增幅约为261.7%;东京野茉莉枝条内游离氨基酸含量在12月份最低,为0.785 mg/g,在2月份最高,为3.041 mg/g,增幅约为287.4%。垂珠花枝条内游离氨基酸含量呈现先上升后下降的趋势,在12月份最低,为3.192 mg/g,在1月份最高,为6.639 mg/g,增幅约为108.0%。由方差分析可知,12月份(p=0.000),1月份(p=0.000),2月份(p=0.000)3种树种枝条内游离氨基酸差异极显著;多重比较结果图4所示。
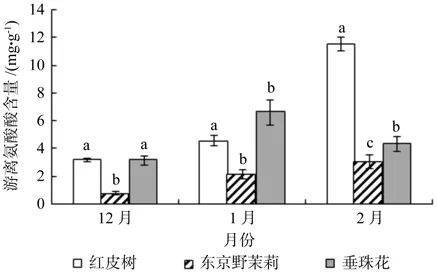
图4 越冬过程中3个树种枝条内游离氨基酸含量的变化Fig.4 Changes in content of free amino acids detected in seedling twigs of Styrax suberifolius, Styrax tonkinensis and Styrax dasyanthus during the period of natural overwintering
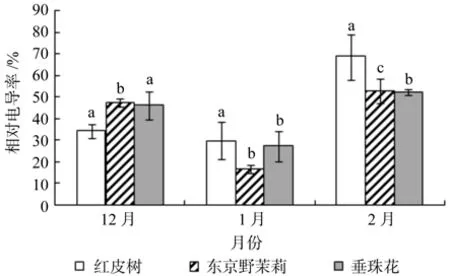
图5 越冬过程中3个树种枝条相对电导率的变化Fig.5 Changes of relative electrical conductivities detected in seedling twigs of Styrax suberifolius, Styrax tonkinensis and Styrax dasyanthus during the period of natural overwintering
2.5 相对电导率的比较
从图5中可以看出,越冬过程中红皮树、东京野茉莉和垂珠花枝条的电导率随着时间的变化其变化趋势一致,均呈现“高-低-高”的趋势。红皮树、东京野茉莉和垂珠花枝条的电导率均在1月份最低,分别为29.8%、16.7%、27.3%,在2月份最高,分别为68.7%、53.0%、52.4%,增幅分别约为130.5%、217.4%、91.9%。由方差分析可知,12月份(p=0.020)3种树种枝条相对电导率差异显著,1月份(p=0.121)和2月份(p=0.211)枝条相对电导率差异不显著;多重比较结果图5所示。
3 讨论与结论
3.1 可溶性糖与抗寒性
可溶性糖是一种低温保护物质,具有渗透调节的功能,同时糖还是冰冻保护剂,对原生质体、线粒体及膜上敏感偶联因子均有保护作用[17]。姜卫兵等发现,从秋末进入冬季,无花果枝皮可溶性糖含量逐渐提高[18]。马翠兰等对渗透调节物质和水分状态与琯溪蜜柚抗寒性的关系进行研究证实,可溶性糖含量从11月份到翌年1月份呈增加趋势[19]。但也有研究与之有差异,Li et al.的研究结果显示,在越冬之前,尤其是在11月间,豆科牧草根积累的可溶性糖最多,此时正值进入越冬状态[20]。
本实验结果与后者较为相似,红皮树、东京野茉莉和垂珠花枝条内的可溶性糖含量最高值均出现在12月,此时苗木逐渐进入越冬状态,苗木积累了大量的可溶性糖,说明已做好了越冬准备。1月份气温进一步降低,可溶性糖含量反而下降,这可能是由于1月份,进入深休眠阶段,为了抵抗寒冷消耗掉了一部分可溶性糖。在1月—2月间,经过一段时间适应后,3个树种枝条内糖含量出现回升,表明低温适应后,植物体内存在某种机制,调节体内糖含量的增加以增加其对低温的抵抗能力,但适应后可溶性糖的含量比初始的含量低,这可能是冬季3个树种不能补充新的光合产物所导致的。越冬结束后,垂珠花枝条内的可溶性糖含量积累是最多的,整个越冬过程中,红皮树枝条内可溶性糖含量整体水平是3个树种中最低的,这在一定程度上说明垂珠花苗木抗寒性较强,红皮树苗木抗寒性较弱。因为“可溶性糖的含量与抗寒性存在平行现象,抗寒性强的树种,可溶性糖的含量高”[21]。戴文浩等对果梅的研究也表明,可以用越冬期间可溶性糖含量的高低来推测品种间抗寒性的差异[22]。由此可见,可溶性糖含量的变化与高低,可以作为评价红皮树、东京野茉莉和垂珠花抗寒性强弱的一个重要指标。
3.2 可溶性蛋白质与抗寒性
可溶性蛋白质具有渗透调节、保水及降低冰点的作用。此外,部分可溶性蛋白质还是功能蛋白酶,低温抑制植物的蛋白质合成会降低植物的抗寒性。多数研究者通过实验研究认为:低温胁迫下,植物的可溶性蛋白含量会增加[23-25]。也有研究发现,有一些植物可溶性蛋白质含量在抗冷锻炼中并没有实质性的改变[26]。
本实验通过对越冬过程中红皮树、东京野茉莉和垂珠花可溶性蛋白含量的测定,发现在12月—翌年2月间红皮树、东京野茉莉和垂珠花苗木枝条内可溶性蛋白含量均呈现上升趋势,说明3个树种苗木枝条内的可溶性蛋白对温度的变化特别敏感,其可溶性蛋白含量的提高极有可能是因为低温诱导,植物体通过蛋白合成尽快适应低温环境。12月—翌年1月间,垂珠花枝条可溶性蛋白含量上升幅度最大,约上升了37.0%,东京野茉莉枝条内可溶性蛋白含量约上升了23.5%,红皮树枝条内可溶性蛋白含量约上升了3.4%。另外,在整个越冬期间,3个树种枝条内可溶性蛋白质含量的高低为:垂珠花>东京野茉莉>红皮树,这可能在一定程度上说明3个树种苗木抗寒性强弱为:垂珠花>东京野茉莉>红皮树,因为很多报道都认为,抗寒性强的树种其可溶性蛋白含量明显高于抗寒性弱的树种。
3.3 脯氨酸与抗寒性
许多研究认为,大部分植物也会通过脯氨酸含量的增加抵御外界低温,因为脯氨酸不但起到平衡细胞代谢的作用,而且还能保持细胞环境的稳定,由此可见,脯氨酸含量的增加对于抗寒性具有积极的意义。也有研究证明,低温胁迫下脯氨酸含量的增加通过降低细胞水势来避免冻害的发生[27-28]。卢少云等在研究低温下的狗牙根时发现脯氨酸含量的增加与植物受伤害程度呈显著正相关,以说明脯氨酸的积累是低温引起的伤害反应[291]。池春玉等也做了不少关于抗寒性的研究,如对3种冷季型草坪草进行低温胁迫处理,研究低温对脯氨酸含量和膜透性的影响,发现低温胁迫下3种草坪草叶片内脯氨酸含量都发生了极显著的变化,即脯氨酸含量均呈现上升的走势[30]。但也有研究认为脯氨酸的累积与抗寒性并不存在任何关系[31]。
本实验对自然越冬条件下红皮树、东京野茉莉和垂珠花枝条内脯氨酸含量的研究发现,红皮树、东京野茉莉和垂珠花枝条内脯氨酸含量变化没有表现出一致的规律性,而且东京野茉莉、垂珠花和红皮树三个树种中脯氨酸含量存在一定的差别,这可能是不同植物脯氨酸的合成对低温影响机制的反应不同造成的,另外一种原因可能是脯氨酸含量的积累只不过是胁迫条件下的一种适应性反应,并不是抵抗低温的途径,或者是不同植物不一定通过同一途径来抵抗外界的不良环境。
3.4 游离氨基酸与抗寒性
在正常条件下,植物体内游离氨基酸含量很低,而低温胁迫条件下,游离氨基酸的含量迅速上升。王英等研究发现,低温处理后山定子和平邑甜茶根系游离氨基酸含量明显上升[32]。栗淑媛等研究发现,螺旋藻在遇低温状况时其蛋白质代谢会发生变化,部分蛋白质水解大于合成,使细胞内游离氨基酸含量升高[33]。
在本实验中,红皮树、东京野茉莉和垂珠花枝条内游离氨基酸含量在12月份较低,在12月—翌年1月间随着温度的降低游离氨基酸含量迅速增加,且垂珠花枝条内游离氨基酸的增长幅度和含量均高于东京野茉莉和红皮树。游离氨基酸含量的增加有效地起到保护原生质的能力,降低冰点,使植物顺利越冬。
3.5 相对电导率与抗寒性
植物细胞膜是细胞重要的组成部分,起着保护细胞和调节控制细胞内外物质交流的屏障作用,低温胁迫对植物伤害的最初部位与主要部位便是膜系统[34]。低温胁迫直接导致细胞膜透性增大,从而使细胞内的电解质有不同程度的外渗,电导率因而上升。电导率能反映植物在胁迫条件下生物膜透性的变化情况。而且,不同树种的相对电导率间有显著的差异[35]。因此,通过测定组织浸出液电导率能看出植物组织受伤害的程度,相对电导率值便成为间接评价植物抗寒性的一种有效手段。
在本实验中,红皮树、东京野茉莉和垂珠花相对电导率在越冬过程中整体呈现先下降后上升的变化趋势。因为第一次采样时间在12月中旬之后,此时3个树种苗木的都经过了一定时间的低温锻炼,自然会对低温产生适应性。但当胁迫加深时,超过了苗木组织承受的限度,细胞膜开始受到严重破坏,电解质开始外渗,电导率上升,因而又呈现出上升趋势。经过自然越冬后,红皮树相对电导率为68.7%,东京野茉莉为53.4%,垂珠花为52%。这说明垂珠花苗木的细胞受损程度轻于东京野茉莉和红皮树,这与徐晓薇等[36]研究发现抗寒性强的树种电解质渗出相对较少相吻合,所以相对电导率可以作为判断红皮树、东京野茉莉和垂珠花抗寒性强弱的一个重要指标。
本次实验还存在不足之处,实验的时间梯度设置过少,应该以10 d或者15 d为一个时间梯度进行设置,以便得到更加可靠的结论。为了进一步研究安息香属东京野茉莉、垂珠花和红皮树的苗木的抗寒生理差异,在接下来的实验中,我会对东京野茉莉、垂珠花和红皮树的苗木进行人工低温胁迫的处理,得到低温胁迫下的3个树种苗木的抗寒生理差异。
3.6 结 论
通过对红皮树、东京野茉莉和垂珠花3个安息香属树种的苗木抗寒生理差异研究发现,在越冬结束后,垂珠花枝条内的可溶性糖含量积累是最多的;而整个越冬过程中,红皮树枝条内可溶性糖含量整体水平是3个树种中最低的。在整个越冬期间,3个树种枝条内可溶性蛋白质含量的高低及上升幅度的大小均为:垂珠花>东京野茉莉>红皮树。在12月—翌年1月间,垂珠花枝条内游离氨基酸的增长幅度和含量均高于东京野茉莉和红皮树。经过自然越冬后,垂珠花苗木的细胞受损程度轻于东京野茉莉和红皮树。根据以上生理指标进行的综合分析,可以初步判断出,垂珠花的抗寒性是这3个树种中最强的,东京野茉莉次之,红皮树的抗寒性最弱。
[1]郑万钧.中国树木志:第2卷[M].北京:中国林业出版社,1985.
[2]叶秀磷,钱南芬,王伏雄.红皮树胚胎发育[J].云南植物研究, 1986,8(2):125-131.
[3]杨 桦,唐仕斌,岳军伟,等.优良生物质能源树种东京野茉莉研究进展[J].安徽农业科学,2012,40(36):17648-17649.
[4]傅立国,陈潭清.中国高等植物: 第六卷[M]. 青岛:青岛出版社,2003.
[5]肖复明,曾志光,杨 桦,等.东京野茉莉种子油营养成分研究[J].天然产物研究与开发,2005,17(3):344-345.
[6]戴晓龙.东京野茉莉及繁育技术[J].特种经济动植物,2002,5(6):21.
[7]Lenahan O M, Graves W R, Arora R. Cold-hardiness and deacclimation of Styrax americanus from three provenances[J].Hortscience, 2010, 45(12): 1819-1823.
[8]黄 滔,唐 红,廖菊阳,等.长果安息香夏季光合蒸腾日变化与其环境因子的关系[J].中南林业科技大学学报,2015,35(7): 62-68.
[9]张 程,彭重华,刘 晖,等.湘西北长果安息香种群数量与分布格局[J].湖南农业大学学报(自然科学版),2011,37(6):615-620.
[10]张 程,彭重华,刘 晖,等.湘西北长果安息香群落区系组成与特征[J].南京林业大学学报(自然科学版),2012,36(3):144-148.
[11]王泽华,秦 伟,闫娟娟,等.不同生境新疆野苹果叶片解剖结构及其与抗寒性的关系[J].经济林研究,2016,34(3):163-168.
[12]李 鹏,田 嘉,唐开文,等.基于隶属函数评估法的扁桃花蕾抗寒性研究[J].中南林业科技大学学报,2017, 37(2):39-43.
[13]陈秀兰.蒽酮比色法测定木薯块根的淀粉[J].分析化学,1984,12(4): 319.
[14]宿 玮,常耀光,薛长湖,等. 海地瓜多糖中蛋白含量测定方法比较[J]. 食品科学,2011,32(2):201-204.
[15]邵金良,黎其万,董宝生,等. 茚三酮比色法测定茶叶中游离氨基酸总量[J]. 中国食品添加剂,2008(2):162-165.
[16]李绍军,龚月桦,王俊儒,等. 关于茚三酮法测定脯氨酸含量中脯氨酸与茚三酮反应之探讨[J]. 植物生理学通讯,2005,41(3): 365-368.
[17]张德基,李玉梅,陈艳秋,等.梨品种枝条可溶性糖、脯氨酸含量变化规律与抗寒性的关系[J].延边大学农学学报,2004,26(4):281-285.
[18]姜卫兵,王业遴,马 凯.渗透保护物质在无花果抗寒性发育中的作用[J].园艺学报,1992,19(4):371-372.
[19]马翠兰,刘星辉,胡又厘.柚品种间的耐寒性差异及其机理[J].福建农业大学学报, 1998,27(2):33-38.
[20]LiR, Volenec JJ, Joern BC, et al. Seasonal changes in nonstructural carbohydrates, protein and macronutrients in roots of alfalfa, red clover, sweet clover and birdsfoot trefoil[J].Crop Science, 1996, 36:617-623.
[21]何开跃,李晓储,黄利斌,等.3种含笑耐寒生理机制研究[J].南京林业大学学报(自然科学版),2004, 28(4):62-64.
[22]戴文浩,章 镇,赵培华,等.果梅若干越冬生理指标的研究[J].果树科学,1997,14(4):248-251.
[23]刘祖棋,王洪春.植物耐寒性及防寒技术[M].北京:学术书刊出版社,1990.
[24]李荣富,王丽雪,张 华.果树抗寒性的细胞生物学研究进展[J].北京农学学报,1996,11(2):79-83.
[25]刘鸿先,王以柔,曾韶西,等.低温对不同耐冷力的黄瓜幼苗呼吸代谢的影响[J].植物生理学报,1984,10(3):191-199.
[26]Sari K.S, Jnane L., and Pekka L. Response of protein and carbohydrate metabolism of Scot pine seedlings to low temperature[J]. Plant Physiology, 2002, 159(2): 175-180.
[27]龚 明.作物抗性鉴定方法与指标及其综合评价[J].云南农业大学学报, 1989,4(1):73-78.
[28]陈杰忠,徐春香,梁立峰.低温对香蕉叶片中蛋白质即脯氨酸的影响[J].华南农业大学报,1999,2(3):54-58.
[29]卢少云,郭振飞,陈永传.狗牙根的组织培养及其矮化变异体研究初报[J].园艺学报,2003,30(4):482-484.
[30]池春玉,丁国华,连永权,等.低温胁迫对三种冷季型草坪草脯氨酸含量及膜透性的影响[J].中国农学通报,2007, 23(1):101-104.
[31]梅俊学.逆境下发菜脯氨酸含量及质膜透性的变化与含水量的关系[J].山东师范大学学报,2000,15(2):178-181.
[32]王 英,吕德国,秦嗣军,等.低温对山定子和平邑甜茶幼苗根系氮代谢酶及游离氨基酸的影响[J].园艺学报,2010, 37(2):179-184.
[33]栗淑媛,马花山,王俊琴,等.低温对螺旋藻可溶性蛋白质和游离氨基酸外渗的影响[J].西北植物学报,2005,25(6):1195-1198.
[34]马英姿,梁文斌,陈建华.经济植物的抗寒性研究进展[J].经济林研究,2005,23(4):89-94.
[35]刘 霞,侯乐峰,郝兆祥,等.以电导法评价12个石榴品种的抗寒性[J].经济林研究,2015, 33(3):150-153.
[36]徐晓薇,林绍生,曾爱平.蝴蝶兰抗寒力鉴定[J].浙江农业科学,2004(5):249-251.
