Physiological response of four wolfberry (Lycium Linn.) species under drought stress
2018-03-07ZHAOJianhuaLIHaoxiaZHANGCunzhiANWeiYINYueWANGYajunCAOYoulong
ZHAO Jian-hua, LI Hao-xia, ZHANG Cun-zhi, AN Wei, YIN Yue, WANG Ya-jun, CAO You-long
1 National Wolfberry Engineering Research Center, Ningxia Academy of Agriculture and Forestry Sciences, Yinchuan 750002,P.R.China
2 Desertification Control Research Institute, Ningxia Academy of Agriculture and Forestry Sciences, Yinchuan 750002, P.R.China
3 Ningxia Professional Technology College, Yinchuan 750021, P.R.China
1.Introduction
Arid and semiarid zones have traditionally contributed around 40% of the total worldwide production of food grains(Bhatt et al.2011).However, severe water deficits, or droughts, can bring devastating effects on crop productivity in these areas.Drought stress can arrest plant growth and reduce its distributive area (Diego et al.2012).In order to survive under drought stress conditions, plants can reduce leaf water potential, stomatal conductance (Gs), and gasexchange, and may slow normal growth rates (Reddy et al.2004; Ribas-Carbo et al.2005; Deng et al.2012).
Drought-induced stomatal closure may increase the oxidative load on plant tissues, causing perturbations in biochemical pathways, if the accumulation of excessive reactive oxygen species (ROS) is not stopped.Oxidative stress causes lipid peroxidation and damage to other important biomolecules (Bhatt et al.2011).In addition to morphological adaptations (e.g., extending deep roots to tap ground water and closing stomata to reduce water loss),plants have demonstrated a series of physiological and biochemical mechanisms to minimize drought stress.These internal responses range from changes in photosynthetic activity to the development of antioxidant defenses to enhance drought tolerance (Gao et al.2009).
The earliest response to drought seems to be stomatal closure, and both stomatal and mesophyll conductance are the primary limitations to photosynthesis during mild to moderate drought stress.Drought results in the inhibition of photosynthesis, either due to diffusion limitations or metabolic impairment (Ávila et al.2012).Plants protect themselves from drought-induced oxidative damage by producing an array of anti-oxidative enzymes such as superoxide dismutase (SOD), catalase (CAT), guaiacol peroxidase (GPX), and ascorbate peroxidase (APX), which act synergistically to limit levels of ROS (Bhatt et al.2011).
Wolfberry species (Lycium Linn.) are perennial,deciduous shrubs that grow in Northwest China and the Mediterranean region.These shrubs possess fast-growth characteristics, a well-developed deep root system, and extensive adaptability to drought and cold (Chen et al.2004;Chang and So 2008).The fruit of Lycium barbarum L.(family Solanaceae, also named fructus lycii or wolfberry) has been used for centuries in China as a traditional herbal medicine and as a valuable nourishing tonic (CCP 2010).Recently,medical research has indicated that these fruits have many pharmacological functions, such as improving visual acuity,maintaining liver and kidney function, reducing blood sugar levels, reducing the risk of cancer and cell senescence, and improving immune function (Xu et al.2000; Xie et al.2001;Chang and So 2008).
The depth of existing research on water-stress tolerance in plants varies depending on plant species.Although the general effects of drought on plant growth are fairly well known (Chaves 1991; Wang et al.2011), the essential consequences of drought stress at the physiological level have not been thoroughly studied for wolfberry species (Lycium Linn.).This study was aimed to research physiological responses of drought tolerance in four wolfberry species under drought stress.The observations will be helpful for developing high-yield cultivation techniques for wolfberry as well as breeding varieties for drought tolerance.
2.Materials and methods
2.1.Plant materials and growth conditions
The experiment was conducted in a greenhouse at the Wolfberry Germplasm Repository of the Ningxia Academy of Agriculture and Forestry Sciences, China (38°38´N,106°9´E).The area has a typical temperate continental climate, with an average temperature of 8.5°C, annual average sunshine between 2 800 and 3 000 h (one of the largest amounts of solar radiation and hours of sunshine in China), and an annual average precipitation of 200 mm,most of which falls during the summer.
The four wolfberry species used in the study were Lycium chinense Mill.var.potaninii (Pojark.) A.M.Lu, Lycium chinense Mill., Lycium barbarum L., and Lycium yunnanense Kuang & A.M.Lu, and the seedlings were collected from one 10-year-old tree, respectively.Semi-lignified branches were collected, and 8-cm hardwood cuttings were taken from the branches with uniform diameters and one or more lateral buds.Cuttings of each clone were collected from stool beds growing at the greenhouse in September 2012.In April 2013, germinating, healthy, and uniform seedlings were transplanted individually to plastic pots (32-cm in diameter and 28-cm in height) containing sand, soil, and vermi-compost (1:3:1, v/v).The mixture weighed about 10 kg per pot and the saturated water content was 24% by mass.All seedlings were grown in a full sunlight greenhouse under a semi-controlled environment (only sheltered from rainfall) during the experiment.
2.2.Experimental design
The experiment was arranged in a completely randomized design for four water supply regimes (75, 55, 35, and 15%field capacity (FC)), of which 75% FC served as the control,and the others served as light, moderate, and severe water drought conditions, respectively.There were 20 pots for each treatment.Watering treatments started on 15 June,2013.Following natural water loss, all of the water supply regimes reached the required FC by 25 June.After reaching the target FC, the pots received compensatory irrigation by weight at 18:00 everyday during the experiment to maintain a constant soil moisture level.The treatment lasted about 2 months.All measurements were taken 50-60 days after drought.
2.3.Chlorophyll and carotenoid pigments
About 0.5 g of fresh leaf tissue was used for each extraction.Leaf tissue was cut into small pieces,placed in a test tube, extracted in 80% acetone at room temperature, and left on a rotary shaker for 24 h in the dark.The absorbance of the extracts was measured on a spectrophotometer (TU-1900, Beijing, China) at 470, 646, and 663 nm wavelengths, respectively.The concentrations of chlorophylls and carotenoids were calculated according to the method of Inskeep and Bloom(1985) and Arnon (1949), respectively.
2.4.Gas-exchange
Net photosynthesis rate (Pn), transpiration rate (E), and Gswere measured using the CI-340 (CID Inc., Camas, WA,USA) on sunny days from 9:00 to 11:00 in the morning.Leaf gas-exchange was measured on the 5th to the 6th recent mature leaves in the middle of the branch from 5-6 seedlings under treatment conditions.Instantaneous water use efficiency (WUE) was calculated as:
WUE=Pn/E
Where, unit of WUE is μmol CO2mmol-1H2O; unit of Pnis μmol CO2m-2s-1; unit of E is μmol H2O m-2s-1.
2.5.Chlorophyll fluorescence
Chlorophyll fluorescence was measured with a pulse amplitude modulated system (Junior-PAM, Germany).Gasexchange measurements were taken from leaves, and then the same leaves were placed in darkness for 30 min.The minimum chlorophyll fluorescence (Fo) was excited by very low intensity light to keep photosystem II (PSII) reaction centers open, while a strong light pulse was used to elicit the maximum chlorophyll fluorescence (Fm), which closed all PSII reaction centers.The maximum quantum yield of PSII was calculated as:
PSII=(Fm-Fo)/Fm
The value of quantum yield efficiency of PSII of light adapted leaves (ΦPSII) was calculated according to the formula:
ΦPSII=(Fm´-F´)/Fm
Where, Fm´ and F´ are the maximum and steady state fluorescence yield of light-adapted leaves, respectively.
Photochemical fluorescence-quenching coefficients(qP) were calculated as in Schreiber et al.(1986).Nonphotochemical fluorescence quenching (NPQ) was calculated as in Bilger and Björkman (1990).Relative rate of electron transport (ETR) was calculated according to Genty et al.(1989).
2.6.Lipid peroxidation
The content of malondialdehyde (MDA) was analyzed according to the method described by Hodges et al.(1999)with slight modification, to account for the possible influence of interfering compounds in the assay for thiobarbituric acid (TBA) reactive substances.A sample of 0.2 g fresh leaf tissue was repeatedly extracted with 5 mL 10% (w:v)trichloroacetic acid (TCA).The homogenate was centrifuged at 10 000×g for 10 min and 3 mL of the supernatant was added to a test tube with an equal volume of 0.6% TBA solution.Samples were heated at 95°C for 15 min, and then cooled immediately.Absorbance was read at 440, 532, and 600 nm, respectively using a spectrophotometer (TU-1900,Beijing, China).MDA equivalents were calculated as:
MDA (μmoL L-1)=6.54×(OD532-OD600)-0.56×OD440
Proline content was determined following the ninhydrin method as described by Bates et al.(1973).A sample of 0.2 g fresh leaf tissue was extracted in 5 mL of 3% sulfosalicylic acid.After centrifugation at 5 000×g for 20 min, 2 mL of the supernatant was added to 2 mL of a mixture of glacial acetic acid and ninhydrin reagent in a 1:1 (v/v) ratio.The reaction mixture was incubated in a water bath at 100°C for 1 h and then added to 5 mL of toluene.Absorbance was read in the organic phase at 520 nm.Proline content was calculated based on a standard curve.
2.7.Extraction and assays of antioxidant enzymes
For assays of antioxidant enzymes, 0.5 g of fresh leaf was homogenized in 50 mmol L-1phosphate buffer (pH 7.8)containing 0.1 mmol L-1EDTA, 1% (w/v) polyvinylpyrrolidone(PVP) and 0.1% (v/v) Triton X-100.The homogenate was centrifuged at 10 000 r min-1for 10 min at 4°C and the supernatant was stored at -80°C or used immediately as a source of enzymes for CAT, POX, SOD, and APX assays.
Peroxidase (POD) activity was based on the determination of guaiacol oxidation (extinction coefficient 26.6 mmol L-1cm-1) at 470 nm by H2O2.The reaction mixture contained 50 mmol L-1potassium phosphate buffer (pH 7.8), 20 mmol L-1guaiacol, 12.3 mmol L-1H2O2, and 50 μL enzyme extract in a 3-mL volume.
CAT activity was determined in the homogenates by measuring the decrease in absorption at 240 nm in a reaction medium containing 50 mmol L-1potassium phosphate buffer(pH 7.2), 10 mmol L-1H2O2, and 50 μL enzyme extract.The activity was calculated using the extinction coefficient(40 mmol L-1cm-1) for H2O2.
SOD activity was determined by measuring the ability of the enzyme to inhibit the photochemical reduction of nitrobluetetrazolium (NBT).The 3 mL reaction mixture solution contained 50 mmol L-1potassium phosphate buffer(pH 7.8), 50 mmol L-1methionine, 75 μmol L-1NBT, 20 μmol L-1riboflavin, 0.1 mmol L-1EDTA, and 0.1 mL of SOD extract.The reaction was performed under 120 μmol photons m-2s-1for 15 min.Blanks and control were run similarly but without illumination and SOD extract respectively.One unit of SOD was defined as the amount of enzyme, which produced 50%inhibition of NBT reduction by monitoring at 560 nm.
APX activity was estimated by monitoring ascorbate oxidation at 290 nm (extinction coefficient 2.8 mmol L-1cm-1).The 1 mL reaction mixture consisted of 50 mmol L-1phosphate buffer (pH 7.0), 0.25 mmol L-1ascorbate,5 mmol L-1H2O2, and 10 μL enzyme extract.The reaction was started by the addition of H2O2, and the reaction rate was determined during the first 3 min.
2.8.Growth measurements
At the end of the experiment, four individuals in each water regime were harvested and divided into roots, stems, leaves,and fruit.Roots were washed thoroughly and carefully with tap water.All of the plant parts were separated and dried at 85°C for 48 h to constant mass and then weighed.Next,root biomass (RB), stem biomass (SB), leaf biomass (LB),and fruit biomass (FB) were determined.Based on these data, the following plant growth traits were calculated as:
Total biomass (TB)=RB+SB+LB+FB
Root dry mass ratio (RMR)=RB/TB
Stem dry mass ratio (SMR)=SB/TB
Leaf dry mass ratio (LMR)=LB/TB
Fruit dry mass ratio (FMR)=LB/TB
Root to shoot ratio (R/S)=RB/(SB+LB+FB)
2.9.Statistical analysis
Each experimental index was measured in at least three pots per treatment.Results are presented as means±standard error (SE).Significance at P<0.05 was assessed by ANOVA(least significant difference (LSD)) using Data Processing Station (DPS) (ver.8.0, Zhejiang University, Hangzhou,China) and Excel.
3.Results
3.1.Effect of drought stress on leaf chlorophyll content and gas-exchange
Our results showed that chlorophyll (a+b), carotenoid content, chlorophyll a/chlorophyll b, and Pndeclined rapidly in the four wolfberry species as drought stress increased(Table 1).Under control condition (75% FC), L.chinense Mill.exhibited the highest carotenoid content, chlorophyll a/chlorophyll b, and Pnamong the studied species.L.barbarum L.had the highest chlorophyll (a+b) content.Under severe drought (15% FC), L.chinense Mill.had the highest chlorophyll (a+b) content, chlorophyll a/chlorophyll b, carotenoid content, and Pnamong the four species,with values 35, 15, 41, and 64.9% lower than the control,respectively.The reduced values of chlorophyll (a+b) content were significantly lower than that in the other three species.However, L.yunnanense Kuang & A.M.Lu exhibited the lowest chlorophyll (a+b) content and Pnunder severe drought (15% FC), and these values were significantly lower than that of L.barbarum L.and L.chinense Mill.Similarly,E was also significantly reduced in the four species under severe drought (15% FC).A rapid decrease in E was recorded for L.chinense Mill., which reached the lowest level observed in the four species.
WUE in leaves of the wolfberry seedlings was higher in L.chinense Mill.than in the other three species under each of the watering regimes (Table 1).WUE was not significantly different among the four wolfberry species between control condition (75% FC) and light drought condition (55% FC).The WUE of L.chinense Mill.reached the maximum of 2.01 μmol mmol-1in moderate drought condition (35%FC), and WUE of L.barbarum L.and L.chinense Mill.var.potaninii (Pojark.) A.M.Lu reached the maxima of 1.87 and 1.75 μmol mmol-1, respectively, in light drought condition(55% FC).The WUE of L.yunnanense Kuang & A.M.Lu decreased during water stress and was the lowest in severe drought condition (15% FC), as well as being lower than the other three species.
3.2.Effect of drought stress on leaf chlorophyll fluorescence
Fluorescence characteristics are shown in Table 2.The maximum quantum yield of PSII (Fv/Fm) decreased significantly under water stress in the four wolfberry species, as well as ΦPSII, qP, and ETR.Under control condition (75% FC) and light drought condition (55% FC),there were no significant differences in Fv/Fm, ΦPSII, qP,and ETR among the four species, except for the ETR of L.chinense Mill., which was significantly lower than the other species under control condition (75% FC).Under severe drought condition (15% FC), Fv/Fm, ΦPSII, qP, and ETR were significantly lower in L.chinense Mill.var.potaninii(Pojark.) A.M.Lu and L.yunnanense Kuang & A.M.Lu than the other two species.However, L.chinense Mill.and L.yunnanense Kuang & A.M.Lu had the highest and lowest values, respectively, of the four fluorescence characteristics.Water stress treatment significantly increased NPQ in all four species (Table 2).The magnitude of this effect was much lower in L.chinense Mill.(190% the control) than in L.yunnanense Kuang & A.M.Lu (214%), L.barbarum L.(220%), and L.chinense Mill.var.potaninii (Pojark.) A.M.Lu (250%).
3.3.Effect of drought stress on leaf proline content and antioxidant enzymes
Proline and MDA contents are shown in Table 3.Drought condition significantly increased the proline and MDA contents in L.chinense Mill.and L.barbarum L.during thewater stress period.However, proline and MDA contents in L.yunnanensa Kuang & A.M.Lu increased from control condition (75% FC) to moderate drought condition (35%FC), and then decreased in severe drought condition (15%FC).Under severe drought condition (15% FC), the proline and MDA contents increased in L.chinense Mill.(210 and 163%, respectively) and in L.barbarum L.(186 and 157%,respectively), compared to the control (75% FC), and proline and MDA contents were significantly higher in these species than in the other two species.
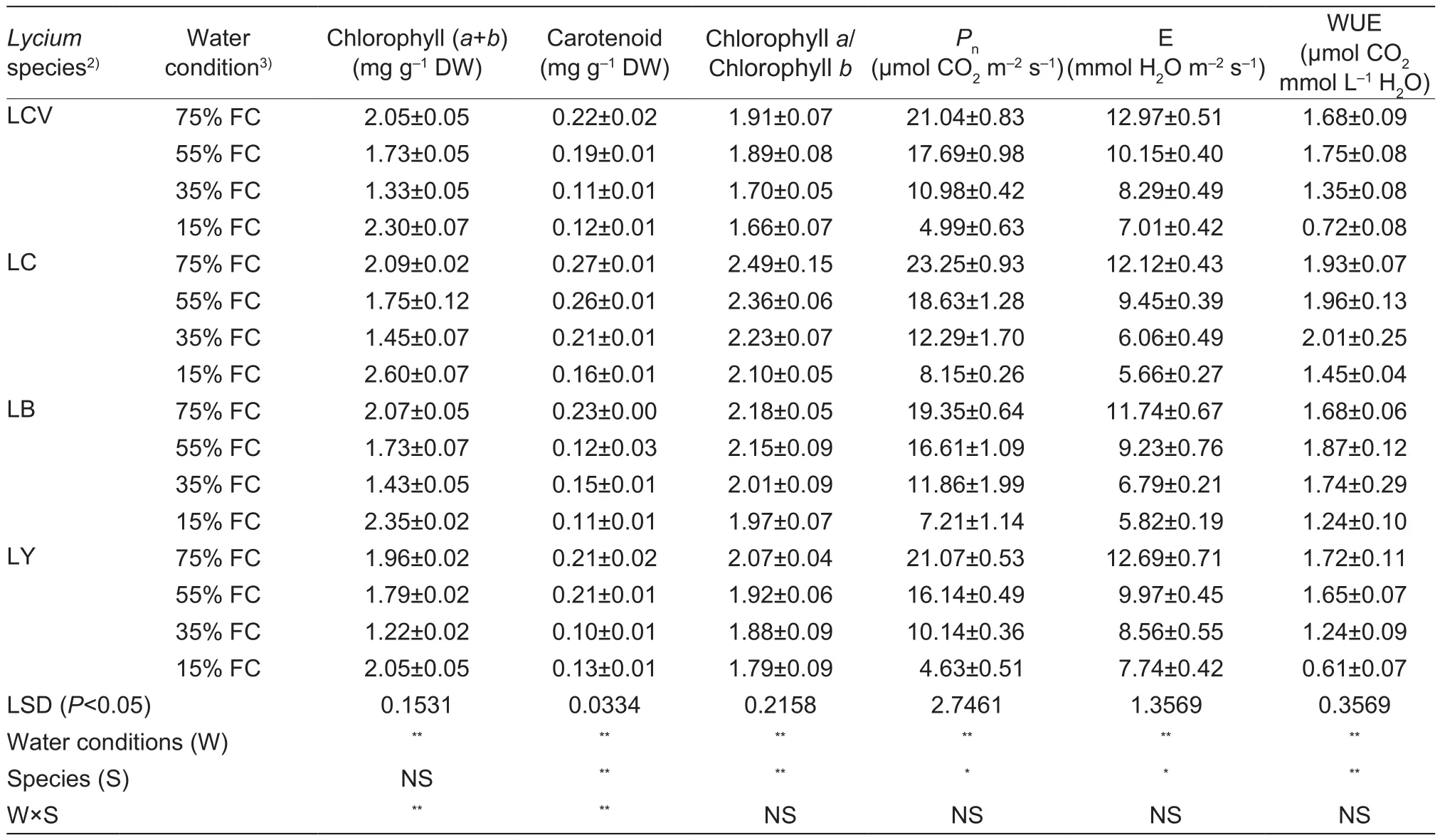
Table 1 Contents of chlorophyll (a+b) and carotenoid, chlorophyll a/chlorophyll b, Pn, E, and WUE of four wolfberry seedlings under four drought treatments1)
The POD, CAT, SOD, and APX activities increased in the initial stages of the experiment and then decreased in the four species (Table 3).There were no significant differences in the activities of the four enzymes between L.chinense Mill.and L.barbarum L.under control condition (75% FC)and light drought condition (55% FC), and enzyme activities for both species were significantly higher than those in the other two species.POD and APX activities in L.yunnanense Kuang & A.M.Lu increased under light drought condition(55% FC) and then decreased in response to moderate drought condition (35% FC); however, enzyme activities in the other three species increased with moderate drought condition (35% FC).CAT activities were similar for the four species, and had the highest value in light drought condition (55% FC) and then decreased under moderate drought condition (35% FC).SOD activities in L.chinense Mill.increased with moderate drought condition (35%FC); however, SOD activities for the other three species increased in response to light drought condition (55% FC)and then decreased under moderate drought condition(35% FC).The activity levels of the four enzymes ranked from the highest to the lowest were: L.chinense Mill.>L.barbarum L.>L.chinense Mill.var.potaninii (Pojark.) A.M.Lu>L.yunnanense Kuang & A.M.Lu.
3.4.Effect of drought stress on biomass partitioning
Dry biomass partitioning is shown in Table 4.Drought condition significantly increased biomass partitioning to roots and stems, and decreased biomass partitioning to leaves and fruits during the water stress period.In particular,L.yunnanense Kuang & A.M.Lu produced no fruit at any of the drought levels.Under the control condition (75%FC), L.yunnanense Kuang & A.M.Lu exhibited the highest biomass partitioning to roots, stems, and leaves among species.However, under light drought condition (55% FC),the increasing degree of biomass partitioning to roots was much larger in L.yunnanense Kuang & A.M.Lu (117%of the control) than in the other three species; in addition,the increasing rate of biomass partitioning to stems was much lower in L.yunnanense Kuang & A.M.Lu than the other three species.There were no significant differences in biomass partitioning to roots and stems among the four species under moderate drought condition (35% FC) or severe drought condition (15% FC).The root/shoot ratio increased in L.chinense Mill.var.potaninii (Pojark.) A.M.Lu, L.chinense Mill.and L.barbarum L., but decreased in L.yunnanense Kuang & A.M.Lu under severe drought condition (15% FC).At the end of the experiment, the root/shoot ratio in L.chinense Mill.was 2.6, 4.4 and 14.2%higher than that in L.chinense Mill.var.potaninii (Pojark.)A.M.Lu, L.barbarum L., and L.yunnanense Kuang & A.M.Lu, respectively.

Table 2 Values of Fv/Fm, ΦPSII, qP, NPQ, and ETR of four wolfberry seedlings under four drought treatments1)
4.Discussion
Chlorophyll content is a reflection of plant photosynthesis,and under conditions of water stress, chlorophyll content varies only slightly in plants that are drought tolerant (Xiao 2001; Wallin et al.2002).In this study, chlorophyll (a+b)content, carotenoid content, chlorophyll a/chlorophyll b,Pn, and E in leaves of wolfberry seedlings declined with increasing drought stress, but the depth of the decline was different for the four wolfberry species over the four drought treatments.Among the four species, the decrease in L.chinense Mill.and L.barbarum L.was lower than that in L.chinense Mill.var.potaninii (Pojark.) A.M.Lu and L.yunnanense Kuang & A.M.Lu; this was particularly true under severe drought conditions.Lycium chinense Mill.had higher values of chlorophyll (a+b), carotenoid, chlorophyll a/chlorophyll b, and Pn, and L.yunnanense Kuang & A.M.Lu had lower values of chlorophyll (a+b) and Pn.Therefore,L.chinense Mill.and L.barbarum L.had stronger drought resistance.
A decline in leaf water content can lead to decreasing chlorophyll concentration, damage to the chloroplast membrane, and a reduction in the number of accepted photons that allow plants to avoid damage from excessive free radicals (Michel and Florence 1999; Li and Qiu 2003;Anjum et al.2011).When the photosynthetic system is damaged and electron transfer is restricted, the proportion of photosynthetic pigment used in photosynthesis decreases gradually and heat dissipates more rapidly (Bruna et al.2010).In this study, Fv/Fm, ΦPSII, qP, and ETR decreased under water stress in the four wolfberry species, and the decreases in L.chinense Mill.and L.barbarum L.were lowerthan for the other two species.These results showed that the light capture and transformation abilities in L.chinense Mill.and L.barbarum L.(two species that had higher Pnunder severe drought) were stronger than that in the other two species.NPQ is one of the most important indices for monitoring excess excitation energy dissipation (Aroca et al.2003; Babita et al.2010).NPQ in seedling leaves increased after water stress treatment in all four species, and the dissipation of energy in the form of heat occurred when PSII received excess light.Under severe drought stress,NPQ in L.barbarum L.and L.chinense Mill.maintained less than the other two: they had less dissipation of heat, and a potential for improvement in the carbon assimilation ability and excitation energy efficiency in PSII.
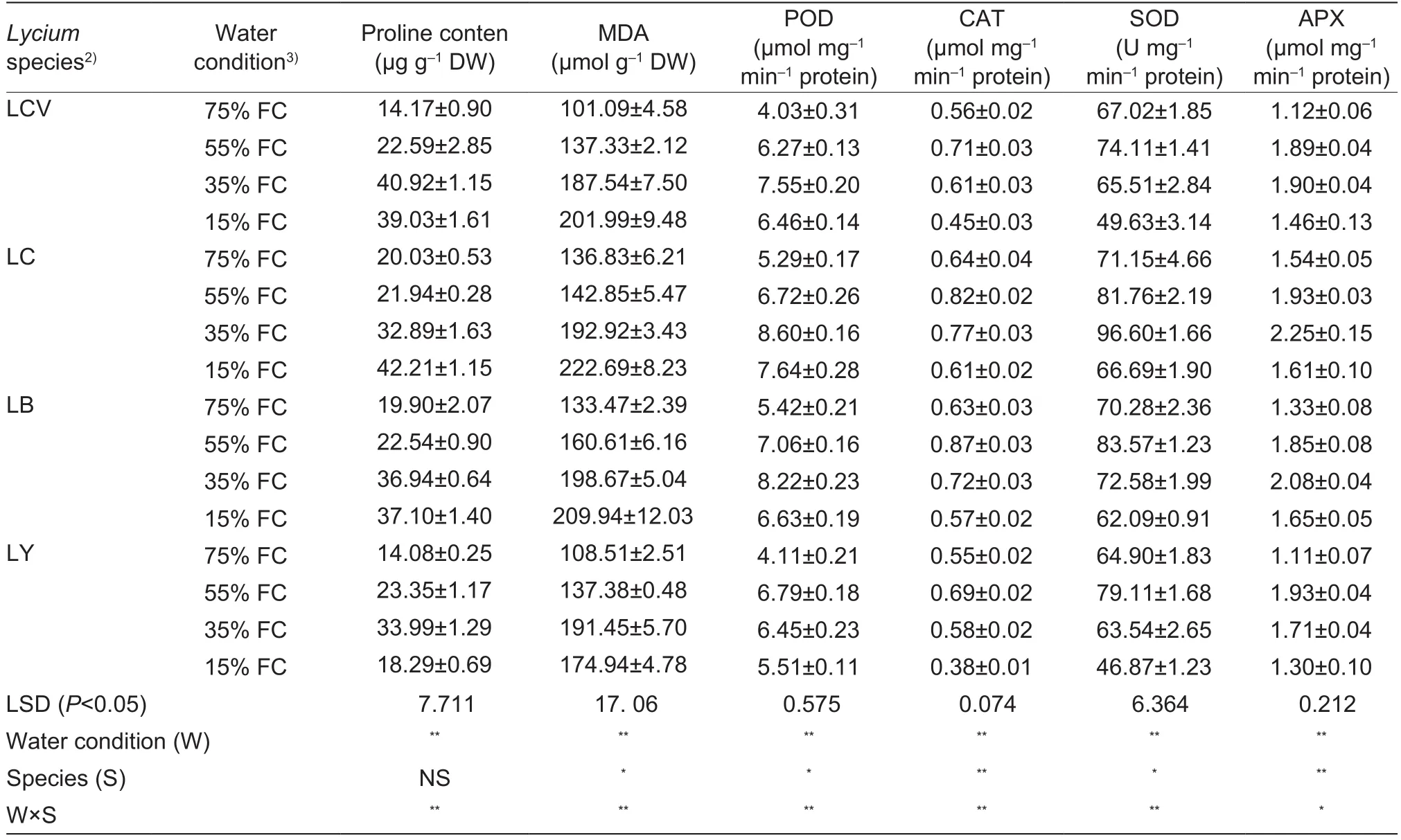
Table 3 Proline content, MDA, POD, CAT, SOD, and APX of four wolfberry seedlings under four drought treatments1)
Cell membranes of leaves were peroxidated and MDA content increased after the plants were subjected to drought stress.Simultaneously, cell membrane osmotic regulation occurred through accumulation of proline as an adaptation to the drought conditions, and proline accumulation was higher in plants with stronger drought resistance (Ashraf and Iram 2005; Chaves and Oliveira 2005).In this study,the proline and MDA contents increased in leaves of L.chinense Mill.and L.barbarum L.as water stress increased.In L.yunnanense Kuang & A.M.Lu, proline and MDA contents increased initially with elevated water stress then decreased at higher levels.Under severe drought conditions, MDA and proline contents accumulated in the four species ranked from the highest to the lowest were:L.chinense Mill.>L.barbarum L.>L.chinense Mill.var.potaninii (Pojark.) A.M.Lu>L.yunnanense Kuang & A.M.Lu.
The antioxidative defense system in plants involves the production of SOD, POD, CAT, APX, and enzymes that help the plant remove ROS and prevent membrane lipid peroxidation (Horváth et al.2007).This study found that the enzyme activities of SOD, POD, CAT, and APX increased steadily under light drought stress.In L.chinense Mill.,enzyme activities of SOD, POD, and APX continued to rise under moderate drought stress.However, in L.yunnanense Kuang & A.M.Lu, activities of all four enzymes declined under all drought conditions.Under severe drought stress,activities of the four enzymes declined in all of the species.Therefore, during drought stress, the antioxidant ability of seedling leaves gradually weakened, which was due tomembrane lipid oxidation increased.
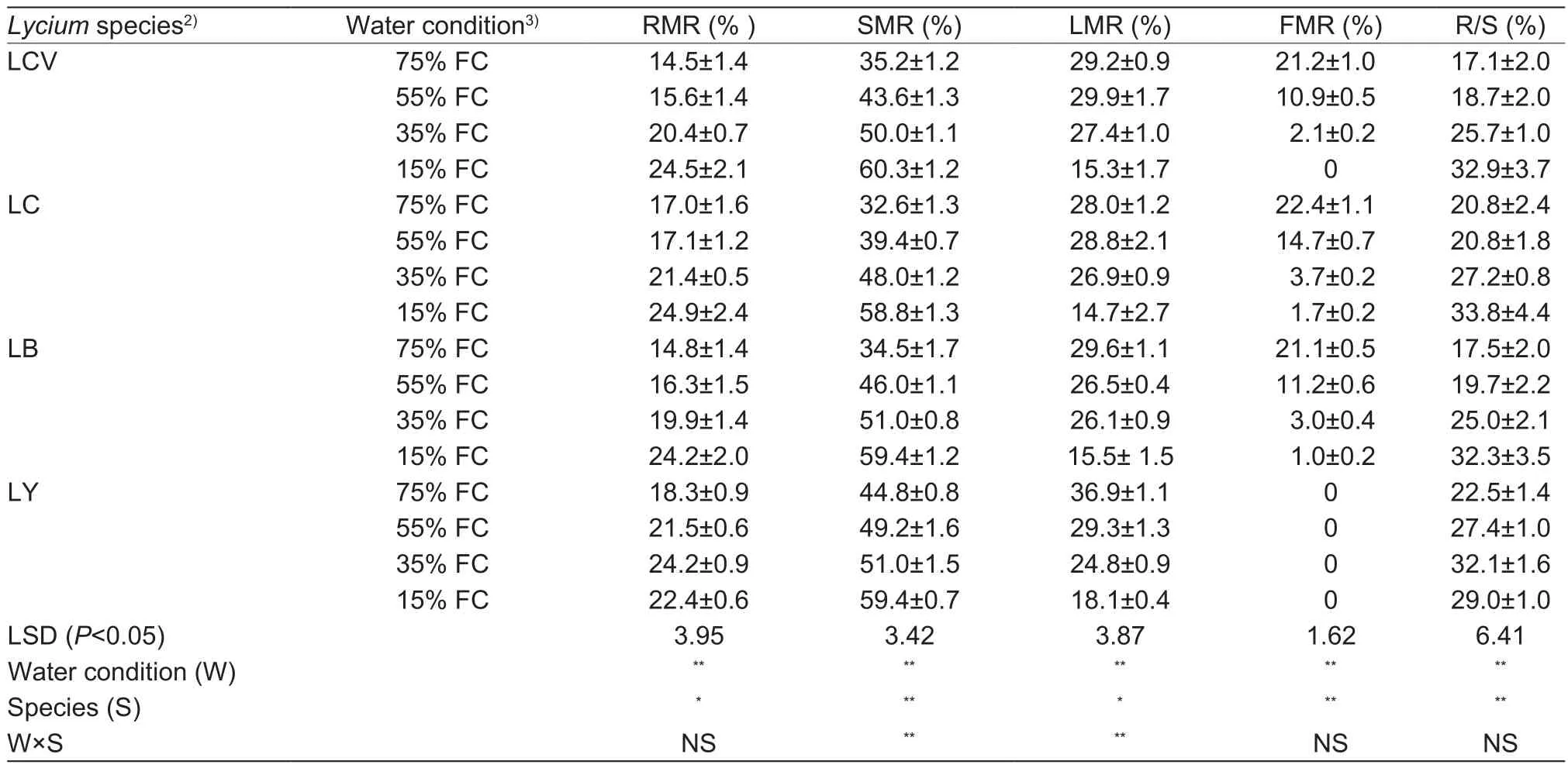
Table 4 Dry biomass partitioning of four wolfberry seedlings under four drought treatments1)
Biomass allocation in plants was typically regulated in order to adapt to a changing environment under drought stress (Zobel 2003; Anjum et al.2011).The total amount of dry mass and biomass allocation in roots and root caps changes in accordance with changes in drought stress conditions (Toorchi et al.2002).In this study, the partitioning rate of dry matter of roots, stems, leaves, and fruits in the four species showed significant differences during drought stress; in particular, distribution to stems was higher and distribution to fruit was lower after light drought stress.L.yunnanense Kuang & A.M.Lu produced no fruits at any level of drought stress.The dry mass distribution of fruits in the other three species showed a significant downward trend.The distribution rate of dry mass in fruits remained at a very low level in L.chinense Mill.and in L.barbarum L.under severe drought stress.Studies have shown that plants transmit more nutrition to stems for temporary storage in response to drought stress (Chen and Zhao 2011).The root to shoot ratio in the four species increased under light and moderate drought conditions, but it decreased in L.yunnanense Kuang & A.M.Lu under severe drought condition while the other three species were still growing.In L.chinense Mill., increased root growth was associated with decreased biomass of leaves and fruits, which was due to the adaptation to the changing drought environment.
5.Conclusion
Drought stress impacts plants in many ways, but plants can reduce or delay the damage caused by drought stress through morphological and physiological changes.This study investigated morphological, photosynthetic, and physiological aspects of Lycium species, and the tolerance of Lycium seedlings to drought stress.L.chinense Mill.and L.barbarum L.have higher capabilities to acclimate to drought conditions than the other two species, especially in severe drought conditions.L.chinense Mill.had the highest tolerance to drought stress, while L.yunnanense Kuang& A.M.Lu was found to be the most susceptible species.
Acknowledgements
This study was financially supported by the National Natural Science Foundation of China (31360191, 31660220), the Natural Science Foundation of Ningxia Hui Autonomous Region, China (NZ16121), and the Self-option and Foundation of Ningxia Academy of Agriculture and Forestry Sciences, China (YES-16-0402, NKYZ-16-0402).
Anjum S A, Xie X, Wang L C, Saleem M F, Man C, Lei W.2011.Morphological, physiological and biochemical responses of plants to drought stress.African Journal of Agricultural Research, 6, 2026-2032.
Arnon D I.1949.Copper enzymes in isolated chloroplasts:Polyphenol oxidase in Beta vulgaris.Plant Physiology,24, 1-15.
Aroca R, Irigoyen J J, Sánchez-díaz M.2003.Drought enhances maize chilling tolerance.II.Photosynthetic traits and protective mechanisms against oxidative stress.Physiologia Plantarum, 117, 540-549.
Ashraf M, Iram A.2005.Drought stress induced changes in some organic substances in nodules and other plant parts of two potential legumes differing in salt tolerance.Flora,200, 535-546.
Ávila C, Guardiola J L, Nebauer S G.2012.Response of the photosynthetic apparatus to a flowering-inductive period by water stress in citrus.Trees, 26, 833-840.
Babita M, Maheswari M, Rao L M, Shanker A K, Rao D G.2010.Osmotic adjustment, drought tolerance and yield in castor (Ricinus communis L.) hybrids.Environmental and Experimental Botany, 69, 243-249.
Bates L S, Waldren R P, Teare I D.1973.Rapid determination of free proline for water-stress studies.Plant & Soil, 39,205-207.
Bhatt D, Negi M, Sharma P, Saxena S C, Dobriyal A K, Arora S.2011.Responses to drought induced oxidative stress in five finger millet varieties differing in their geographical distribution.Physiology & Molecular Biology of Plants, 17,347-353.
Bilger W, Björkman O.1990.Role of the xanthophyll cycle in photo-protection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis.Photosynthesis Research, 25, 173-185.
Bruna D S, Bruno M R, Laurício E, Mauro G S.2010.Ecophysiology parameters of four Brazilian Atlantic Forest species under shade and drought stress.Acta Physiologiae Plant, 32, 729-737.
Chang R C, So K F.2008.Use of anti-aging herbal medicine,Lycium barbarum, against aging-associated diseases.What do we know so far? Cellular & Molecular Neurobiology, 28,643-652.
Chaves M M.1991.Effects of water deficits on carbon assimilation.Journal of Experimental Botany, 42, 1-16.
Chaves M M, Oliveira M M.2005.Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture.Journal of Experimental Botany, 55, 2365-2384.
Chen J Y, Zhang Z T, Li Y N.2004.Effects of NaCl stress on betaine, chloroplast pigment of leaves chlorophyll fluorescence and of Lycium barbarum L.Agricultural Research in the Arid Areas, 22, 109-114.(in Chinese)
Chen M T, Zhao Z.2011.Effects of drought on root characteristics and mass allocation in each part of seedlings of four tree species.Journal of Beijing Forestry University,33, 16-22.(in Chinese)
CCP (Committee of Chinese Pharmacopoeia).2010.Chinese Pharmacopoeia.Committee of Chinese Pharmacopoeia,Beijing.(in Chinese)
Deng B, Du W, Liu C, Sun W, Tian S, Dong H.2012.Antioxidant response to drought, cold and nutrient stress in two ploidy levels of tobacco plants: Low resource requirement confers polytolerance in polyploids? Plant Growth Regulation, 66,37-47.
Diego N D, Pérezalfocea F, Cantero E, Lacuesta M, Moncaleán P.2012.Physiological response to drought in radiata pine:Phytohormone implication at leaf level.Tree Physiology,32, 435-449.
Gao D, Gao Q, Xu H Y, Ma F, Zhao C M, Liu J Q.2009.Physiological responses to gradual drought stress in the diploid hybrid Pinus densata and its two parental species.Trees, 23, 717-728.
Genty B, Briantais J M, Baker N R.1989.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.Biochimica et Biophysica Acta (BBA)-General Subjects,990, 87-92.
Hodges D M, DeLong J M, Forney C F, Prange R K.1999.Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds.Planta, 207,604-611.
Horváth E, Pál M, Szalai G, Páldi E, Janda T.2007.Exogenous 4-hydroxybenzoic acid and salicylic acid modulate the effect of short term drought and freezing stress on wheat plants.Biologia Plantarum, 51, 480-487.
Inskeep W P, Bloom P R.1985.Extinction coefficients of chlorophyll a and b in N,N-dimethylformamide and 80%acetone.Plant Physiology, 77, 483-485.
Li H C, Qiu Z J.2003.A review of studies of drought resistance in tree species and drought resistant forestation technology.World Forestry Research, 16, 17-22.
Michel H, Florence T.1999.Loss of chlorophyll with limited reduction of photosynthesis as an adaptive response of Syrian barley landraces to high-light and heat stress.Australian Journal of Plant Physiology, 26, 569-578.
Reddy A R, Chaitanya K V, Vivekanandan M.2004.Droughtinduced response of photosynthesis and antioxidant metabolism in higher plants.Journal of Plant Physiology,161, 1189-1202.
Ribas-Carbo M, Taylor N L, Giles L, Busquets S, Finnegan P M, Day D A, Lambers H, Medrano H, Berry J A, Flexas J.2005.Effects of water stress on respiration in soybean leaves.Plant Physiology, 139, 466-473.
Schreiber U, Schliwa U, Bilger W.1986.Continuous recording of pho-tochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer.Photosynthesis Research, 10, 51-62.
Toorchi M, Shashidhar H E, Hittalmani S, Gireesha T M.2002.Rice root morphology under contrasting moisture regimes and contribution of molecular marker heterozygosity.Euphytica, 126, 251-257.
Wallin G, Karlsson P E, Selldn G, Ottosson S, Medin E L, Pleijel H, Skarby L.2002.Impact of four years exposure to different levels of ozone, phosphorus and drought on chlorophyll,mineral nutrients, and stem volume of Noway spruce, Picea abies.Physiologia Plantarum, 11, 192-206.
Wang Y, Ma F, Li M, Liang D, Zou J.2011.Physiological responses of kiwifruit plants to exogenous ABA under drought conditions.Plant Growth Regulation, 64, 63-74.
Xiao Y A.2001.The physiology responses and adjective adaptability of water stress on Cleme spinosa L.seedlings.Journal of Wuhan Botanical Research, 19, 524-528.(in Chinese)
Xie C, Xu L Z, Li X M, Zhao B H, Yang S L.2001.Studies on chemical constituents in fruit of Lycium barbarum L.China Journal of Chinese Materia Medica, 26, 323-324.(in Chinese)
Xu Y H, Xu Y, An W T.2000.The progress in studies on antitumor pharmacodynamics of Lycium barbarum Medicine and Materia Medica Research, 11, 946-947.(in Chinese)
Zobel R W.2003.Sensitivity analysis of computer based diameter measurement from digital images.Crop Science,43, 583-591.
杂志排行

Journal of Integrative Agriculture的其它文章
- Characteristic analysis of tetra-resistant genetically modified rice
- A wheat gene TaSAP17-D encoding an AN1/AN1 zinc finger protein improves salt stress tolerance in transgenic Arabidopsis
- Characterization of GhSERK2 and its expression associated with somatic embryogenesis and hormones level in Upland cotton
- GmNAC15 overexpression in hairy roots enhances salt tolerance in soybean
- Molecular cloning and functional characterization of a soybean GmGMP1 gene reveals its involvement in ascorbic acid biosynthesis and multiple abiotic stress tolerance in transgenic plants
- Responses of the antioxidant system to fluroxypyr in foxtail millet(Setaria italica L.) at the seedling stage