3个牡丹品种花期进程中花瓣的生理生化特征
2018-03-05吴桂容王上伟苏德生卢云天
吴桂容,王上伟,苏德生,张 萍,卢云天
(1.贺州学院化学与生物工程学院,广西贺州 542800;2.福建省政和县林业局,福建政和 353600;3.福建省政和县镇前镇油用牡丹专业合作社,福建政和 353600;4.福建富之卿牡丹科技发展有限公司,福建福州 350000)
牡丹(Paeoniasuffruticosa)为芍药科芍药属多年生落叶小灌木,是我国特有的传统名花,具有极高的观赏、药用和经济价值。牡丹的花期及其持续时间是影响其观赏价值和市场开发的关键因素,研究牡丹花期进程中生理、生化特征的改变,延长开花质量和花期持续时间具有重要的意义[1-2]。目前,已有一些研究对开花进程中牡丹的生理生化特征进行了研究,如郭丽丽等对盆栽与地栽牡丹花芽的生理生化特性动态进行了研究,指出不同栽培方式的牡丹花芽营养和抗氧化酶系统等生理生化参数有较大差别,但在花期进程中有着相似的变化趋势[3];张圣旺等对牡丹花衰老过程中的生理生化特征进行了研究,指出细胞质膜透性、超氧化毒害是影响牡丹花衰老的决定性因素[4],但这些研究系统性尚较差。因此,本研究对不同牡丹品种乌龙捧盛、景玉、香玉花期进程中的可溶性糖、可溶性蛋白、脯氨酸、超氧化物歧化酶(SOD)、过氧化物酶(POD)、丙二醛(MDA)等营养、渗透调节物质及抗氧化酶系统进行比较研究,从而为牡丹花期中衰老进程提供一定的参考,并筛选出适合引种栽培推广的牡丹品种。
1 材料与方法
1.1 试验地概况
试验地位于福建省南平市政和县镇前镇,地处武夷山鹫峰山脉北瑞,位于闽东北结合部,118°56′~119°16′E,27°08′~27°09′N之间。平均海拔936 m,年降水量1 726 mm,年均气温13.9 ℃。夏季干爽凉人,昼夜温差大。冬季寒冷,普遍低温。
1.1 材料
供试材料为牡丹品种乌龙捧盛、香玉、景玉,为福建省南平市政和基地初步筛选的适生品种。在2016年3月牡丹现蕾后,选择花蕾生长状况一致的牡丹植株标记为研究对象。按照郭倩等的标准[5],从标记的牡丹植株上采摘小风铃期(S)、大风铃期(L)、露色期(Ⅰ)、绽口期(Ⅱ)、初放期(Ⅲ)、半开期(Ⅳ)、盛开期(Ⅴ)和始衰期(Ⅵ期)等不同发育状态的花朵,各花期每个品种各选择5朵花,采集的花朵样品于实验室内进行生理生化检测。
1.2 测试方法
可溶性糖含量测定采用蒽酮比色法;可溶性蛋白含量测定采用考马斯亮蓝G-250比色法;脯氨酸含量测定采用酸性茚三酮法;MDA含量测定采用硫代巴比妥酸法[6]。SOD、POD活性采用南京建成生物工程研究所生产的SOD、POD试剂盒(羟胺法、比色法)进行测定。
1.3 统计学方法
用Excel 2013进行数据整理与分析。
2 结果和分析
2.1 花期进程中花瓣的可溶性糖含量变化
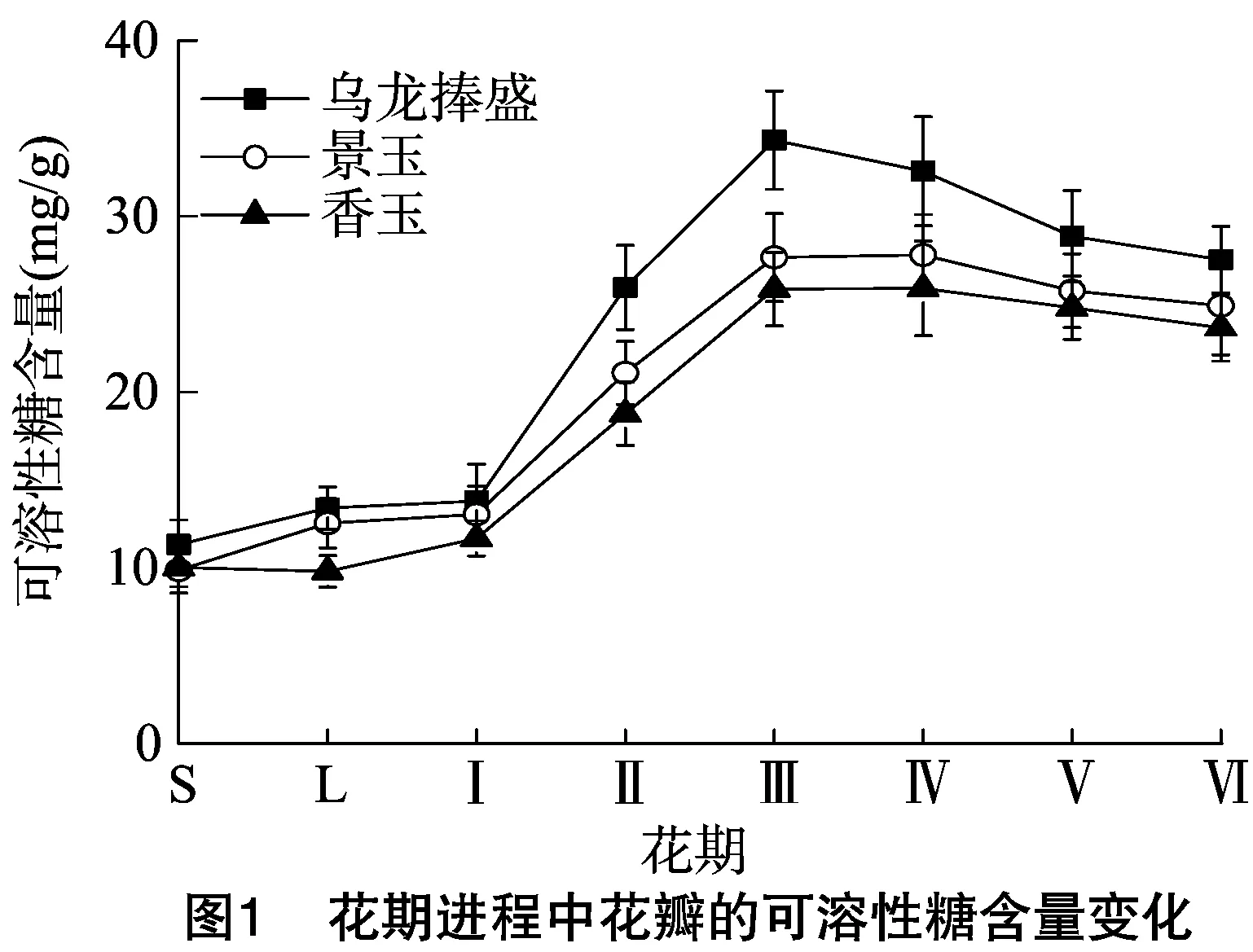
花期进程中花瓣的可溶性糖含量变化如图1。乌龙捧盛、香玉、景玉花瓣中的可溶性糖含量均呈先上升后下降的趋势,均从露色期(Ⅰ)开始快速上升并在初放期(Ⅲ)达最高值,随后缓慢下降。乌龙捧盛不同花期花瓣的可溶性糖含量最高,其次为景玉,再次为香玉。可溶性糖是花瓣可以直接利用的养分形式,是碳水化合物的暂存形式[7]。在花朵绽放之前,花瓣储存可溶性糖供花朵开放,而在花朵绽放过程中则大量消耗能量,造成花瓣的可溶性糖含量下降,这与崔洋等对湖南牡丹的研究结果[8]是一致的。
2.2 花期进程中花瓣的可溶性蛋白含量变化
花期进程中花瓣的可溶性蛋白含量变化如图2所示。乌龙捧盛、香玉、景玉花瓣中的可溶性糖含量均呈总体下降的趋势,其中在中期快速下降而后则放缓。乌龙捧盛、景玉均在绽口期(Ⅱ)到初放期(Ⅲ)快速下降,但乌龙捧盛的下降幅度要小于景玉;而香玉则从露色期(Ⅰ)到绽口期(Ⅱ)即开始快速下降。不同花期乌龙捧盛花瓣的可溶性蛋白含量最高,其次为景玉,再次为香玉。可溶性蛋白是花朵开放中主要的营养物质基础,其含量可以作为开花进程中衰老的指标之一[9]。本研究中3个牡丹品种花瓣中可溶性蛋白含量随着花期的进展而下降,但其快速下降的起始时期和幅度有着较大的差异,说明可溶性蛋白含量对花期维持时间有着重要的影响。

2.3 花期进程中花瓣的脯氨酸含量变化
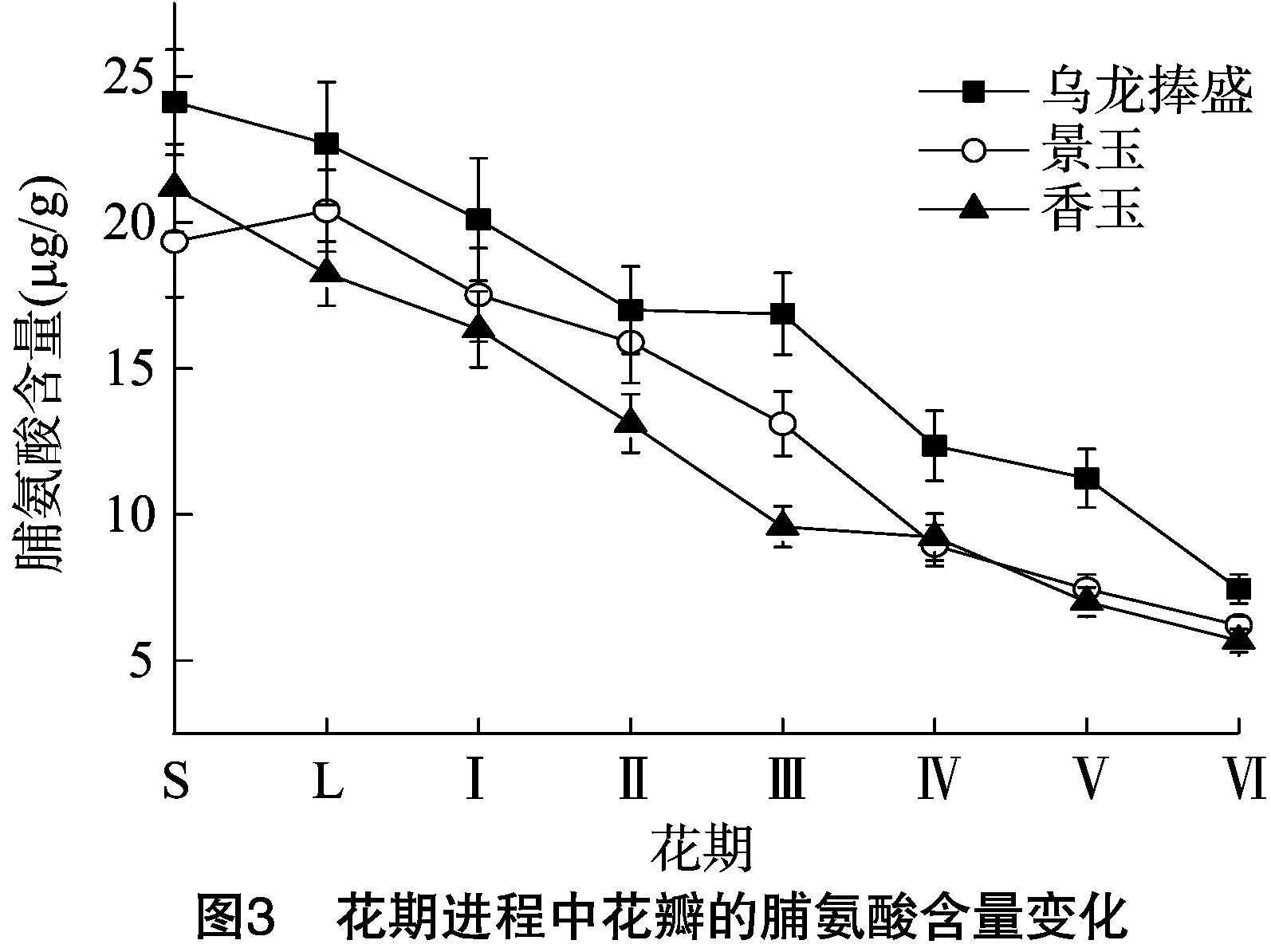
脯氨酸是细胞内的渗透调节物质,是植物面对逆境时的重要调节物质,能一定程度上说明植物的抗逆性[10],花期进程中花瓣的脯氨酸含量变化如图3所示。乌龙捧盛、香玉、景玉花瓣中的脯氨酸含量均呈总体下降的趋势,且各期的下降速度基本一致,说明随着花期的进展,花瓣不断衰老,其抗逆性下降。不同花期乌龙捧盛花瓣的脯氨酸含量最高,其次为景玉,再次为香玉,品种间花瓣中脯氨酸含量差异说明其抗逆性存在一定差异。
2.4 花期进程中花瓣的SOD活性变化
SOD是生物体内最重要的抗氧化酶,是抵御活性氧毒害的第一道防线[11],花期进程中花瓣的SOD活性变化见图4。乌龙捧盛、香玉、景玉花瓣中的SOD活性均呈总体下降的趋势,但开始下降的初始时期有所不同。乌龙捧盛从半开期(Ⅳ)开始下降,景玉从初放期(Ⅲ)开始下降,而香玉则从绽口期(Ⅱ)开始下降。不同花期乌龙捧盛花瓣的SOD活性最高,而景玉和香玉的差异较小。本研究中3个牡丹品种花瓣中SOD活性随着花期的进展而下降,说明随着花期的进展,花瓣不断受到活性氧毒害。不同牡丹品种花瓣中SOD活性开始下降的起始花期不同,则说明造成花瓣开始衰老的时间不同,清除氧自由基的能力存在差异,进而影响花期持续时间。
2.5 花期进程中花瓣的POD活性变化
花期进程中花瓣的POD活性变化见图5。乌龙捧盛、香玉、 景玉花瓣中的POD活性也均呈总体下降的趋势,其中景玉、香玉从半开期(Ⅳ)开始快速下降,而乌龙捧盛不同花期下降的幅度较为一致。不同花期乌龙捧盛花瓣的POD活性最高,而景玉和香玉的活性相对较低。POD是生物体内另一种重要的抗氧化酶,通常与SOD协同发挥抗氧化作用[12-15]。本研究中3个牡丹品种花瓣中POD活性的变化趋势与SOD基本一致,花瓣中POD活性也随着花期的进程其活性氧毒害不断下降。


2.6 花期进程中花瓣的MDA含量变化
MDA是一种过氧化合物,能强烈地与细胞内各种成分发生反应,造成细胞内酶和膜系统的损害[16],花期进程中花瓣的MDA含量变化如图6所示。乌龙捧盛、香玉、景玉花瓣中的MDA含量均呈总体上升的趋势,且随着花期进展上升幅度基本一致,说明花瓣中MDA含量上升是其衰老的重要标志。不同花期景玉花瓣的MDA含量最高,景玉次之,乌龙捧盛最低。

3 结论与讨论
牡丹花期初始阶段花瓣代谢旺盛,能不断从营养器官吸收和储存营养物质,因此可溶性糖、可溶性蛋白等营养物质含量上升。而随着花期进展,牡丹花开放需要使用大量的营养,造成花瓣中的营养物质含量降低。与此同时,随着花期的进展,牡丹花瓣中的抗逆物质脯氨酸的含量开始不断下降,抗氧化酶SOD、POD活性不断下降,且过氧化合物MDA的累积不断增加,花瓣不断衰老。不同牡丹品种花瓣中营养物质、抗逆物质、抗氧化酶的含量越高,过氧化合物的累积量越少则能延缓花瓣的衰老,维持更长的花期。在牡丹的引种中通过以上指标的对比,能够挑选到抗逆性强、花色优美、花期持续时间长的牡丹品种。本研究中供试的3种牡丹品种中乌龙捧盛最适于引种,景玉次之,最差则为香玉。
[1]陈富慧,索志立,赵孝庆,等.中国牡丹品种的花期[J].东北林业大学学报,2005,33(6):55-61.
[2]刘 娜,秦安臣,陈 雪,等.牡丹花期对生长调节剂调控响应的研究[J].河北农业大学学报,2014,37(2):31-39.
[3]郭丽丽,侯小改,郭 琪,等.盆栽与地栽牡丹花芽的生理生化特性的动态变化[J].江苏农业科学,2015,43(10):220-222.
[4]张圣旺,郑荣生,孟 丽,等.牡丹花衰老过程中的生理生化变化[J].山东农业大学学报(自然科学版),2002,33(2):166-169.
[5]郭 倩,施 江,史国安.花期进程中2个牡丹品种花瓣的生理生化特性[J].江苏农业科学,2014,42(11):196-198.
[6]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[7]Boriboonkaset T,Theerawitaya C,Yamada N,et al.Regulation of some carbohydrate metabolism-related genes,starch and soluble sugar contents,photosynthetic activities and yield attributes of two contrasting rice genotypes subjected to salt stress[J].Protoplasma,2013,250(5):1157-1167.
[8]崔 洋,张 凌,李永华,等.不同生育时期的牡丹叶片和花瓣光合相关生理指标变化分析[J].河南农业大学学报,2014,48(1):29-33.
[9]乔永旭.蝴蝶兰花衰老过程中主要生理指标的变化[J].西南农业学报,2014,27(3):1045-1048.
[10]Kishor P B K,Kumari P H,Sunita M S,et al.Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny[J].Frontiers in Plant Science,2015,6:544.
[11]吴永波,叶 波.高温干旱复合胁迫对构树幼苗抗氧化酶活性和活性氧代谢的影响[J].生态学报,2016,36(2):403-410.
[12]王宏民,李亚芳,张仙红,等.几种杀虫剂胁迫对黄瓜幼苗抗氧化酶活性和相关生理指标的影响[J].中国生态农业学报,2015,23(9):1185-1190.
[13]穆蓁蓁,克热木·伊力,王一静.高温干旱对库尔勒香梨叶片生理指标的影响[J].江苏农业科学,2016,44(2):209-212.
[14]黄雪妮,屈 凡,马名立,等.镉胁迫对2个宁夏主栽水稻品种幼苗期抗氧化同工酶亚基及其活性的影响[J].江苏农业科学,2016,44(7):107-112.
[15]韩金龙,李 慧,蔺 经,等.钙对盐胁迫下杜梨叶片抗氧化系统的影响[J].江苏农业科学,2016,44(6):245-248.
[16]唐秀梅,陈 元,钟瑞春,等.广西花生品种(系)苗期生长耐旱性及相关生理特性研究[J].广东农业科学,2014,41(14):9-13.
