消化对益生菌干酪抗氧化活性的影响
2018-02-28曲秀伟王海霞李晓东张秀秀
曲秀伟,刘 璐,王海霞,李晓东*,张秀秀,陈 萍,沈 琪
契达干酪(Cheddar cheese)是一种酸性半硬质成熟干酪[1],是世界干酪生产的主要品种之一,也是世界上购买与消费最多的一种干酪。契达干酪在成熟过程中伴随着一系列复杂的生化反应,如糖酵解、脂肪和蛋白质水解等[2]。其中,蛋白质水解是干酪成熟中最重要和复杂的反应之一,不仅对干酪的质地和风味产生重要影响[3-4],而且在此过程中,酪蛋白被来自发酵剂微生物菌群所分泌的蛋白酶和肽酶水解成各种生物活性肽[5],所以干酪具有较高的营养价值。
近年来,关于功能型益生菌乳制品的开发受到各国学者的关注。益生菌能够调节肠道菌群、合成维生素和促进营养物质吸收,同时具有提高机体免疫、降胆固醇、抗氧化等生理功能[6-7]。研究表明,干酪的稠密基质和相对较高的脂肪含量可以对高酸性环境中的益生菌起到缓冲和保护作用,是传递益生菌的良好载体[8]。有研究表明,将益生菌应用到干酪等乳制品中,既对产品的抗氧化活性产生影响,又能够产生具有抗氧化活性的多肽[1,9]。Mushtaq等[10]研究表明益生菌的添加能提高贮藏期卡拉里干酪的抗氧化活性,且不同益生菌对干酪抗氧化活性的提高程度不同。有学者从添加干酪乳杆菌的契达干酪中提取了5 种分子质量低于3 kDa的短肽,其中氨基酸序列为VKEAMAPK的短肽与天然抗氧化剂丁基羟基茴香醚、特丁基对苯二酚的抗氧化活性接近[11]。然而,食品中的活性成分只有顺利经过胃肠消化并被机体吸收利用,才能发挥其生物活性;因此,一个必要的前提是在体外建立胃肠道消化模型对其进行体外验证。目前,关于益生菌干酪的抗氧化活性在胃肠消化时的变化研究较少,主要集中在活菌数的变化,研究表明干酪中的益生菌经消化后有良好的存活,能够维持在106CFU/g左右[12]。有学者对干酪模拟胃肠道消化后血管紧张素转换酶(angiotensin converting enzyme,ACE)抑制活性的变化进行研究,结果显示模拟胃肠道消化后干酪ACE抑制活性有所增加,可能是由于大分子质量的蛋白水解产生一些具有ACE抑制活性的短肽[13]。
因此,本实验研究益生菌契达干酪经模拟胃肠道消化后抗氧化活性的变化,探究模拟胃肠道消化对干酪抗氧化活性的影响并明确影响因素,以期为评价干酪成熟过程中形成的抗氧化肽及存活的菌种对人体的抗氧化作用提供理论参考,为益生菌在干酪中的应用开发提供理论支持。
1 材料与方法
1.1 菌株、材料与试剂
双歧杆菌(Bifidobacterium bifidum)07-300B、嗜酸乳杆菌(Lactobacillus acidophilus)1.0357来自东北农业大学乳品重点实验室的菌种库;商业发酵剂(L. lactis subsp. cremoris和L. lactis subsp. lactis混合菌种)、凝乳酶Stamix 1150 北京科汉森公司。
生牛乳(新鲜无抗乳)取自哈尔滨市香坊农场完达山奶源基地;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)、2,2’-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐(2,2’-azino-bis(3-ethylbenzthiozoline-6)-sulphonic acid,ABTS) 美国Sigma公司;氯化铁、铁氰化钾 天津市科密欧化学试剂有限公司;三氯乙酸(trichloroacetic acid,TCA) 上海凌峰化学试剂有限公司;胰蛋白酶(250 U/L)、胃蛋白酶(3 000 U/L)诺维信(中国)生物技术有限公司;谷胱甘肽(还原型) 美国Amresco公司。其他试剂均为分析纯国药集团化学试剂有限公司。
1.2 仪器与设备
BSA124S电子分析天平、pH计 北京赛多利斯科学仪器有限公司;数显恒温水浴箱 常州丹瑞实验仪器设备有限公司;3K15离心机 德国Sigma公司;FUT-1800紫外-可见分光光度计 北京普析通用仪器有限责任公司;Mini-PROTEAN Tetra Cell电泳仪 美国伯乐公司。
1.3 方法
1.3.1 实验设计
以空白、单一及复合添加双歧杆菌及嗜酸乳杆菌的契达干酪为研究对象,以DPPH自由基清除率、还原力及ABTS+·清除率为评价指标,研究干酪在模拟胃肠道消化后抗氧化活性的变化;同时研究模拟胃肠道消化后益生菌活菌数、多肽含量的变化以及不同分子质量多肽的抗氧化活性,探究益生菌活菌数及多肽的变化对干酪抗氧化活性的影响,明确干酪在模拟胃肠道消化过程中抗氧化活性变化的影响因素。
1.3.2 双歧杆菌、嗜酸乳杆菌辅助发酵剂的制备
双歧杆菌、嗜酸乳杆菌辅助发酵剂的制备参照Chaves等[12]的方法略作改动。双歧杆菌07-300B、嗜酸乳杆菌1.0357分别以体积分数2%接种到60 mL质量分数12%灭菌复原脱脂乳中,厌氧培养24 h,培养两代至活菌数为1.0×109CFU/mL左右备用,以体积分数1.2%添加于干酪中。
1.3.3 益生菌契达干酪的制作
益生菌契达干酪的制作及分组如下:空白组:只添加质量分数0.01%商业发酵剂;实验组:1)双歧杆菌组:添加质量分数0.01%商业发酵剂和体积分数1.2%双歧杆菌;2)嗜酸乳杆菌组:添加质量分数0.01%商业发酵剂和体积分数1.2%嗜酸乳杆菌;3)双歧杆菌+嗜酸乳杆菌组:添加质量分数0.01%商业发酵剂、体积分数0.6%双歧杆菌和体积分数0.6%嗜酸乳杆菌。干酪的制作参照Ayala-Bribiesca等[14]的方法。
1.3.4 干酪模拟胃肠道实验
模拟胃液、肠液的制备参照Mozzi等[15]的方法略作改动。人工胃液的组成包括氯化钾(1.12 g/L)、氯化钠(2.0 g/L)、氯化钙(0.11 g/L)和磷酸二氢钾(0.4 g/L),需经过121 ℃杀菌15 min。用之前,添加胃蛋白酶(0.2 g/L),用1 mol/L HCl溶液调pH值到2.0。模拟肠液则将胰蛋白酶添加到配制好的胃液中,达到最终浓度为1.95 g/L,用1 mol/L NaHCO3溶液调pH值到7.0。
将成熟24 周后的各组干酪研碎,取10 g干酪放入装有100 mL模拟胃液的锥形瓶中,置于37 ℃恒温水浴振荡器中模拟胃液消化2 h;胃液消化后,向消化液中加入0.195 g胰蛋白酶,并用1 mol/L的NaHCO3溶液调整pH值为7.0,置于37 ℃恒温水浴振荡器中模拟肠液消化4 h[13]。
1.3.5 胃液、肠液消化后及消化前样品的制备
将各组干酪经胃液、肠液消化后的样液,在4 ℃、3 000×g离心30 min,取上层溶液通过Whatman No. 2滤纸过滤,所得提取液即为胃液、肠液消化后样品。消化前的样品制备参照Bottesini等[16]的方法略作修改,将10 g干酪加入到100 mL蒸馏水中,混合匀浆1.5 min,然后在4 ℃,3 000×g下离心30 min,取上层溶液通过Whatman No. 2滤纸过滤,所得水溶性提取液即为消化前样品[16]。
1.3.6 干酪经模拟胃肠道消化抗氧化活性的测定
DPPH自由基清除率的测定参照Shen Yingbin等[17]的方法;还原能力的测定参照Duh等[18]的方法,以A700nm表示;ABTS+·清除率的测定参照Abadía-García等[9]的方法。
1.3.7 干酪经模拟胃肠道消化益生菌活菌数测定
益生菌的计数参照de Oliveira等[19]的方法,采用MRS稀释平板法计数。
1.3.8 干酪经模拟胃肠道消化多肽变化研究
1.3.8.1 干酪SDS-PAGE
十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDSPAGE)参照金江玉等[20]的方法,选用低分子质量蛋白Marker(1.06~26.60 kDa)。
1.3.8.2 干酪经模拟胃肠道消化多肽质量浓度变化测定
标准曲线的绘制:配制10.0 mg/mL的谷胱甘肽标准溶液,依次吸取上述溶液0、1、2、3、4、5 mL置于6 支试管中,分别加入4 mL双缩脲试剂,用蒸馏水定容至10 mL,于60 ℃水浴下显色5 min,水浴期间不停地振荡,然后冷却至室温,于540 nm波长处测定吸光度(以第1管作空白对照)。以多肽质量浓度(mg/mL)为横坐标,吸光度为纵坐标,绘制标准曲线。
多肽含量的测定参照徐娟等[21]的方法略作修改。取5.0 mL样品溶液,加入5.0 mL 10 g/100 mL的TCA水溶液,于漩涡混合仪上混合均匀,静置20 min,然后4 000×g离心10 min。然后取2.0 mL上述溶液置一试管中,加入双缩脲试剂4.0 mL,于漩涡混合仪上混合均匀,60 ℃水浴显色5 min,2 000×g离心10 min。取上清液于540 nm波长处测定吸光度,对照标准曲线求得样品溶液中的多肽质量浓度(mg/mL)。
1.3.8.3 超滤分离干酪多肽及抗氧化活性测定
选用截留分子质量(10、3 kDa)的超滤膜对干酪多肽进行分离,先后得到不同截留分子质量的超滤液,得到3 个不同的组分:组分Ⅰ:(>10 kDa)、组分Ⅱ:(3~10 kDa)、组分Ⅲ:(<3 kDa)收集备用。不同分子质量多肽DPPH自由基清除率、还原能力、ABTS+·清除率的测定方法同1.3.6节所述。
1.4 数据处理
2 结果与分析
2.1 干酪经模拟胃肠道消化抗氧化活性的变化2.1.1 DPPH自由基清除率变化
DPPH自由基能够稳定存在,且易与具有氢键供体的化合物发生电子转移反应,已成为检测物质抗氧化活性的一种常用方法[22]。本实验测定了干酪消化后DPPH自由基清除率的变化,结果如图1所示。
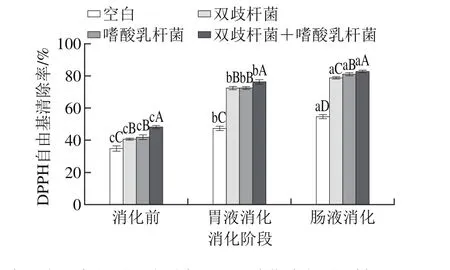
图1 干酪经模拟胃肠道消化DPPH自由基清除率变化Fig. 1 Change in DPPH radical scavenging activity of cheese after simulated gastrointestinal digestion
由图1可知,空白组及各实验组益生菌干酪DPPH自由基清除率经模拟胃肠道消化后均有显著提高(P<0.05)。空白组干酪DPPH自由基清除率从(34.79±3.11)%增加到(54.69±2.89)%,增幅为57.20%;复合添加双歧杆菌和嗜酸乳杆菌的干酪DPPH自由基清除率从(48.28±1.42)%增加到(82.76±3.19)%,增幅达到71.42%,益生菌干酪经胃肠道消化后DPPH自由基清除率的变化更加显著,表明益生菌干酪经胃肠道消化后产生了更多具有抗氧化活性的物质,为DPPH自由基提供了氢,使其形成稳定的分子[23]。研究表明,酪蛋白经胰蛋白酶水解后DPPH自由基清除率有所升高,由15%提高到50%[24],同时多肽的浓度和分子质量均显著影响自由基清除率,多肽分子质量越小,对生物体内自由基清除效果越好,清除率越高[25]。因此本研究中益生菌干酪经胃肠道消化后DPPH自由基清除率的增加,有可能是由于干酪中的酪蛋白在胃蛋白酶、胰蛋白酶以及益生菌产生的蛋白酶作用下,降解生成了一些具有抗氧化活性的多肽。
2.1.2 还原力变化
还原能力是表征物质在氧化还原反应中给出电子,自身发生氧化的能力,是物质具有抗氧化活性的重要表现[26]。
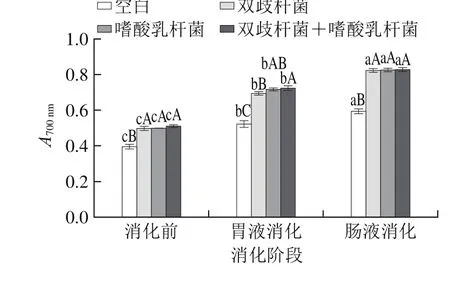
图2 干酪经模拟胃肠道消化还原力变化Fig. 2 Change in reducing power of cheese after simulated gastrointestinal digestion
由图2可知,消化前空白组、各实验组益生菌干酪的还原力分别为0.394±0.035、0.496±0.032、0.497±0.009、0.510±0.007,经过6 h的模拟胃肠道消化后各组干酪的还原力显著提高(P<0.05),分别增加到0.593±0.042、0.823±0.025、0.826±0.019、0.828±0.021,增幅分别为50.51%、65.93%、66.20%、62.35%。还原力的显著提高可能是由于益生菌干酪经模拟胃肠道消化后产生更多的物质提供氢原子,将Fe3+还原成Fe2+,从而打断自由基连锁反应[27]。有研究表明酸乳中分子质量大于30 kDa和在10~30 kDa之间的多肽的还原能力分别为0.028和0.055,明显低于抗坏血酸的0.429;相反,分子质量低于3 kDa的多肽的还原能力达到0.917,要显著高于抗坏血酸,表明不同分子质量的多肽还原力不同,即抗氧化活性也就不同[28]。本研究的4 组干酪中,复合添加双歧杆菌和嗜酸乳杆菌的干酪还原力在各个时期均为最高,但与除空白组外的其他组别无明显差异(P>0.05),此结果表明,单一或复合添加益生菌对干酪还原力的变化影响较小。
2.1.3 ABTS+·清除率变化
ABTS+·也是一种稳定的自由基,根据样品对ABTS+·溶液吸光度的影响测定其对ABTS+·的清除能力,从而评价样品的抗氧化活性[22],干酪消化后ABTS+·清除率的变化如图3所示。
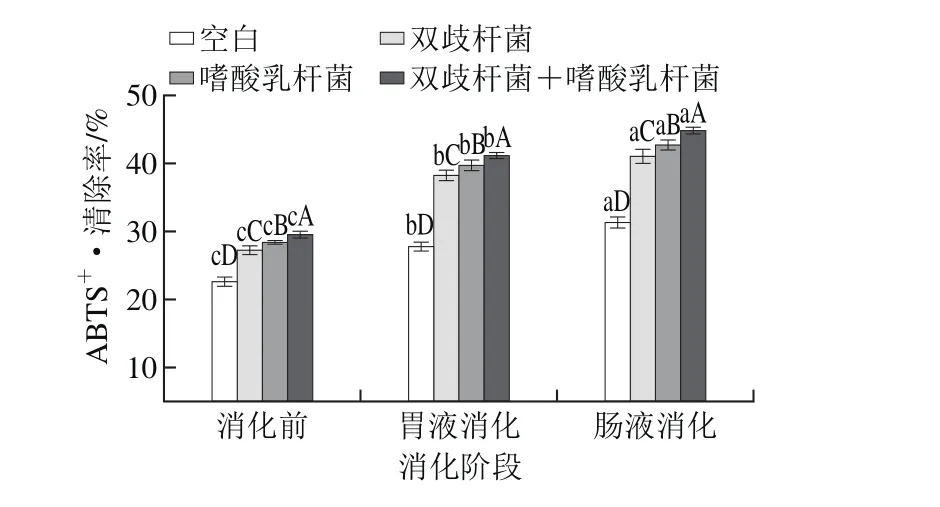
图3 干酪经模拟胃肠道消化ABTS+ ·清除率变化Fig. 3 Change in ABTS+· scavenging activity of cheese after simulated gastrointestinal digestion
由图3可知,空白组及各实验组益生菌干酪经模拟胃肠道消化后ABTS+·清除率均显著提高(P<0.05),各益生菌干酪ABTS+·清除率均显著高于空白组(P<0.05)。复合添加双歧杆菌和嗜酸乳杆菌的干酪ABTS+·清除率为最高(P<0.05),从消化前的(29.56±1.43)%增加到模拟胃肠道消化后的(44.84±1.36)%,增幅达到51.69%。Corrêa等[27]研究表明羊乳酪乳清经6 h蛋白酶水解后,ABTS+·清除率同样有所提升,从15.98%上升到55.30%。本研究中各实验组益生菌干酪在各阶段的ABTS+·清除率均有显著性差异,表明不同的益生菌对干酪ABTS+·清除率有显著不同影响,这与Mushtaq等[10]研究结果相一致。此外,研究已证实一些氨基酸如组氨酸(由于其咪唑环分解后可作为氢受体,捕捉自由基)、酪氨酸、蛋氨酸和半胱氨酸都具有较强的自由基清除能力[29]。因此本研究中干酪模拟胃肠道消化后ABTS+·清除率的升高,也可能与产生了这类氨基酸有关。
2.2 干酪经模拟胃肠道消化益生菌活菌数变化
图4显示了干酪经模拟胃肠道消化后益生菌活菌数的变化。消化前空白组、各实验组益生菌干酪中益生菌活菌数分别为(4.67±0.13)、(7.52±0.26)、(7.48±0.29)、(7.62±0.03)lg(CFU/g),模拟胃肠道消化后活菌数有所降低,分别为(3.68±0.11)、(6.37±0.12)、(6.19±0.36)、(6.33±0.14)lg(CFU/g),分别降低了21.20%、15.29%、17.25%、16.93%。本研究结果表明干酪中的益生菌经模拟胃肠道消化后有良好的存活,原因可能是益生菌受到干酪蛋白质-脂肪基质的保护作用[19]。本研究结果显示益生菌活菌数经模拟胃肠道消化后显著减少,而2.1节中的研究表明模拟胃肠道消化后抗氧化活性显著升高,可以说明在模拟胃肠道环境下益生菌菌体本身的功能特性对干酪抗氧化活性的增加作用较小,抗氧化活性变化的原因仍需进一步研究。
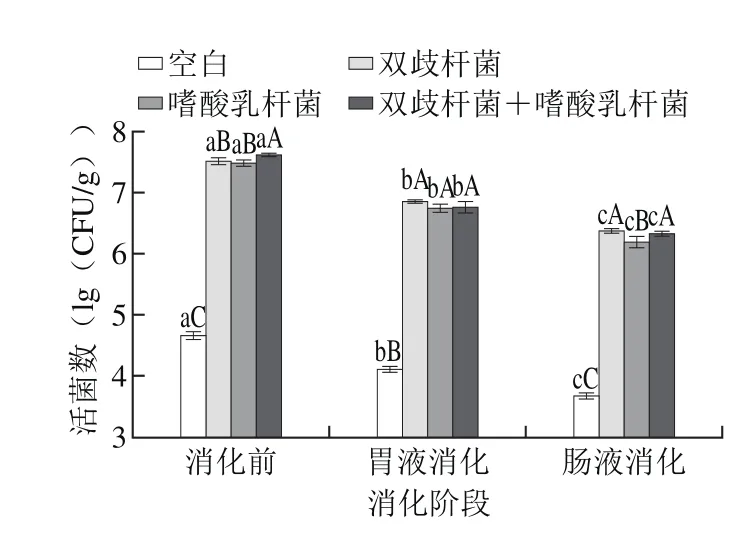
图4 干酪经模拟胃肠道消化益生菌活菌数变化Fig. 4 Change in viable counts of probiotic bacteria in cheese after simulated gastrointestinal digestion
2.3 干酪经模拟胃肠道消化多肽的变化
2.3.1 干酪SDS-PAGE结果
SDS-PAGE具有分子筛效应,根据蛋白分子质量亚基的不同,能够有效分离乳中的主要蛋白质,达到定性和定量的目的[20]。
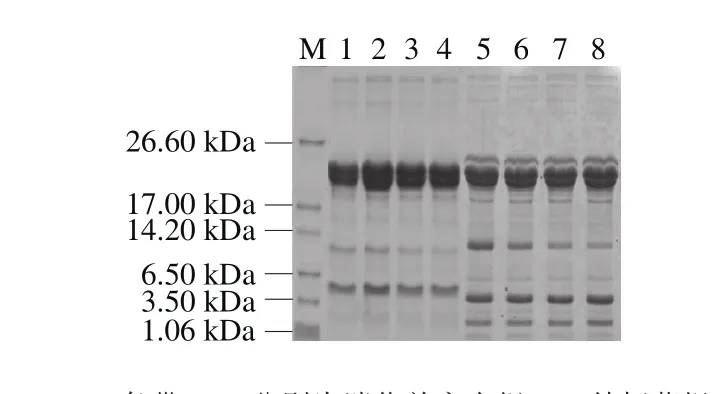
图5 干酪模拟胃肠道消化前后的SDS-PAGEFig. 5 SDS-PAGE of cheese proteins before and after simulated gastrointestinal digestion
图5为干酪模拟胃肠道消化前后的电泳结果,模拟胃肠道消化前各组均有两条分子质量较大的条带,有学者指出,这两条大的条带可能是αs1-酪蛋白和β-酪蛋白,它们的分子质量和条带相接近,分别为23.6 kDa和24.0 kDa,这两种酪蛋白分别占牛乳酪蛋白的36%和30%[13]。Sánchez-Rivera等[30]研究表明,干酪消化后小肽的数量有所增加,且主要由αs1-酪蛋白和β-酪蛋白产生。本研究中模拟胃肠道消化后,两条大的条带仍存在,但是染色情况变弱,在分子质量为1.06~14.2 kDa区间出现了3 条清晰的条带,分子质量分别为2、4、10 kDa左右,表明干酪经模拟胃肠道消化后产生更多小分子质量多肽。
2.3.2 干酪经模拟胃肠道消化多肽含量变化
以谷胱甘肽为标准物质,在540 nm波长处建立谷胱甘肽质量浓度与吸光度之间的关系,回归方程为y=0.054 8x-0.008 1(R2=0.996 2),线性关系良好,可用于计算干酪中多肽含量。干酪消化后多肽含量的变化如图6所示。
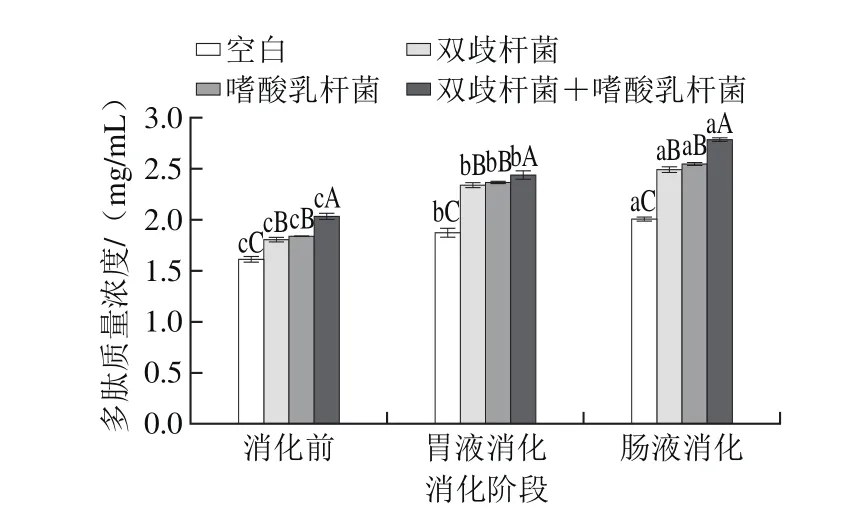
图6 干酪经模拟胃肠道消化多肽质量浓度变化Fig. 6 Change in polypeptide content in cheese after simulated gastrointestinal digestion
由图6可知,消化前空白组、各实验组益生菌干酪提取液中多肽质量浓度分别为(1.62±0.09)、(1.81±0.07)、(1.84±0.01)、(2.01±0.06)mg/mL,模拟胃肠道消化后多肽质量浓度显著升高(P<0.0 5),分别达到(2.0 1±0.0 6)、(2.50±0.10)、(2.55±0.07)、(2.80±0.02)mg/mL,增幅分别为24.07%、38.12%、38.59%、39.30%,表明干酪在模拟胃肠道消化过程中,胃蛋白酶和胰蛋白酶对干酪蛋白质降解作用显著,产生更多的生物活性肽。Sánchez-Rivera等[30]研究指出,蓝纹干酪中存在ACE抑制肽和阿片肽,经胃肠道消化后会产生更多的肽类,比如抗氧化肽和抗菌肽。本研究的实验组益生菌干酪中,复合添加双歧杆菌和嗜酸乳杆菌的干酪中多肽含量在各个阶段均为最高,且各实验组益生菌干酪多肽含量都显著高于空白组干酪(P<0.05)。乳杆菌已经被证明具有产生二肽酶、羧肽酶、三肽酶、氨肽酶和肽链内切酶的能力[31],因此本研究中双歧杆菌及嗜酸乳杆菌的蛋白酶可能在蛋白水解中发挥了作用[32]。此外,研究表明,酪蛋白多肽的抗氧化活性不仅与多肽的质量浓度有关,还与多肽分子质量的大小有关[23,25],因此,有必要研究不同分子质量多肽的抗氧化活性。
2.3.3 所产多肽抗氧化活性的分析
利用超滤膜将模拟胃肠道消化后复合添加双歧杆菌和嗜酸乳杆菌的干酪多肽进行分离,得到3 个不同分子质量的组分,组分Ⅰ(>10 kDa)、Ⅱ(3~10 kDa)、Ⅲ(<3 kDa)。由表1可知,不同分子质量的多肽组分其抗氧化指标均存在显著性差异(P<0.05)。分子质量小于3 kDa的多肽组分各抗氧化指标均为最高,DPPH自由基清除率、还原力、ABTS+·清除率分别为(96.47±2.06)%、0.917±0.078、(83.58±4.83)%,显著高于超滤前样品的各抗氧化指标(P<0.05)。本研究结果表明分子质量小于3 kDa的多肽在干酪中是最主要的抗氧化组分,模拟胃肠道消化后干酪抗氧化活性的提高很可能与这类具有高抗氧化活性的小分子质量多肽有关。
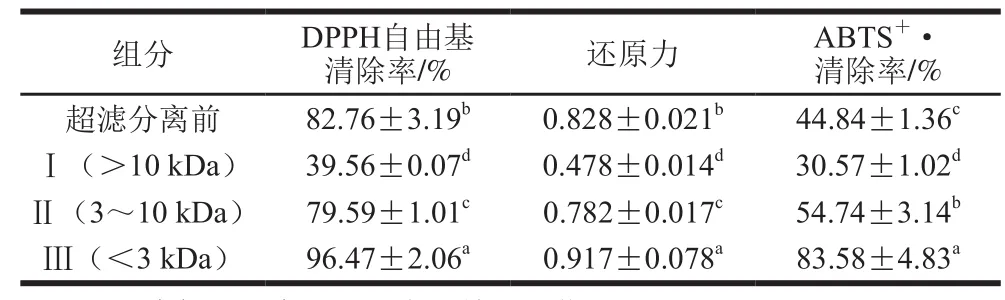
表1 不同分子质量多肽抗氧化活性比较Table 1 Comparison of antioxidant properties of polypeptides with different molecular weights
3 结 论
本研究得出,模拟胃肠道消化后益生菌干酪抗氧化活性显著提高(P<0.05),其中复合添加双歧杆菌、嗜酸乳杆菌的干酪抗氧化指标显著高于其他组(P<0.05);消化后干酪中益生菌活菌数降低,而多肽含量显著增加(P<0.05),其中分子质量小于3 kDa的多肽抗氧化活性最高,表明干酪在益生菌蛋白酶、胃蛋白酶和胰蛋白酶的作用下产生了更多具有抗氧化活性的小分子多肽,使其抗氧化活性得以提高。而对于益生菌干酪在体内代谢环境下抗氧化活性的变化及机制则有待进一步研究。
[1] REALE A, IANNIELLO R G, CIOCIA F, et al. Eあect of respirative and catalase-positive Lactobacillus casei adjuncts on the production and quality of Cheddar-type cheese[J]. International Dairy Journal,2016, 63: 78-87. DOI:10.1016/j.idairyj.2016.08.005.
[2] MURTAZA M A, UR-REHMAN S, ANJUM F M, et al. Cheddar cheese ripening and flavor characterization: a review[J]. Critical Reviews in Food Science and Nutrition, 2014, 54(10): 1309-1321.DOI:10.1080/10408398.2011.634531.
[3] 王娉婷. 传统发酵食品来源的功能性益生菌筛选及其在干酪加工中的应用[D]. 长春: 吉林农业大学, 2015: 1-44.
[4] 罗洁, 任发政, 王紫薇, 等. 干酪质构与风味控制技术研究进展[J]. 农业机械学报, 2016, 47(1): 190-201; 208. DOI:10.6041/j.issn.1000-1298.2016.01.025.
[5] LU Y, GOVINDASAMY-LUCEY S, LUCEY J A. Angiotensin-Ⅰ-converting enzyme-inhibitory peptides in commercial Wisconsin Cheddar cheeses of diあerent ages[J]. Journal of Dairy Science, 2016,99(1): 41-52. DOI:10.3168/JDS.2015-9569.
[6] CAGGIA C, DE ANGELIS M, PITINO I, et al. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses[J]. Food Microbiology, 2015, 50: 109-117. DOI:10.1016/j.fm.2015.03.010.
[7] SAH B N P, VASILJEVIC T, MCKECHNIE S, et al. Effect of probiotics on antioxidant and antimutagenic activities of crude peptide extract from yogurt[J]. Food Chemistry, 2014, 156: 264-270.DOI:10.1016/j.foodchem.2014.01.105.
[8] 贾宏信, 龚广予, 郭本恒. 益生菌干酪的研究进展[J]. 食品科学,2013, 34(15): 355-360. DOI:10.7506/spkx1002-6630-201315072.
[9] ABADÍA-GARCÍA L, CARDADOR A, DEL CAMPO S T M, et al.Inぼuence of probiotic strains added to cottage cheese on generation of potentially antioxidant peptides, anti-listerial activity, and survival of probiotic microorganisms in simulated gastrointestinal conditions[J].International Dairy Journal, 2013, 33(2): 191-197. DOI:10.1016/j.idairyj.2013.04.005.
[10] MUSHTAQ M, GANI A, MASOODI F A, et al. Himalayan cheese(Kalari/Kradi): effect of different probiotic strains on oxidative stability, microbiological, sensory and nutraceutical properties during storage[J]. LWT-Food Science and Technology, 2016, 67: 74-81.DOI:10.1016/j.lwt.2015.11.039.
[11] GUPTA A, MANN B, KUMAR R, et al. Identiベcation of antioxidant peptides in cheddar cheese made with adjunct culture Lactobacillus casei ssp. Casei 300[J]. Milchwissenschaft-Milk Science International,2010, 65(4): 396-399.
[12] CHAVES K S, GIGANTE M L. Prato cheese as suitable carrier for Lactobacillus acidophilus La5 and Bifidobacterium Bb12[J].International Dairy Journal, 2016, 52: 10-18. DOI:10.1016/j.idairyj.2016.08.009.
[13] PARROT S, DEGRAEVE P, CURIA C, et al. In vitro study on digestion of peptides in Emmental cheese: analytical evaluation and influence on angiotensin Ⅰ converting enzyme inhibitory peptides[J]. Molecular Nutrition & Food Research, 2003, 47(2): 87-94.DOI:10.1002/food.200390032.
[14] AYALA-BRIBIESCA E, LUSSIER M, CHABOT D, et al. Effect of calcium enrichment of Cheddar cheese on its structure, in vitro digestion and lipid bioaccessibility[J]. International Dairy Journal,2016, 53: 1-9. DOI:10.1016/j.idairyj.2015.09.002.
[15] MOZZI F, GERBINO E, DE VALDEZ G F, et al. Functionality of exopolysaccharides produced by lactic acid bacteria in an in vitro gastric system[J]. Journal of Applied Microbiology, 2009, 107(1): 56-64. DOI:10.1111/j.1365-2672.2009.04182.x.
[16] BOTTESINI C, PAOLELLA S, LAMBERTINI F, et al. Antioxidant capacity of water soluble extracts from Parmigiano-Reggiano cheese[J]. International Journal of Food Sciences and Nutrition, 2013,64(8): 953-958. DOI:10.3109/09637486.2013.821696.
[17] SHEN Yingbin, ZHANG Hui, CHENG Liling, et al. In vitro and in vivo antioxidant activity of polyphenols extracted from black highland barley[J]. Food Chemistry, 2016, 194: 1003-1012. DOI:10.1016/j.foodchem.2015.08.083.
[18] DUH P D, TU Y Y, YEN G C. Antioxidant activity of water extract of Harng Jyur (Chrysanthemum morifolium Ramat)[J]. LWT-Food Science and Technology, 1999, 32(5): 269-277. DOI:10.1006/fstl.1999.0548.
[19] DE OLIVEIRA M E G, GARCIA E F, DE OLIVEIRA C E V, et al.Addition of probiotic bacteria in a semi-hard goat cheese (coalho):survival to simulated gastrointestinal conditions and inhibitory eあect against pathogenic bacteria[J]. Food Research International, 2014, 64:241-247. DOI:10.1016/j.foodres.2014.06.032.
[20] 金江玉, 宋子明, 史海莹, 等. SDS-PAGE凝胶电泳在鉴别水解蛋白掺假液态乳中的应用[J]. 中国乳品工业, 2012, 40(7): 4-7.
[21] 徐娟, 吕嘉枥. 乳蛋白水解液中多肽含量测定方法的研究[J]. 食品科技, 2010, 35(12): 275-278. DOI:10.13684/j.cnki.spkj.2010.12.051.
[22] 曾维才, 石碧. 天然产物抗氧化活性的常见评价方法[J].化工进展, 2013, 32(6): 1205-1213; 1247. DOI:10.3969/j.issn.1000-6613.2013.06.001.
[23] TIMÓN M L, PARRA V, OTTE J, et al. Identification of radical scavenging peptides (< 3 kDa) from Burgos-type cheese[J]. LWTFood Science and Technology, 2014, 57(1): 359-365. DOI:10.1016/j.lwt.2014.01.020.
[24] LUO Y C, PAN K, ZHONG Q X. Physical, chemical and biochemical properties of casein hydrolyzed by three proteases: partial characterizations[J]. Food Chemistry, 2014, 155: 146-154.
[25] 陈东平, 牟光庆. 不同分子量酪蛋白肽对自由基清除作用研究[J].中国酿造, 2010, 29(4): 33-35.
[26] 曾军, 石国荣. 天然产物抗氧化活性的测定方法和原理[J]. 安徽农学通报, 2008, 14(22): 35-36. DOI:10.16377/j.cnki.issn1007-7731.2008.22.001.
[27] CORRÊA A P F, DAROIR D J, FONTOURA R, et al. Hydrolysates of sheep cheese whey as a source of bioactive peptides with antioxidant and angiotensin-converting enzyme inhibitory activities[J]. Peptides,2014, 61: 48-55. DOI:10.1016/j.peptides.2014.09.001.
[28] FARVIN K H S, BARON C P, NIELSEN N S, et al. Antioxidant activity of yoghurt peptides: part 1-in vitro assays and evaluation in ω-3 enriched milk[J]. Food Chemistry, 2010, 123(4): 1081-1089.DOI:10.1016/j.foodchem.2010.05.067.
[29] MEIRA S M M, DAROIT D J, HELFER V E, et al. Bioactive peptides in water-soluble extracts of ovine cheeses from Southern Brazil and Uruguay[J]. Food Research International, 2012, 48(1): 322-329.DOI:10.1016/j.foodres.2012.05.009.
[30] SÁNCHEZ-RIVERA L, DIEZHANDINO I, GÓMEZ-RUIZ J Á, et al.Peptidomic study of Spanish blue cheese (Valdeón) and changes after simulated gastrointestinal digestion[J]. Electrophoresis, 2014, 35(11):1627-1636. DOI:10.1002/elps.201300510.
[31] PETERSON S D, MARSHALL R T. Nonstarter Lactobacilli in Cheddar cheese: a review[J]. Journal of Dairy Science, 1990, 73(6):1395-1410. DOI:10.3168/jds. S0022-0302(90)78804-2.
[32] OH N S, JOUNG J Y, LEE J Y, et al. Characterization of the microbial diversity and chemical composition of Gouda cheese made by potential probiotic strains as an adjunct starter culture[J]. Journal of Agricultural and Food Chemistry, 2016, 64(39): 7357-7366. DOI:10.1021/acs.jafc.6b02689.
