基因生物信息学的脓毒症潜在发病机制研究及生物标记物筛选*
2018-02-28王海清陈睦虎胡迎春杨蕊萍
王海清,左 文,陈睦虎,胡迎春,杨蕊萍,钟 武
脓毒症是指机体对感染的反应失调而导致危及生命的器官功能障碍[1]。病情危重时可导致多器官功能障碍综合征(MODS)和(或)循环衰竭,是临床上常见的一种急危重症[2~5]。每年全球有超过180万重症脓毒症的病例,其发病速度快,临床病死率高,若合并感染性休克,病死率约80%[6]。目前认为,微循环障碍是发生MODS甚至死亡的主要因素,其中内皮细胞以及相关促凝效应等起决定性作用,但其具体分子机制仍不明确。因脓毒症是机体应对感染所发生的一系列复杂的免疫反应,早期主要表现为强力抗感染,晚期主要表现为剧烈而持久的免疫抑制反应,这表明多种生物标记物或许可以用来早期识别脓毒症[7]。而脓毒症治疗效果差的根本原因在于目前脓毒症发病机制尚不清楚,用于脓毒症临床诊断及判断疗效的指标欠缺,尤其需要获取其早期诊断指标和治疗作用的关键靶点。随着基因芯片技术的发展及应用,找出的差异基因成千上万,如何在众多差异基因中筛选出对疾病起关键作用的基因已成为技术瓶颈,生物信息学技术在此基础上应运而生,可以帮助研究者快速找到核心基因群。
本文拟基于GEO基因表达数据,全面分析脓毒症疾病状态,从而构建基因网络,并筛选潜在的关键分子靶点,这些分子靶点可能为我们寻找脓毒症发病机理提供新的见解,或许可以用于早期诊断脓毒症并为临床治疗脓毒症提供新思路。
1 材料和方法
1.1 数据来源
GEO数据库隶属于美国国立卫生研究院的美国国立生物技术信息中心(NCBI),是目前全球最大、最完整的公共基因表达数据资源[8]。基因表达谱GSE28750下载于GEO数据库,种属为人类,芯片平台GPL570[HG-U133_Plus_2]Affymetrix Human Ge⁃nome U133 Plus 2.0 Array。该芯片数据包括10例脓毒症患者及19例健康人的基因表达阵列数据[9]。
1.2 差异表达基因处理
GCBI(Gene-Cloud of Biotechnology Information,网址:https://www.gcbi.com.cn)是我国研发的基于R语言的在线分析软件,能便捷的对芯片数据进行相关处理。通过GCBI软件,对基因芯片数据先进行对数化标准处理以利于分析,再筛选差异表达基因,筛选条件为:①P<0.01;②差异倍数≥2。
1.3 基因本体论分析
基因本体论(GO)数据库从功能、参与的生物途径及细胞中的定位三个方面对基因产物进行了标准化描述(即对基因产物进行了简单注释)。通过GO富集分析可以大致明了差异表达基因在哪些生物学功能、途径或者细胞定位方面进行富集[10]。通过GC⁃BI平台对上传的差异表达基因进行GO分析,选择差异最显著的10个功能进行排序并分析。
1.4 pathway分析
KEGG(京都基因与基因组百科全书,网址:http://www.genome.jp/kegg/)是日本京都大学生物信息学中心的Kanehisa实验室建立的以基因组、化学和系统功能信息为基础、全面展示生物及细胞所包括的生物学信息的数据库,它可以预测蛋白质在各种细胞活动中所起作用并将其绘制成网络。通过对差异表达基因进行信号通路分析,可以了解疾病状态下显著改变的代谢通路,这对于实验机制的探索意义重大。基于KEGG数据库对差异表达基因进行基因信号通路富集分析,选择差异最显著的10个功能进行排序并分析。
1.5 信号网络分析
基因信号网络分析是基于KEGG数据库中蛋白和蛋白之间的相互作用关系而构建的基因间相互作用的网络图。它解构了KEGG数据库,克服了KEGG-Pathway数据库中基因间相互作用关系受限于某一Pathway的局限,因而,基因信号网络是在整个KEGG-Pathway数据库中筛选某个蛋白的上游及下游蛋白。对差异表达基因使用基于KEGG的GC⁃BI平台进行基因信号网络分析,并绘制成网络,以便了解差异基因之间的相互调控关系。
1.6 共表达网络分析
共表达网络分析通过基因表达的相似性简单直观的分析基因产物可能存在的相互作用关系,从而更深入的发现关键调控基因及其相互作用脉络。对差异表达基因使用GCBI平台进行共表达分析,并绘制成网络,推测其潜在的关键靶点。
2 结 果
2.1 差异表达基因筛选结果
对数化标准处理前后的脓毒症组和健康对照组基因芯片数据结果(见图1~2),可知本研究标本经对数化处理后数据均一性较好,两组之间具有可比性。
图1 样本芯片数据对数化标准处理前
2.2 差异基因筛选结果
通过对两组数据的差异表达基因进行筛选,共获得2 457个差异表达基因,与对照组相比,脓毒症组上调基因1 282个、下调基因1 175个(见图3)。由图可知,我们筛选的差异基因能较好的区分出2个组。
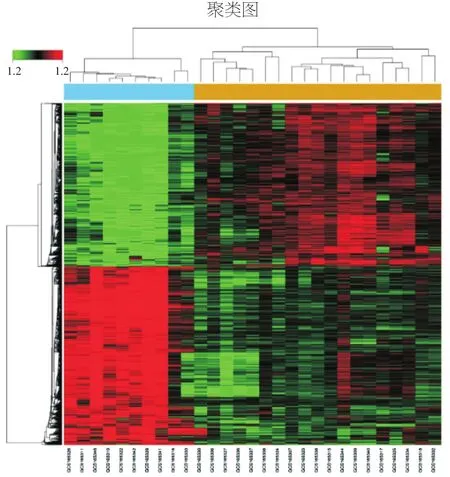
图3 差异表达基因聚类图
2.3 差异表达基因GO分析结果
采用GCBI平台对差异表达基因进行GO功能富集分析,其中最显著的10功能依次为:T细胞共刺激、T细胞受体信号通路、细胞表面受体信号通路、免疫反应、翻译、固有免疫反应、血液凝固、基因表达、小分子物质代谢过程、信号转导(见图4)。

图2 样本芯片数据对数化标准处理后
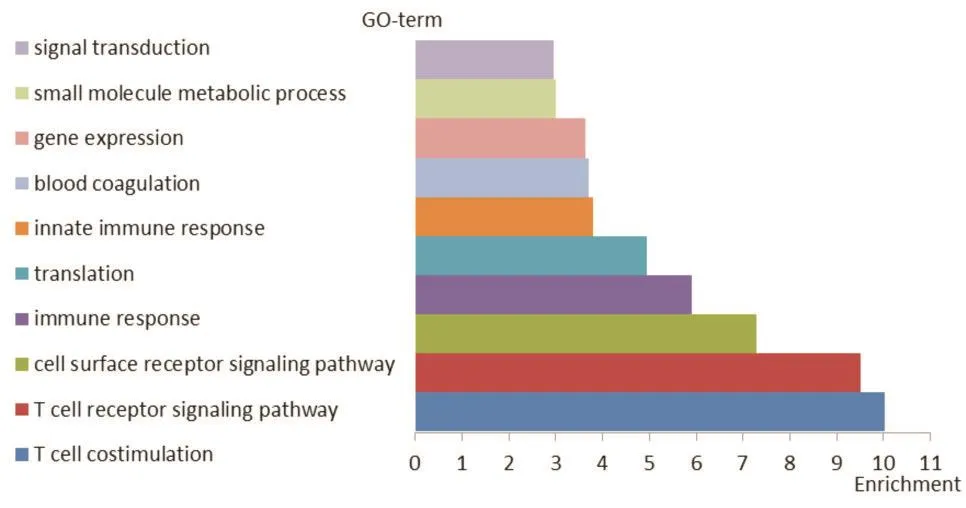
图4 差异表达基因G0分析结果图
2.4 差异表达基因pathway分析结果
表1列出了15个差异最显著的富集通路,其主要涉及凋亡、细胞周期、免疫反应、信号转导等。
2.5 差异表达基因信号网络分析结果
图5显示了前20个网络的中心节点基因。由图可知,GNAI3、PIK3CB、MAPK14、IL8为其关键基因。
2.6 差异表达基因共表达分析
使用GCBI对差异表达基因进行共表达分析,其中前216个节点共表达图见图6,由图可发现,上调和下调基因分别呈聚集性分部,符合共表达理论。其中前15个节点名称、功能见表2。
3 讨 论
脓毒症是临床常见且死亡率极高的急危重症,是一个复杂的病理过程,在此过程中,病原微生物作为始发因素,其后多种促炎细胞因子、抗炎细胞因子、炎性细胞、信号通路等共同参与其中[9]。已有研究者报道脓毒症的发生机制,但很少涉及其分子标记物,事实上,分子生物标记物对于脓毒症的诊断、监测及预后具有很大的潜力[11]。

表1 差异表达基因pathway分析
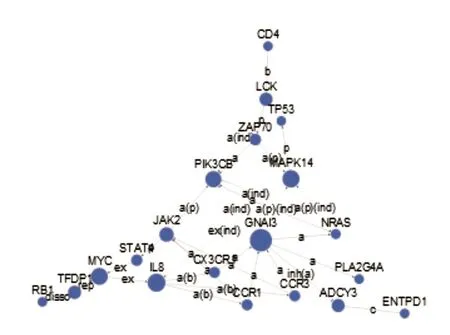
图5 差异表达基因信号网络分析

图6 差异表达基因前216个节点共表达图

表2 差异表达基因共表达显著差异前15个节点
本研究中通过使用微阵列的方法对脓毒症组及健康对照组基因芯片进行分析比较,共筛选出2 457个差异表达基因,其中上调基因1 282个,主要涉及细胞膜稳定性、基因转录及翻译过程、物质代谢的调节、免疫反应等方面;下调基因1 175个,主要涉及免疫反应、信号转导、细胞增殖、细胞凋亡等过程。通过对差异表达基因进行GO分析发现,二者差异最显著的功能主要涉及固有免疫反应、信号转导、物质代谢、基因的转录及翻译等方面。通过对二者进行pathway分析发现,其差异最显著富集通路仍主要涉及物质代谢、基因表达、免疫反应等方面。将基因间相互作用形成网络后可见脓毒症组及健康对照组之间差异也主要集中在信号转导、物质代谢、免疫反应、基因表达等方面,其中MAPK signaling pathway在网络图中有38个上游信号通路、5个下游信号通路,Apoptosis有27个上游信号通路、3个下游信号通路,以此推测脓毒症患者死亡率高可能和免疫细胞凋亡调节异常有关。
其次,通过対差异表达基因进行了基因信号间相互作用分析,分析发现:GNAI3上游有11个基因与其联系,下游有9个基因与其联系,GNAI3功能主要涉及细胞周期、血液凝固、突触传递、细胞分裂等[12],推测其为脓毒症发病过程中的关键基因;PIK3CB上游有24个基因与其联系,下游有6个基因与其联系,其功能主要涉及内皮细胞增殖、自噬的正调节、血液凝固、适应性免疫应答等[13-15],推测PIK3CB为脓毒症发病过程中的关键基因;MAPK14上游有15个基因与其联系,下游有8个基因与其联系,主要对炎症反应中细胞因子产生的调节、在DNA损伤反应中的信号转导、RNA聚合酶Ⅱ启动子转录的正调控等[16],也可能为脓毒症发病过程中的关键基因;IL8上下游分别有4个及5个基因与其联系,其功能主要参与信号转导、抗感染、中性粒细胞的活化等[17],推测此基因也可能为脓毒症发病过程中的关键基因。
对差异表达基因的共表达分析发现GYG1、SERPINB1、SAMSN1、ATP11B 这四个基因分别有39、39、33、33个基因与其他基因表达呈相同趋势,这些基因的作用主要体现在物质代谢、免疫反应、细胞周期、信号转导等方面[18-20],或许为脓毒症的关键基因。
目前,生物信息学已被广泛应用于建立各种生物数据库并对其进行管理、研发数据库接口及检索工具、从海量数据中提取新知识等方面,更重要的是,生物信息学还可以研究基因表达过程中的聚群关系,并从中提取其调控网络以及代谢途径方面的信息,进而从整体水平模拟细胞内的全部相关的生化反应,从而在亚细胞水平阐释生命活动。GCBI是我国研发的一款发掘生物信息学信息的软件。
4 结论
本研究通过GCBI使用基因信号间相互作用网络分析联合共表达网络分析的方法筛选脓毒症关键基因,既克服了基因信号间相互作用分析缺乏创新性和共表达网络分析可靠性欠佳的缺点,又保留了基因信号间相互作用分析结果真实可靠和共表达网络分析创新性强的优点,二者相互结合能够更好的筛选出受实验因素影响较大的核心基因。但生物信息学仍然需要分子生物学实验以验证其准确性以及后续基因、蛋白功能,生物信息学分析加进一步实验验证必将成为以后科研流行趋势。
1. Singer M,Deutschman CS,Seymour CW,et al.The third international consensus definitions for sepsis and septic shock(sepsis-3)[J].JAMA,2016,315(8):801-810.
2. Ince C.The microcirculation is the motor of sepsis[J].Crit Care,2005,9(4):13-19.
3. M,C,Bellamy.Wet,dry or something else[J].Br J An⁃aesth,2006,97(6):755-757.
4. 王静,李熙鸿.脓毒症发病机制的研究进展[J].实用儿科临床杂志,2012,27(10):786-789.
5. 胡迎春,张千,钟武,等.大鼠脓毒血症相关基因筛选与生物信息学分析[J].泸州医学院学报,2013,36(1):80-82.
6. Galley HF.Oxidative stress and mitochondrial dysfunction in sepsis[J].Br J Anaesth,2011,107(1):57-64.
7. Sutherland A,Thomas M,Brandon RA,et al.Develop⁃ment and validation of a novel molecular biomarker diag⁃nostic test for the early detection of sepsis[J].Crit Care,2011,15(3):R149.
8. 端木欣欣.基于GEO数据库筛选镉致前列腺癌主要环境应答基因的研究[D].北京:中央民族大学,2014:4-8.
9. 栾德琴,白云峰,包文斌,等.基因本体论研究进展及在生物信息数据库中的应用[J].江苏农业科学,2009,(4):14-16.
10. Roos DS.Computational biology.Bioinformatics--try⁃ing to swim in a sea of data[J].Science,2001,291(557):1 260-1 261.
11. Gohla A,Klement K,Piekorz RP,et al.An obligatory requirement for the heterotrimeric G protein Gi3 in the antiautophagic action of insulin in the liver[J].PNAS,2007,104(8):3 004-3 008.
12. Lin ZQ,Zhou PZ,Von Gise A,et al.Pi3kcb links Hip⁃po-YAP and PI3K-AKT signaling pathways to promote cardiomyocyte proliferation and survival[J].Circ Res,2015,116(1):35-U98.
13. Chen HB,Mei L,Zhou LZ,et al.PTEN restoration and PIK3CB knockdown synergistically suppress glioblas⁃toma growth in vitro and in xenografts[J].J Neurooncol,2011,104(1):155-167.
14. Yan F,Tan XX,Wan WF,et al.ErbB4 protects against neuronal apoptosis via activation of YAP/PIK3CB signal⁃ing pathway in a rat model of subarachnoid hemorrhage[J].Exp Neurol,2017,297(297):92-100.
15. Meng W,Veluchamy A.A genome-wide association study suggested that the mitogen-activated protein kinase 14 gene(MAPK14)is associated with diabetic foot ulcer[J].The British journal of dermatology,2017,10(2):157-158.
16. Dakal TC,Kala D,Dhiman G,et al.Predicting the func⁃tional consequences of non-synonymous single nucleotide polymorphisms in IL8 gene[J].Sci Rep,2017,7(1):145-149.
17. Fanin M,Torella,Savarese M,et al.A GYG1 gene mu⁃tations in a family with polyglucosan body myopathy[J].Neurol Genet,2015,1(3):211-216.
18. Glicksman M,Asthana A,Bethesda,et al.Plasma ser⁃pinB1 is related to insulin sensitivity but not pancreatic β-cell function in non-diabetic adults[J].Physiol Rep,2017,5(5):201-210.
19. Yan Y,Zhang L,Xu T,et al.SAMSN1 is highly ex⁃pressed and associated with a poor survival in glioblastoma multiforme[J].PLoS One,2013,8(11):57-63.
20.H MS,S RA,W PA,et al.Reanalysis of ATP11B,a type IV P-type ATPase[J].J Biol Chem,2002,277(12):9 736-9 740.
