14株H3亚型禽流感病毒全基因组序列分析
2018-02-18程善菊李金平侯广宇王素春蒋文明温建新
程善菊,李金平,侯广宇,彭 程,王素春,蒋文明,温建新
(1.青岛农业大学动物医学院,山东 青岛 266109;2.中国动物卫生与流行病学中心,山东 青岛 266032)
禽流感是一种禽类全身性或者以呼吸症状为主的传染病[1],由正黏病毒科A型流感病毒属禽流感病毒(Avian influenza virus,AIV)引起。根据致病力的不同AIV可分为高致病性禽流感病毒和低致病性禽流感病毒[2]。H3亚型AIV分离率较高,属于低致病性禽流感病毒,其致病力虽然低,但危害持久,能够长期的存在于家禽体内[3]。并且,感染范围比较广,不仅能感染禽类,在哺乳动物猪、马、犬等体内也可以分离到[4]。有研究发现,AIV还与人的H3亚型流感病毒有密切联系,1968年发生于香港的人流感病毒HA和PB1基因,就是来源于禽类[5]。另外,有些H3亚型流感病毒能够与H9N2和H5N1亚型流感病毒发生基因重组[6],人源以及禽源流感病毒还能通过猪这个“混合器”发生基因重排[7],基因重排能造成AIV致病性变强,给禽流感的防控带来很大难度。因此,对家禽进行长期的AIV分子流行病学检测,密切关注AIV传播和进化情况,具有重要的公共卫生学意义。
本试验以2016年至2017年在我国部分省市活禽市场的家禽体内分离得到的14株H3亚型禽流感病毒为研究对象,对其进行全基因测序和遗传进化分析,并对H3亚型AIV的遗传进化关系和流行情况进行分析,以期对H3亚型AIV的进化研究提供一些参考依据。
1 材料与方法
1.1 毒株 本试验14株H3亚型禽流感毒株来源省份与具体监测情况见表1,毒株由中国动物卫生与流行病学中心分离鉴定并保存。10日龄SPF鸡胚,购自青岛市城阳区康信种鸡场。
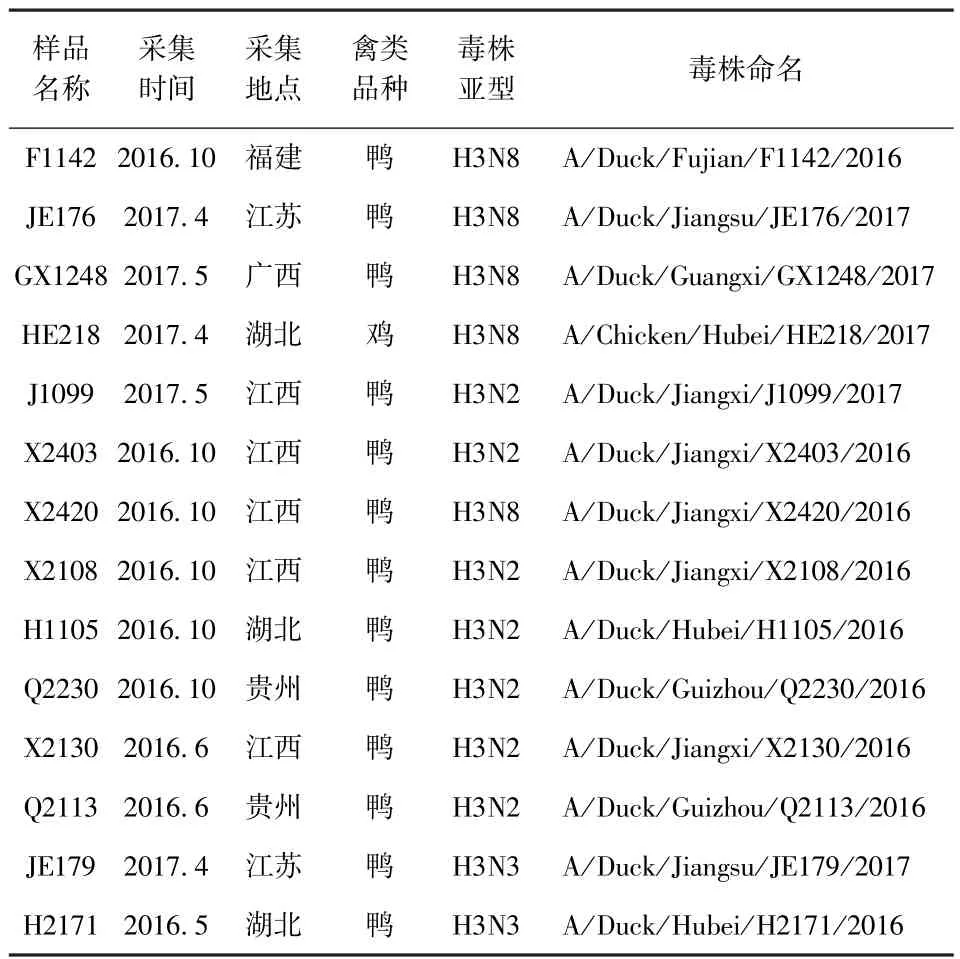
表1 14株H3亚型AIV监测情况
1.2 主要试剂 病毒RNA纯化试剂盒(凯杰企业管理(上海)有限公司);胶回收试剂盒,购自生工生物工程(上海)股份有限公司;连接试剂盒,购自中美泰和生物技术(北京)有限公司;大肠杆菌DH5α购自TaKaRa公司。
1.3 方法
1.3.1 病料采集与处理 采集健康家禽的咽拭子和泄殖腔拭子,并将其放入PBS缓冲液中,充分混匀后抽取2 mL接种10日龄鸡胚,收集在24~96 h死亡及未死亡鸡胚尿囊液,-70℃保存备用。
1.3.2 病毒RNA提取、RT-PCR及胶回收 按照RNA抽提试剂盒提取病毒的RNA,RT-PCR扩增体系采用 25 μL体系,其中 2×One Step Mix为12.5 μL,上下游引物各 1 μL,酶为 1.25 μL,模板为3 μL,纯水6.25 μL。 反应程序:55 ℃ 30 min,94 ℃2 min;94℃30 s,55℃30 s,72℃2 min,35个循环,72℃ 5 min,4℃保存。将25 μL扩增产物在1%琼脂糖凝胶里进行电泳,切取目的条带,按照胶回收试剂盒说明书进行回收纯化。
1.3.3 连接转化 将胶回收后的产物与P Clone EZ-TOPO载体进行连接。然后转入大肠杆菌DH5α感受态细胞中进行转化,挑取单个菌落,进行PCR鉴定,选择阳性克隆菌,将菌液送往青岛睿博兴科生物技术有限公司进行测序。
1.3.4 序列的拼接与分析 采用DNAStar软件中Seqman对测定的各个基因序列进行拼接,用MegAlign软件进行序列比对,得到完整序列后利用软件MEGA 7.0制作进化树并分析。
2 结果
2.1 全基因RT-PCR扩增结果 经1%琼脂糖凝胶电泳鉴定,14株分离株各基因扩增片段与预期扩增大小相符。
2.2 全基因测序结果与序列分析
2.2.1 血凝素(HA)基因序列分析 14株H3亚型禽流感毒株 HA基因HA1区编码344个氨基酸,HA2区编码221个氨基酸,共编码565个氨基酸,没有发生核苷酸插入和缺失的现象。将核苷酸序列翻译为氨基酸序列,结果显示,14株H3亚型分离株HA基因上均存在6个潜在的糖基化位点,分别是位于 HA1 基因上的22/24、38、54、181,301,HA2 基因上499。而且14株毒株HA基因裂解位点氨基酸序列均为340PEKQTR↓GLF348均只有一个碱性氨基酸,具有典型的低致病性禽流感病毒特征。14株毒株的HA基因氨基酸位点在226、228分别为Q、G(人源为226(L)、228(S)),符合典型的禽源特征[8]。另外,位点190(E)和225(G)是维持唾液酸α-2,3-半乳糖受体重要结构的关键位点,并未发生突变,有关HA基因特征性位点详见表2。
遗传进化分析表明,14株毒株之间核苷酸同源性在82.9% ~99.3%之间,其氨基酸序列同源性为91.2% ~98.5%。HA基因进化树如图1,观察进化树可知14株毒株HA基因均属于欧亚分支,在欧亚分支中又分出3个独立小分支。毒株 JE176、JE179、H1105、X2108、X2130、HE218、GX1248 在同一分支,核苷酸同源性为88.2% ~98.7%,其中毒株JE179与安徽参考毒株A1651同源性最高,为94%。 Q2113、Q2230、X2403、H2171、F1142、X2420在同一分支,核苷酸同源性为87.1% ~99.3%,其中毒株H2171与蒙古参考毒株2076同源性最高,为96.5%。毒株J1099单独一个分支,与广西参考毒株175D12同源性最高,为94.5%。14株毒株在相近年份的同源性较高显示出一定的年代性,遗传进化上与猪源人源不在同一分支,说明亲缘关系较远。
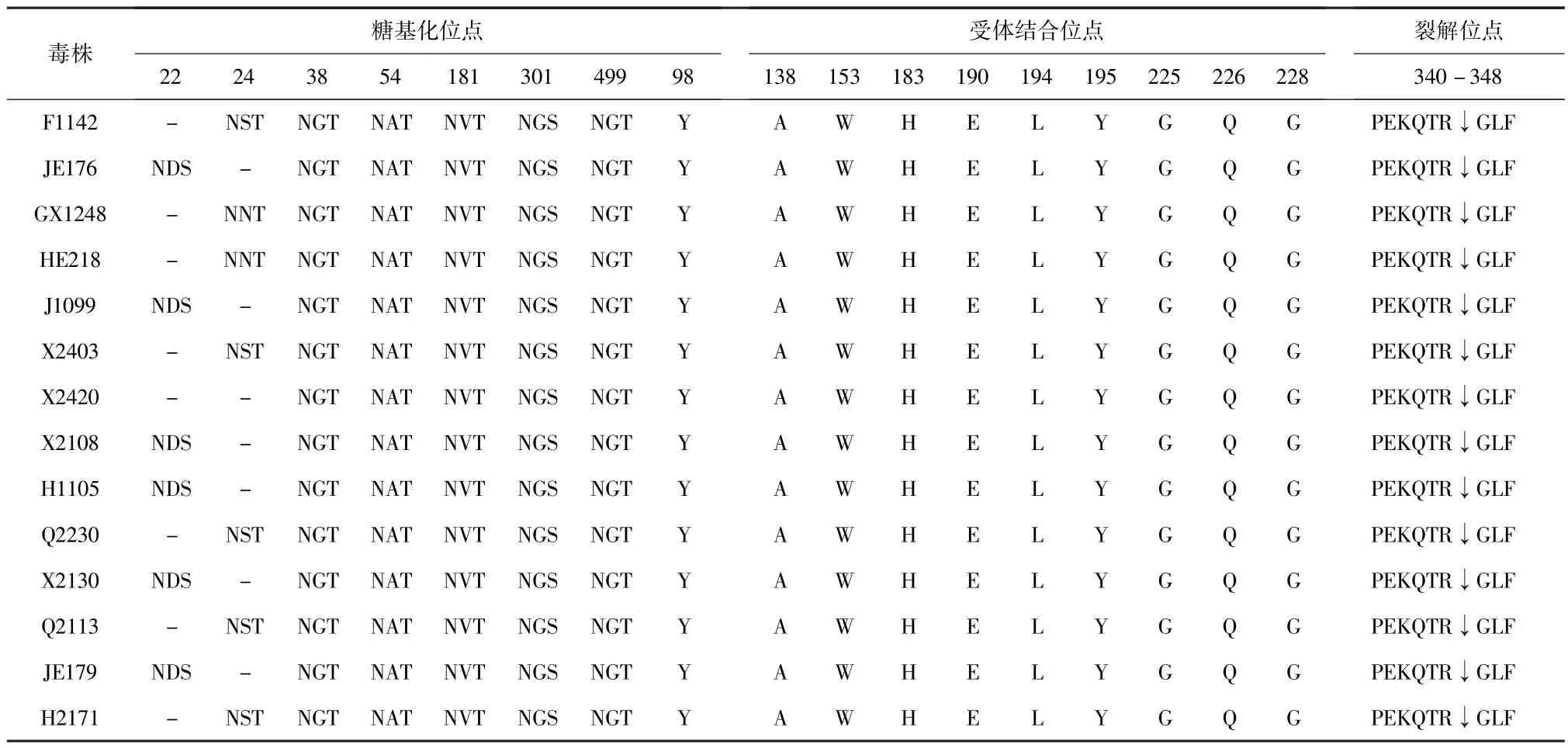
表2 14株H3亚型分离毒株HA基因特征性位点

图1 HA基因遗传进化树
2.2.2 神经氨酸酶(NA)基因序列分析 NA基因共编码470个氨基酸,没有发生核苷酸缺失现象。14株H3亚型分离株中有7株H3N2,5株H3N8,2株H3N3,在蛋白质氨基酸序列中,预测其潜在的糖基化位点,其中7株H3N2中X2130、Q2230、Q2113、X2403、X2108、J1099均存在6个潜在糖基化位点,在氨基酸序列中位点为 61、69、86、20、234、402,其中J1099在第69位为 NST其余均为NNT,毒株H1105在306位增加了一个糖基化位点NMT。5株H3N8中 X2420、GX1248、HE218、F1142 均存在 6个潜在糖基化位点,在氨基酸序列中位点分别为46、54、67、84、144、293,毒株 JE176 糖基化位点在42位增加了一个 NES。在2株 H3N3中毒株JE179 在位点 14、57、66、72、146、235、307 存在 6个潜在糖基化位点,毒株H2171糖基化位点在23位增加了一个 NGT。氨基酸位点119E、152R、274H、292R、294N是NA基因可能的耐药性位点,均未发生变异。
14株毒株中7株H3N2核苷酸同源性在88.2% ~99.8% 之间,氨基酸序列同源性在88.2% ~99.8%之间。5株H3N8核苷酸同源性在79.1% ~99.0% 之间,氨基酸序列同源性78.6% ~98.0%。2株H3N3核苷酸同源性为96.0%,氨基酸序列同源性为97.5%。14株分离株NA基因遗传进化树如图2,不同亚型之间共分为3个小分支,其中Q2230与湖南参考毒株139同源性最高,为95%。H2171与江西参考毒株26175同源性最高,为96.5%。HE218与湖南参考毒株S1824同源性最高,为97.7%。处于同一分支的毒株亲缘关系较近,可能来源于相同祖先。

图2 NA基因遗传进化树
2.2.3 内部基因序列特殊位点分析 对14株H3亚型分离株的内部基因序列进行分析,结果表明,PB2基因编码区由2280个核苷酸组成,共编码759个氨基酸。PB2基因的第627(E)和701(D)是哺乳动物适应性关键位点,与人源的627(K)和701(N)不同[9]。PB1的编码区共有2 274个核苷酸,编码757个氨基酸,PB1-F2蛋白共有90个氨基酸编码,没有发生终止密码子的提前终止[10]。PB1基因的位点198(K)和317(M)被认为与小鼠的致病性有关[11]。PA基因第97位为苏氨酸(Thr)而不是异亮氨酸(Ile),异亮氨酸(Ile)可以使毒力显著增强,对病毒的毒力有非常大的影响[12]。14株分离株M1基因编码252个氨基酸,M2基因编码97个氨基酸,M1相对保守,M2蛋白所含的特异性氨基酸 11(T)、14(G)、16(E)、20(S)和57(Y)未发生变异符合禽流感病毒特征。NS1蛋白的第92位氨基酸为天冬氨酸(Asp)而不是谷氨酸(Glu),谷氨酸(Glu)对拮抗干扰素和肿瘤坏死因子介导的抗病毒反应具有重要作用[13]。
遗传分析表明,14株分离株核苷酸同源性分别为,PB2基因92.9% ~100%,PB1基因87.0% ~97.4%,PA基因85.4% ~97%,NP基因87.8% ~99.7%,NS基因71.5%~99.5%,M基因92.9%~100%。各个内部基因进化树如图3、4、5、6、7、8,其中毒株 A/Duck/Fujian/F1142/2016(H3N8)的NS基因为北美分支,其余毒株均为欧亚分支。
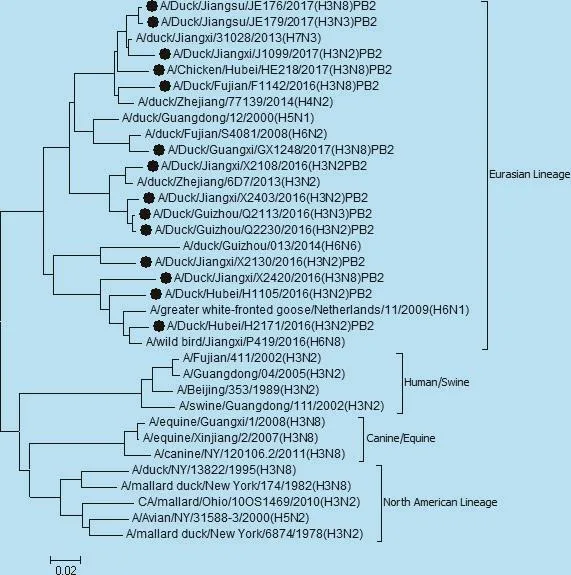
图3 PB2基因遗传进化树
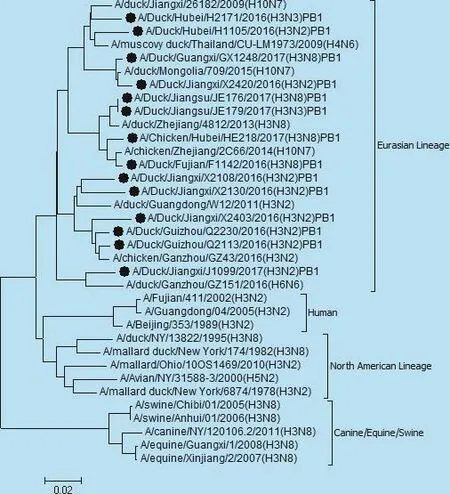
图4 PB1基因遗传进化树
3 讨论
本试验对14株H3亚型禽流感病毒的全基因进行了测序与序列分析,结果表明,HA基因在其裂解位点均为340PEKQTR↓GLF348,只有一个碱性氨基酸,能被特定蛋白酶识别,符合低致病性禽流感特征。14株分离毒株的HA基因氨基酸位点在226、228分别为Q、G而人源为226(L)、228(S),具有典型的禽源特征。有研究表明,HA基因上潜在的糖基化位点还能够影响AIV的毒力[14],对14分离株HA基因潜在的糖基化位点进行了分析,有部分毒株第24位氨基酸发生突变,导致24位(NST)糖基化位点消失,但是出现了新的糖基化位点22(NDS)。另外部分毒株NA基因糖基化位点也发生了增加现象,新出现的糖基化位点对AIV毒力的影响还需进一步试验验证。
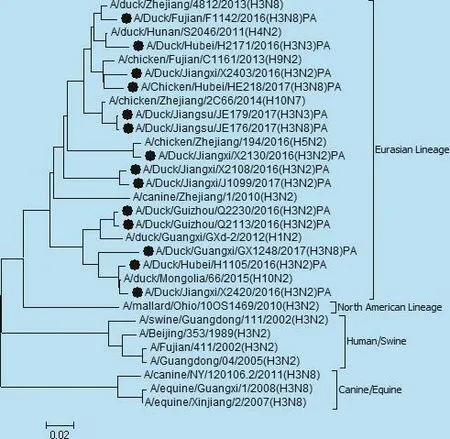
图5 PA基因遗传进化树
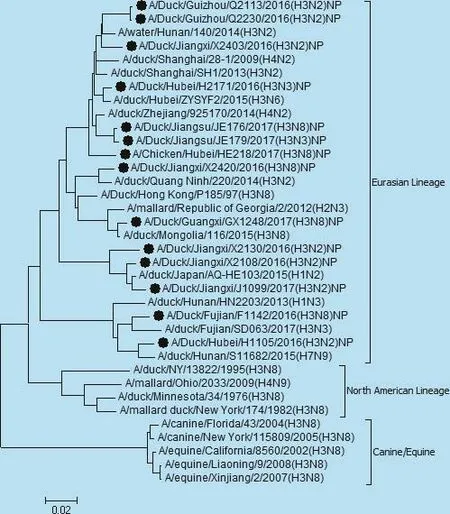
图6 NP基因遗传进化树
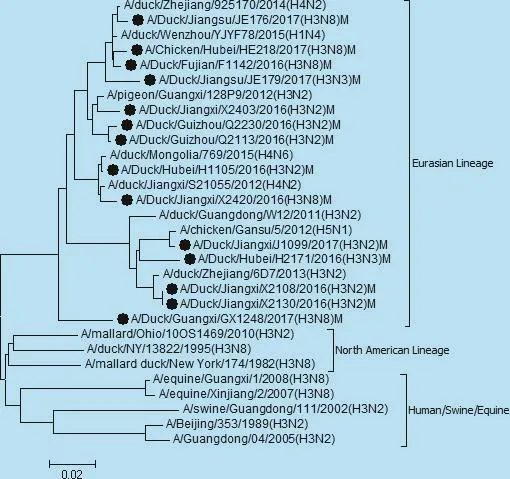
图7 M基因遗传进化树
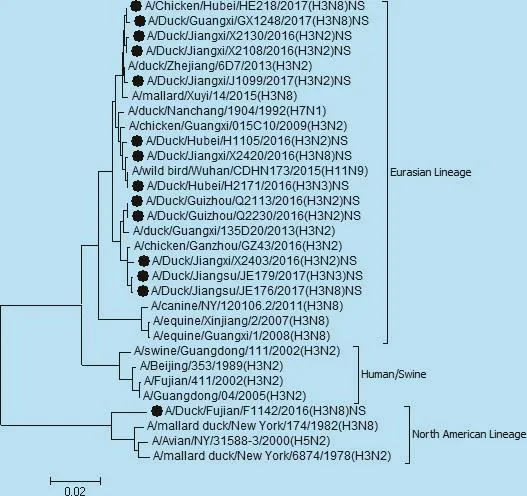
图8 NS基因遗传进化树
14株分离毒株来自我国2016年至2017部分省市活禽市场,其中除了NS基因的A/Duck/Fujian/F1142/2016(H3N8)毒株为北美谱系外其余各基因均为欧亚分支。观察进化树可知分离年份相近的的毒株,亲缘关系较近,各个基因核苷酸序列同源性高,可能来自相同的祖先。AIV容易通过抗原漂移或抗原转变发生变异,产生具有新的抗原特征的流感病毒。由于H3亚型 AIV致病力较低,在流行病学中的作用常被忽视,低致病性禽流感病毒感染家禽后虽然表现温和,但是存在的临床危险不容忽视,通过基因重排低致病性禽流感很有可能为形成高致病性禽流感病毒提供基因片段,因此必须加强对H3亚型禽流感病毒的监测。
