机械伤害处理橡胶树萌条树皮的DNA甲基化分析
2018-02-13吴绍华张世鑫田维敏
吴绍华,张世鑫,田维敏
(中国热带农业科学院 橡胶研究所, 农业部橡胶树生物学与遗传资源利用重点实验室, 省部共建国家重点实验室培育基地-海南省热带作物栽培生理学重点实验室, 海南儋州 571737)
巴西橡胶树(简称橡胶树),一种多年生热带乔木,是天然橡胶的主要来源。天然橡胶由于具有良好的弹性、绝缘性、耐磨性及抗高温等优良特性被广泛用作工业原材料。橡胶树的乳管是一种特化的细胞,分化于维管形成层细胞,成列分布[1-2],是天然橡胶合成和储存的场所[3]。在天然橡胶生产中,人们通常通过周期性切割橡胶树的树皮切断乳管(割胶)采集排出的胶乳提炼天然橡胶。割胶是橡胶树的一种周期性伤害,研究表明,割胶创伤会导致乳管的增加来增强植物保护伤口的潜力[4]。由于橡胶树树干树皮中次生乳管的数量与天然橡胶产量显著正相关,因此,开展乳管分化机理的研究对于指导割胶生产和改良橡胶树的产胶潜力具有重要的意义。由于橡胶树幼嫩萌条的第1~2伸长单位没有次生乳管[2,5],这有助于区分诱导的次生乳管。以橡胶树萌条为材料,研究证实机械伤害、茉莉酸、冠菌素及曲古菌素A均能诱导次生乳管的分化[2,4,6-7]。伤害部位脱水引起的活性氧爆发导致内源茉莉酸的积累是伤害诱导乳管分化的内因[5]。由于乳管的形成是一种极其复杂的细胞分化的过程,目前对乳管分化调控机理的认识仅停留在细胞形态学以及较浅显的分子生物学层面,对于其他层面的调控机理认识较少。DNA甲基化是一种保守的表观遗传修饰,在动植物基因调控及基因组稳定中起着重要的作用[8-10]。植物中的DNA甲基化主要发生在CG、CHG、CHH(H代表A, T, C)位点的胞嘧啶上[8,11],其中CG位点上的胞嘧啶甲基化在植物中占的丰度最高[12-14]。DNA甲基化与很多的生物进程有关,在调控植物的生长发育[10,15-17]、逆境胁迫[18-20]及细胞分化[21-23]中起着重要的作用。然而,对于表观遗传特别是DNA甲基化是否参与乳管分化的过程还未见报道。因此,本研究采用甲基化敏感扩增多态性(methylation-sensitive amplification polymorphism,MSAP)技术,探讨了伤害处理橡胶树萌条树皮的DNA甲基化的变化,为进一步从表观遗传学角度研究伤害诱导次生乳管的分化提供参考。
1 材料和方法
1.1 植物材料
巴西橡胶树无性系‘热研73397’植株,种植于海南省儋州市中国热带农业科学院橡胶研究所试验场。这些植株每年都被锯干,由茎干基部的潜伏芽长出萌条。萌条的顶芽在年生长周期中通常活动5~6次,这些叶簇中间隔一定长度的无叶茎秆。每一个这种明显的增长形态被称为伸长单位(extension unit,EU)。在自然条件下,从顶端往下数EU1和EU2是没有次生乳管的[2,5]。本研究取稳定期萌条EU1为材料进行处理。
1.2 实验方法
1.2.1机械伤害处理及材料的采集对稳定期萌条EU1的树皮,采用单面刀片轻轻刮去萌条树皮的上表皮后直接暴露于空气中。分别于伤害处理后暴露0.5、1、2、4、8、24、48、72 h后采集伤害部位的树皮,每个时间点采集3株混合为一个样品,液氮保存用于基因组DNA提取。不做任何处理的萌条树皮样品为对照。
1.2.2DNA甲基化敏感扩增多态性分析采用DNAsecure Plant Kit(天根,北京)提取树皮样品的基因组DNA。MSAP分析包含的酶切、连接、PCR扩增等参照Xiong等的方法[24]。采用2组限制性内切酶组合分别对基因组DNA进行双酶切,其中EcoR Ⅰ+HpaⅡ酶组合标记为EH,EcoR I+MspⅠ酶组合标记为EM。酶切后进行接头连接及PCR,接头及PCR扩增引物序列如表1,从中选择20对PCR引物进行MSAP分析。PCR产物变性后采用6%聚丙烯酰胺凝胶(PAGE)进行分离和银染。采用“+”和“-”统计PAGE胶条带,相同的位点有带记为“+”,无带记为“-”。
由于同裂酶HpaⅡ和MspⅠ对胞嘧啶甲基化的敏感性不同,根据酶组合EH和EM条带的有无,可将MSAP条带可分为4种类型(表 2):类型 I 条带为“+, +”,表明CCGG位点未甲基化;类型 Ⅱ 条带为“+, -”,表明 CCGG 位点发生单链外部甲基化(CHG 甲基化/半甲基化);类型 Ⅲ 条带为“-, +”, 表明 CCGG 位点发生双链内部甲基化(CG甲基化/全甲基化);类型 Ⅳ 条带为“-, -”,表明CCGG位点全甲基化[25-26]。甲基化水平采用以下公式进行计算:总甲基化率(%)=[(Ⅱ+Ⅲ+Ⅳ)/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100,全甲基化率(%)=[(Ⅲ+Ⅳ)/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100,半甲基化率(%)=[(Ⅱ)/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100。
1.2.3目的条带的回收、测序及序列分析从PAGE胶切下DNA甲基化差异位点条带,采用高盐法[27]回收目的片段,进行二次PCR后用pMD19-T Clone kit (TaKaRa)克隆差异片段后进行测序。用DNAMAN软件去除载体序列后,采用NCBI(http://www.ncbi.nlm.nih.gov/)网站的nucleotide Basic Local Alignment Search Tool (Blastn) 和translated DNA Blast (Blastx)工具进行序列的同源比对。
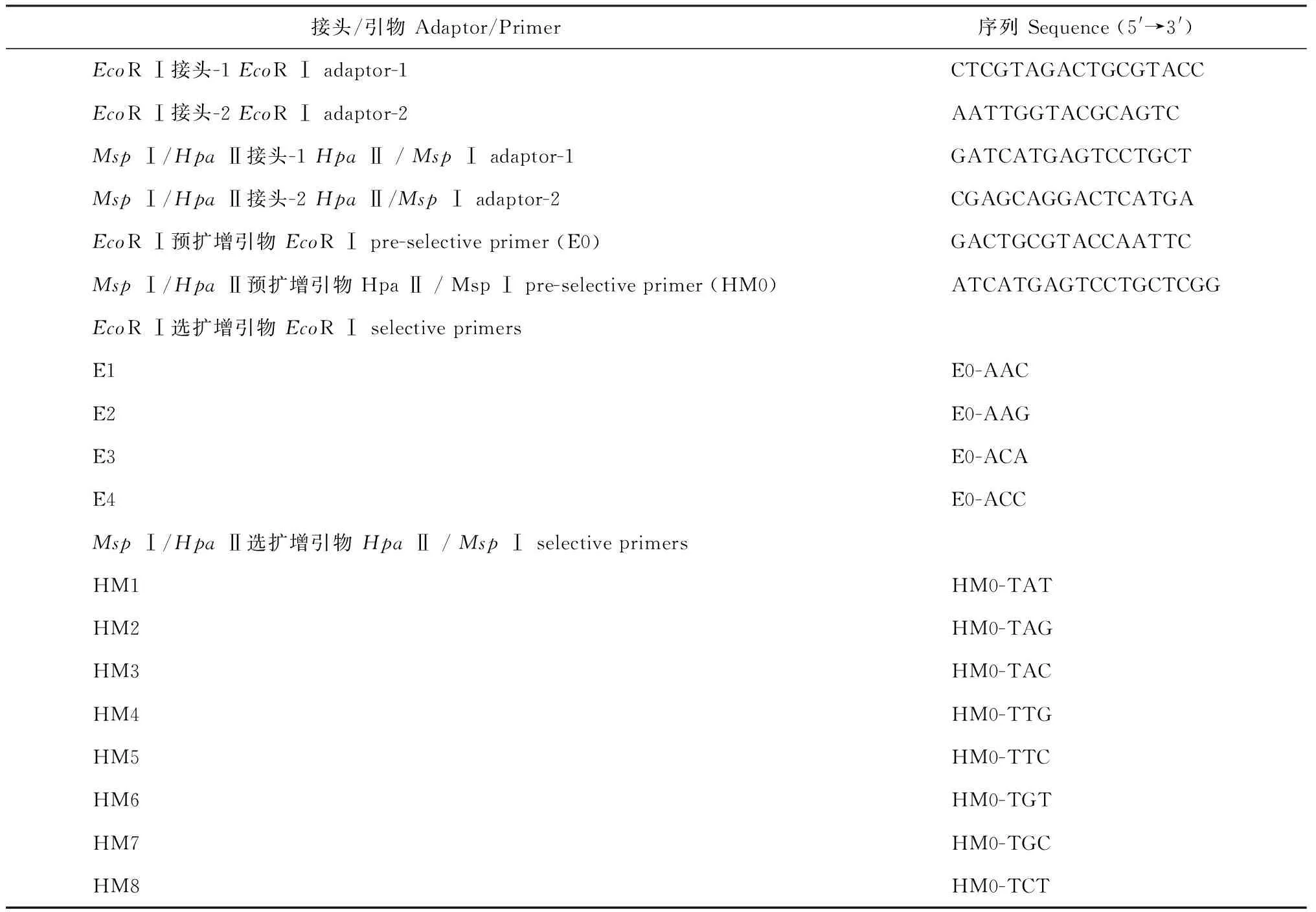
表1 用于MSAP分析的接头和引物序列
1.2.4重亚硫酸盐测序采用Epitect Plus DNA Bisulfite Kit (Qiagen, USA)试剂盒对差异位点对应的DNA样品进行重亚硫酸钠处理,处理后的DNA采用Premix TaqTM(TaKaRa, Japan)聚合酶进行甲基化特异性PCR扩增,引物采用MethPrimer[28]进行设计。编号15-1和45-2位点的扩增引物为15-1F(5′-ATTTTTAAATTATTGAGGGTTCG-AC-3′)和15-1R(5′-TTCTAATAAAATTATTAATTCCGCA-3′);45-2F(5′-TTCGTAGGATTTTAATT-CGTTTATTC-3′)和45-2R(5′-CTACTACCAACAATCTTTCTTCGAA-3′)。PCR产物送Invitrogen公司进行测序,并采用CyMATE[29]软件对胞嘧啶甲基化进行分析,通过对重亚硫酸钠处理样品与未处理样品的DNA序列进行分析,如胞嘧啶(cytosine,C)转换为胸腺嘧啶(thymine,T)表示胞嘧啶没有发生甲基化,如胞嘧啶无转换则表示胞嘧啶发生甲基化。
2 结果与分析
2.1 伤害处理橡胶树萌条树皮DNA胞嘧啶甲基化水平分析
本研究从32对选择性扩增引物组合(表1中E1~E4分别与HM1~HM8配对)中,筛选出条带多态性及清晰度较好的20对引物,分别为E1+HM5、E1+HM7、E2+HM1、E2+HM3、E2+HM4、E2+HM5、E2+HM6、E2+HM7、E2+HM8、E3+HM1、E3+HM2、E3+HM3、E3+HM4、E3+HM7、E3+HM8、E4+HM1、E4+HM2、E4+HM3、E4+HM5、E4+HM8,对萌条树皮基因组DNA的CCGG位点的胞嘧啶甲基化进行MSAP分析(图 1),共检测到了688个位点(表2)。在对照样品中,即自然状态下萌条树皮的甲基化水平为29.51%,其中全甲基化(类型Ⅲ+Ⅳ)占的比重较大,为27.76%。而伤害后暴露不同时间的样品,其甲基化水平变化范围为26.16%~29.94%(表2),其中伤害处理0.5、2和8 h这3个时间点的甲基化水平比对照略有升高,升高幅度较低,只有0.4%左右。而伤害处理1、4、24、48和72 h的甲基化水平比对照有所下降,其中甲基化水平下降幅度最大是发生在伤害处理48 h后,下降幅度达到了3.35%。与对照相似,伤害处理样品的全甲基化(类型Ⅲ+Ⅳ)占甲基化水平的比重也较大(表2)。而相比对照的半甲基化比率(1.74%),只有48(1.60%)和72 h(1.31%)样品的半甲基化位点比率有较低幅度的降低。因此,甲基化水平的变化主要是由于全甲基化(类型Ⅲ+Ⅳ)位点的改变。
2.2 伤害处理橡胶树萌条树皮DNA胞嘧啶甲基化变化类型的分析
与对照相比,当PAGE胶上同一位置的条带由于伤害处理发生了变化,说明此位点的甲基化模式发生了改变。为了阐明伤害处理导致橡胶树萌条树皮DNA甲基化变化的程度和类型,对PAGE条带的统计和分析结果(表3)显示,伤害处理与对照相比,存在13种DNA甲基化类型,依次将其命名为类型A~M(表3)。这些甲基化类型可以归纳为3大类:DNA甲基化无变化、去甲基化和甲基化。其中DNA甲基化无变化包含A~D 4种类型,对照和伤害处理的DNA甲基化是一致的;E~I 5种类型代表的是胞嘧啶去甲基化,即伤害处理导致CCGG位点发生了去甲基化;J~M 4种类型代表的是胞嘧啶的甲基化,即伤害处理导致CCGG位点发生了甲基化。与对照相比,超过94% CCGG位点在伤害处理时甲基化保持不变。伤害处理0.5 h、1 h、2 h、4 h、8 h、24 h、48 h、72 h后,去甲基化发生的概率分别为0.14%、1.89%、0.29%、1.31%、0.29%、2.18%、4.22%、1.31%;甲基化发生的概率分别为0.73%、0.44%、3.78%、0.87%、0.87%、0.87%、1.02%、0.73% (表 3)。去甲基化事件发生频率最高的是在伤害处理48 h(4.22%),而甲基化事件则主要发生在伤害处理2 h后(3.78%)。这与甲基化水平的变化趋势是一致的,与对照的甲基化水平29.51%相比,伤害处理2 h的甲基化水平升高至29.94%,伤害处理48 h的甲基化水平下降至26.16%。
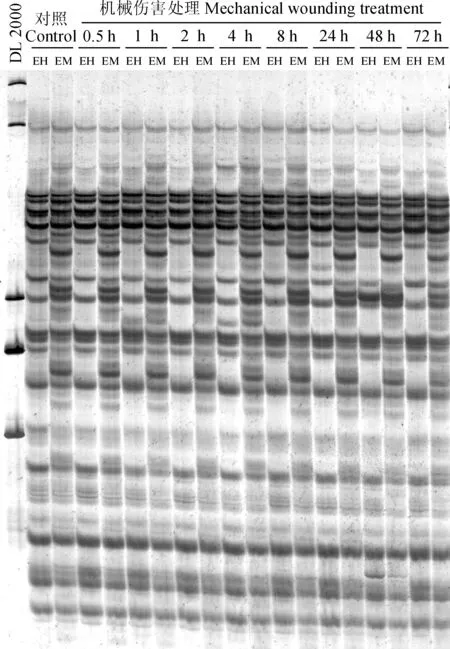
图1 MSAP分析胶图Fig.1 The PAGE of MSAP analysis
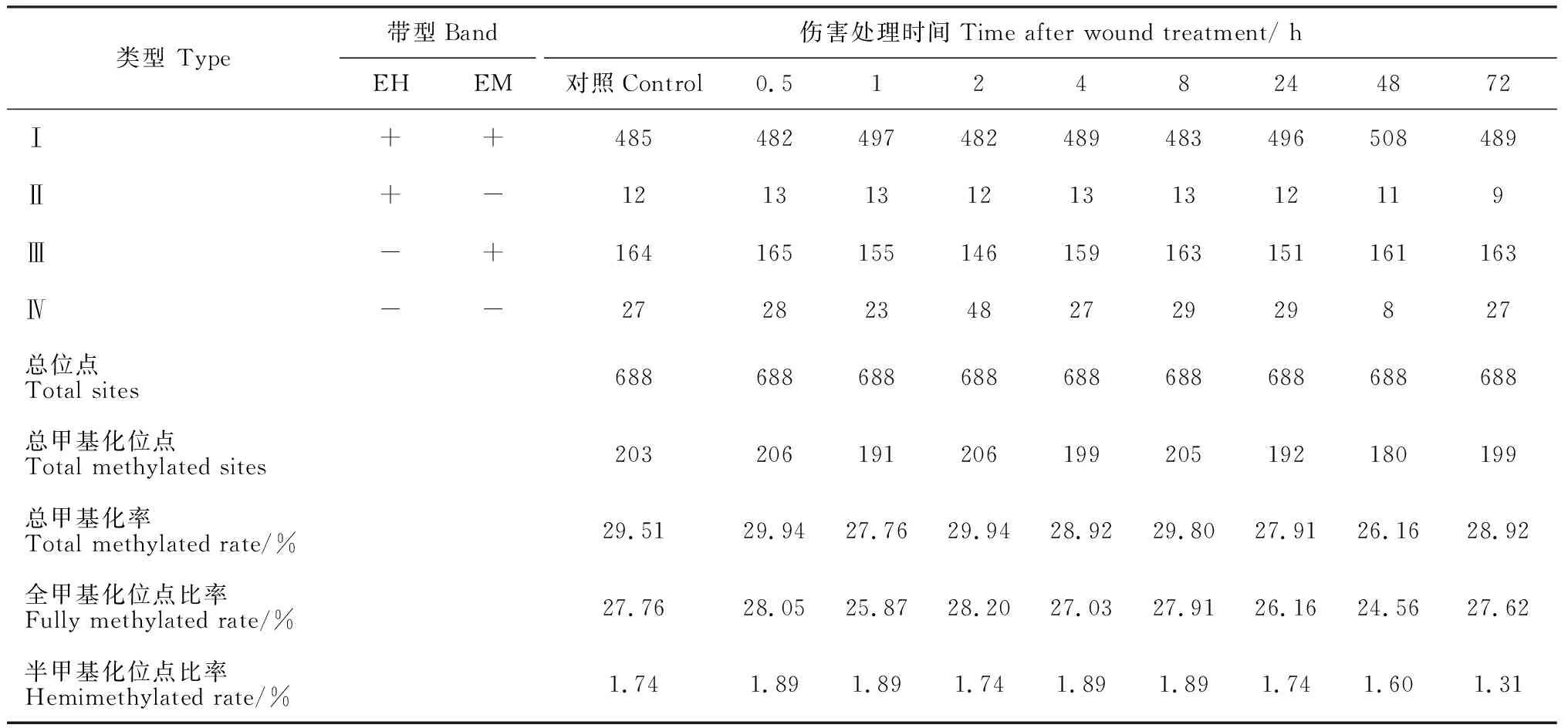
类型 Type带型Band伤害处理时间Time after wound treatment/ hEHEM对照Control0.5 1 24 8 24 48 72 Ⅰ++485482497482489483496508489Ⅱ+-12131312131312119Ⅲ-+164165155146159163151161163Ⅳ--27282348272929827总位点Total sites688688688688688688688688688总甲基化位点Total methylated sites203206191206199205192180199总甲基化率Total methylated rate/%29.5129.9427.7629.9428.9229.8027.9126.1628.92全甲基化位点比率Fully methylated rate/%27.7628.0525.8728.2027.0327.9126.1624.5627.62半甲基化位点比率Hemimethylated rate/%1.741.891.891.741.891.891.741.601.31
注:+表示有条带,-表示无条带;下同
Note: + represented the bands presented, - represented no bands; The same as below
2.3 差异DNA甲基化片段的分析及重亚硫酸盐测序验证
通过比较对照和伤害处理的甲基化状况,回收PAGE胶相应的甲基化差异位点,鉴定了20条DNA甲基化差异片段序列,片段的长度范围为102~459 bp,其中有8个片段与橡胶树‘热研73397’的基因组序列匹配[30],这些序列的DNA甲基化变化主要发生在基因的外显子及UTR区(表 4)。比对分析显示,这些片段匹配的基因注释为ATP 合酶F1亚基1、磷酸核糖胺-甘氨酸类连接酶、冷激结构域蛋白3、光系统Ⅱ 47 kDa 蛋白、E3泛素蛋白连接酶RING1、NADH泛醌氧化还原酶和一些假定蛋白(表 4)。其中,采用重亚硫酸盐测序对ATP 合酶F1亚基1(15-1)和NADH泛醌氧化还原酶(45-2)的甲基化进行了验证。重亚硫酸盐测序结果表明,与对照相比,这2个CCGG位点受伤害处理后均发生了去甲基化,与PAGE胶显示的结果一致(图 2)。
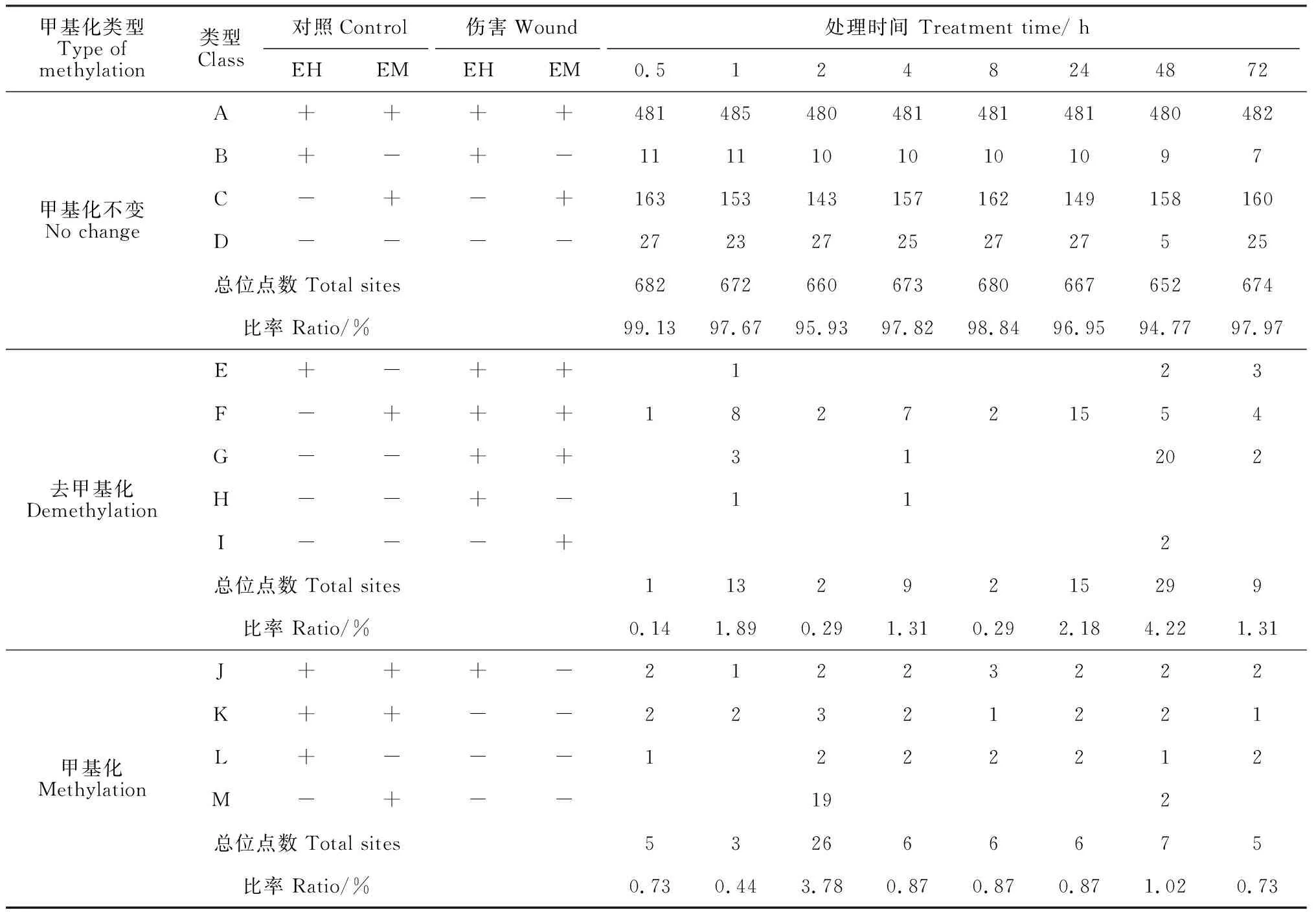
表3 伤害处理橡胶树萌条树皮DNA甲基化与对照比较
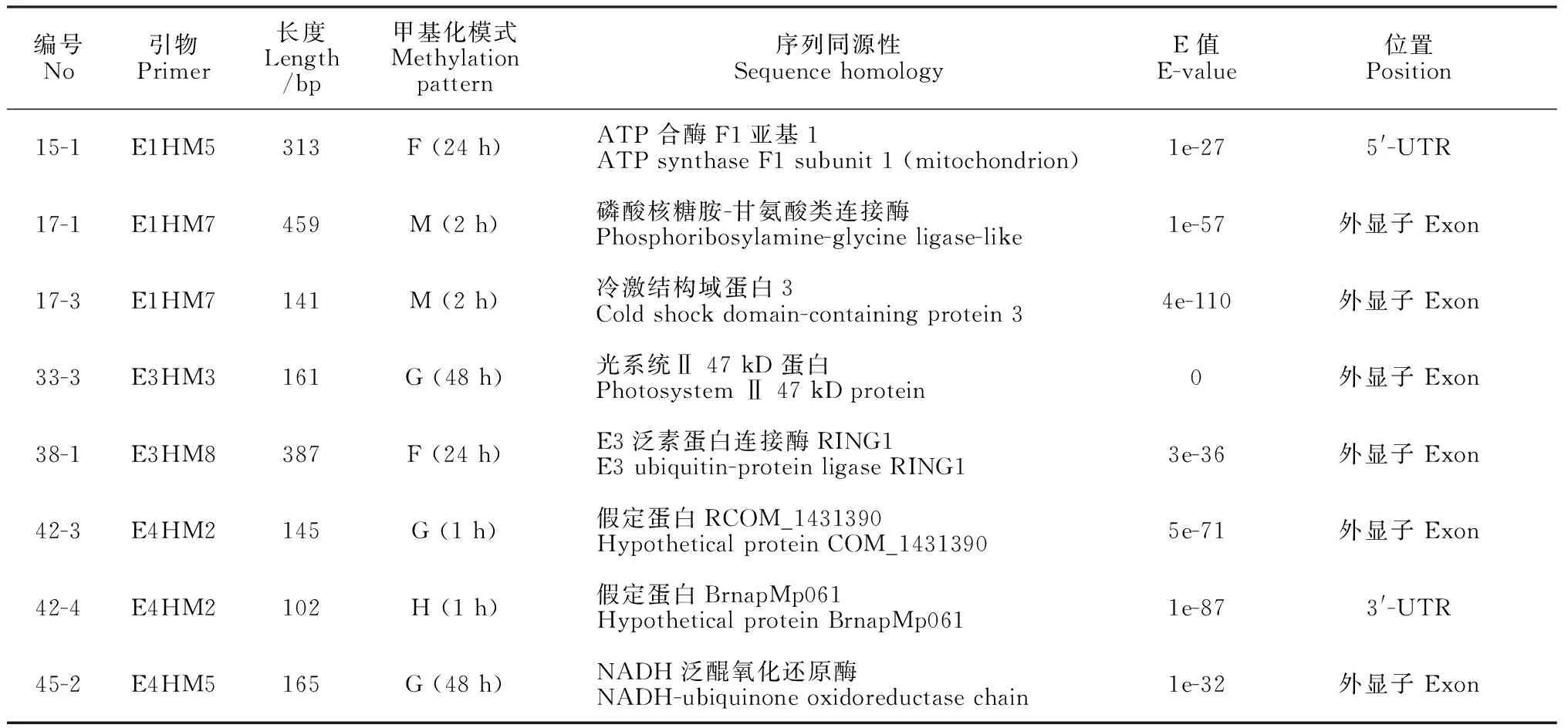
表4 DNA甲基化差异位点的匹配基因的功能注释
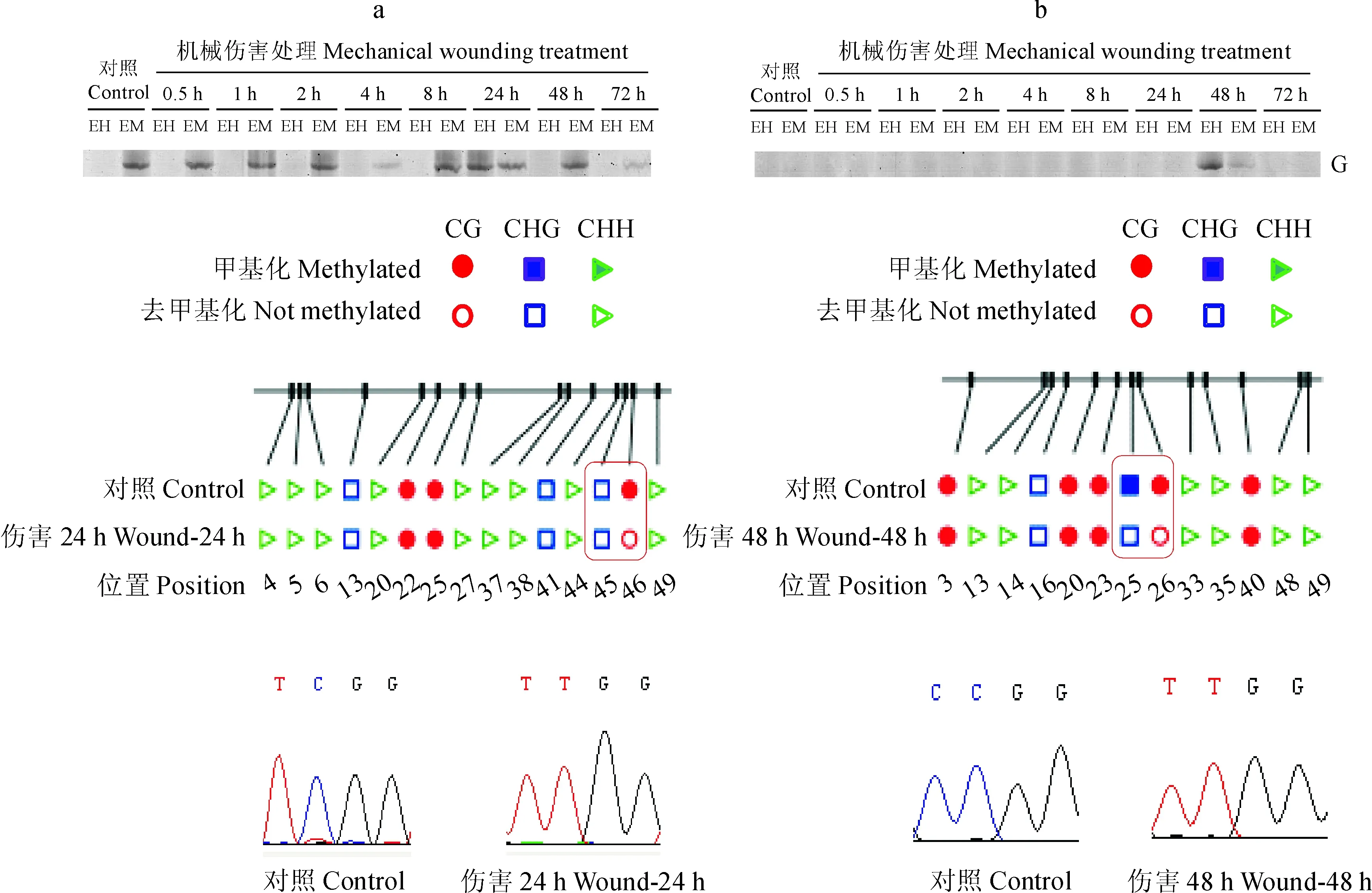
图2 15-1 (a) 和 45-2 (b)甲基化差异位点重亚硫酸盐测序结果Fig.2 The bisulfite sequencing analysis of the CCGG sites in 15-1 (a) and 45-2 (b)
3 讨 论
胞嘧啶甲基化是一种主要的甲基化修饰,与转录沉默具有密切的相关性[31]。MSAP技术常用来检测胞嘧啶甲基化的改变和分离基因组中随机分布的甲基化位点[24,32-33]。本研究采用MSAP技术首次分析了伤害处理橡胶树萌条树皮的甲基化变化及模式。机械伤害处理萌条1 h导致甲基化水平的下降,2 h后甲基化水平恢复至对照水平并有所升高,这种变化趋势与玉米叶片的受伤害后DNA甲基化的变化相似:伤害1 h导致玉米的DNA甲基化水平瞬时下降至20%~30%,随后恢复至原先的水平[34]。除此之外,机械伤害处理橡胶树萌条树皮还有一个效应,就是伤害暴露能诱导萌条次生乳管的产生。在本研究中,伤害处理橡胶树萌条24 h和48 h,甲基化水平分别降至27.91%和26.16%。而且,甲基化变化模式分析表明,超过94%的甲基化位点在伤害处理后没有发生变化。甲基化事件主要发生在2 h(3.78%),去甲基化事件主要发生在48 h(4.22%)。
另外,对8个匹配基因组的DNA甲基化差异位点的功能注释显示,伤害导致6个位点发生去甲基化,分别是ATP合酶F1亚基1、光系统Ⅱ 47 kD 蛋白、E3泛素蛋白连接酶RING1、NADH泛醌氧化还原酶和2个假定蛋白;伤害导致位点发生甲基化的有磷酸核糖胺-甘氨酸类连接酶和冷激结构域蛋白。ATP合酶F1亚基1和NADH泛醌氧化还原酶是电子传递链中组分,与活性氧的产生或消除相关。E3泛素蛋白连接酶主要包含3大类:HECT结构域家族、RING结构域家族及U-box蛋白家族。有研究表明,RING型E3泛素连接酶通过介导植物的多个信号途径参与植物的生长发育及逆境响应[35]。橡胶树伤害诱导的次生乳管分化主要是伤害部位暴露脱水引起的内源茉莉酸大量积累有关[5]。因此,伤害导致了相关基因发生了甲基化或去甲基化,是否会参与次生乳管的发育?这一推断有待于进一步的深入研究。
