补骨脂酚对UVB诱导HaCaT细胞凋亡因子p53和caspase—3表达的影响
2018-02-02孙琪朴成玉陈丹丹等
孙琪+朴成玉+陈丹丹等

[摘要]目的:探讨补骨脂酚对UVB诱导HaCaT细胞凋亡因子p 53和caspase-3表达的影响及其机制。方法:四甲基噻唑蓝[3-(4,5-Dimethyl-2-Thiazolyl)-2,5-Diphenyl Tetrazolium Bromide,MTT]法检测不同浓度补骨脂酚对正常细胞增殖的影响。实验设计分为四组,空白组、模型组、雌二醇组及补骨脂酚组。以照射强度0.61mW/cm2,照射时间10min,建立HaCaT细胞凋亡模型;MTT法检测各组细胞增值率;活性氧(Reactive Oxygen Species,ROS)试剂盒检测各组细胞中ROS含量变化;线粒体膜电位检测试剂盒(JC-1)检测各组细胞线粒体膜电位变化;实时定量PCR(Reverse transcriptional PCR,RT-PCR)检测各组细胞中caspase-3 mRNA表达水平;蛋白免疫印迹法(Western Blot)检测各组细胞中p53、caspase-3蛋白表达水平。结果:10-6mol/L补骨脂酚组细胞增值率无明显变化(P>0.05),但是可降低ROS含量、线粒体膜电位、caspase-3mRNA表达量、p53及caspase-3蛋白表达量(P<0.05)。结论:补骨脂酚可抑制UVB诱导的HaCaT细胞凋亡,其作用机制主要是通過降低细胞内活性氧的含量、降低细胞线粒体膜电位、抑制p53及caspase-3的表达。
[关键词]补骨脂酚;UVB;HaCaT;凋亡;p53;caspase-3
[中图分类号]R285.5 [文献标志码]A [文章编号]1008-6455(2017)07-0037-04
紫外线包括UVA(长波)、UVB(中波)及UVC(短波)三种不同波长。其中UVB虽然波长短,但是辐射强度大,可以穿透表皮,因此UVB越来越引起人们的关注。紫外线照射剂量超过皮肤承受能力,会引发皮肤炎症、老化、凋亡甚至皮肤癌。豆科植物补骨脂干燥成熟种子中除了含有补骨脂素及异补骨脂素,还含有少量的补骨脂酚。补骨脂酚是补骨脂挥发油的主要成分,约占60%。研究表明,补骨脂酚具有抗炎、抗氧化、抗菌、抗病毒、抗肿瘤、抗抑郁以及雌激素样作用。近几年来,补骨脂酚的研究越来越多,有报道称,补骨脂酚对成纤维光老化衰老基因具有调控作用,与本实验研究结果相近。本研究选择UVB诱导HaCaT细胞凋亡模型作为研究对象,观察补骨脂酚对UVB诱导HaCaT细胞凋亡因子p53、caspase-3 mRNh及蛋白表达的影响。
1材料和方法
1.1实验材料:细胞株为人永生化角质形成细胞,来源于上海中乔新舟有限公司。
主要仪器:SS-01B型UVB紫外光疗仪(上海希格玛高技术公司);MK3型酶标仪(上海热电仪器有限公司);SmartChemi500型一体式微型化学发光成像仪(北京赛智创业有限公司)。
试剂:补骨脂酚标准品(成都曼思特生物科技有限公司,批号:MUST-15012210);雌二醇标准品(中国药品生物制品检定所);DMEM培养液(Hyclone公司,批号:NZM1301);胎牛血清(FBS)(Hyclone公司,批号:NYB0614);四甲基噻唑蓝(MTT)(Sigma公司,批号:021005);线粒体膜电位检测试剂盒(JC-1)(碧云天生物技术);小鼠抗人β-actin单克隆抗体(北京中杉金桥有限责任公司);兔抗人p53多克隆抗体(南京恩晶生物科技有限公司);兔抗人caspase-3多克隆抗体(博奥森生物有限公司);羊抗小鼠二抗(博士德生物有限公司);羊抗兔二抗(博士德生物有限公司)。
1.2实验方法
1.2.1溶液配制:雌二醇溶液配制:将0.23mg雌二醇粉末放入到2ml无水乙醇中进行混合,加入吐温250μl,再使用DMEM培养液进行10倍稀释,最终浓度达到1×10-7mol/L,最后0.22Pm微孔滤膜过滤除菌,-20℃冰箱保存待用。
补骨脂酚溶液配置:精密称取补骨脂酚2.54mg,用10μl二甲基亚砜(Dimethylsulfoxide,DMS0)将其充分溶解,再加I)MEM基础培养基定容至10ml,浓度为10-3mol/L,0.22μm滤膜滤过除去细菌及杂质,DMEM培养液稀释补骨脂酚,稀释浓度分别为10 4mol/L、10-5mol/L、10-6mol/L、10-7mol/L、10-8mol/L、10-9mol/L、10-10mol/L、10-11lmol/L等9个浓度,-20℃冰箱保存待用。
JC-1染色工作液配置:将8ml纯水加入到50μl JC-1(200×)中进行混匀,再加入2ml JC-1染色缓冲液(5×)混匀即可。
JC-1染色缓冲液(1×)配置:取3ml JC-1染色工作液(5×)加入12ml纯水,混匀,放于冰浴中保存待用。
1.2.2 HaCaT细胞凋亡模型的建立及分组:参照文献,按每孔15×103个细胞接种于96孔板,将其分为空白组、模型组、10-7mol/L雌二醇及10-6mol/L补骨脂酚组。培养24h后,空白组及模型组分别加入新的培养液继续培养;10-7mol/L雌二醇及10-6mol/L补骨脂酚组分别加入相应的药物溶液培养。24h后,空白组更换新的培养液,使用铝箔纸盖上继续培养24h;模型组、10-7mol/L雌二醇及10-6mol/L补骨脂酚组弃去培养液,分别加入200μl磷酸盐缓冲液(Phosphate Buffered Solution,PBS)覆盖,选择最佳照射时间10min进行照射,弃去PBS后继续加入新的培养液继续培养24h。最终,加入20μl MTT及150μl DMSO,酶标仪检测各组细胞吸光值。endprint
1.2.3细胞增长率测定:参照文献,取对数生长期细胞,接种于96孔板,培养24h,根据预实验结果,选定10-11、10-10、10-9、10-8、10、1-7、10-6、10-4及10-3mol/L九个浓度补骨脂酚加入96孔板中,继续培养24h,弃去旧培养液,继续加入新培养液培养24h,再加入20μl MTT及150μl DMSO,于492nm波长处酶标仪检测每组间细胞的吸光值。细胞增殖率=给药组OC值/空白组OD值(其值设为1)×100%。
1.2.4 ROS试剂盒检测ROS含量:取对数生长期的细胞,消化、离心,接种于6孔板后,进行分组,分别为空白组、模型组、10-7mol/L雌二醇及10-6mol/L补骨脂酚组,于37℃5%CO2培养箱中培养,72h后取出,弃掉培养液,加入1mlDCFH-DA(10μM/L)溶液,放于培养箱孵育20min,每隔5min颠倒混匀一下,使探针和细胞充分接触。采用无血清的培养液洗涤3次,PBS清洗3次,加入胰蛋白酶进行消化,培养液终止消化,离心,放于1.5ml离心管中,加入1mlPBS重悬细胞,滤网滤过。最后,使用流式细胞仪检测各组细胞中ROS含量。
1.2.5线粒体膜电位试剂盒检测各组细胞膜电位变化:建模成功后,收集每组细胞分别放于1.5ml离心管中,参照文献,每管中分别加入0.5ml培养液及0.5ml JC-1染色工作液混匀,37℃培养箱中孵育20min,加入1ml JC-1染色缓冲液(1×)重悬,6009 4℃离心5min,弃上清,重復2次,再加入1ml JC-1染色缓冲液(1×)重悬,流式细胞仪进行检测。
1.2.6 RT-PCR测定各组细胞中mRNA表达水平:建模成功后,Trizol试剂、氯仿、异丙醇、75%乙醇提取各组RNA,使用Nano-100微量分光光度计测定RNA纯度和浓度,计算所需体积数,按照逆转录试剂盒说明书将RNA逆转录为eDNA,然后以cDNA为模板进行实时定量PCR。caspase-3引物(F:5-TGGTTCATCCAGTCECl3TG-3,R:5-ATTCTGTTGCCAC-TITTCG-5)。
反应程序:预变性95℃ 5min,变性95℃ 30s,退火/延伸60℃ 60s,30个循环。本实验的循环数即Ct值、融解曲线和扩增曲线可从real-time PCR上直接读取。β-actin作为各处理组的内参基因,caspase-3等目的基因在处理组相对于对照组的AACt值计算得出:AACt=(Ct目的基因Ct内参基因)处理组-(Ct目的基因-Ct内参基因)对照组,按照公式可算出各组caspase-3等目的基因相对于对照组表达量:目的基因相对表达量=2-ΔΔCt。
1.2.7 Western Blot检测各组细胞中蛋白表达水平:建模成功后,按300μl/瓶加入RIPA裂解液,使用前2~3min需向裂解液内加入3μl PMSF(终浓度1mM),选用BCA试剂盒测蛋白浓度,电泳75min,转膜35min,封闭液封闭2h,一抗(1:300)孵育,于4℃条件下过夜。次日,二抗(1:10 000)孵育1h,加入TBST后放于床上震荡,重复3次,在膜上滴加发光工作液,把膜置于化学发光成像仪内,曝光并拍照,分析图像结果,应用Lane ID凝胶分析系统分析条带灰度(IOD),分析并获得各条带的灰度值,目的蛋白的表达水平根据靶条带的灰度值与内参(β-actin)的灰度值的比值来反映。
1.3统计学分析:采用SPSS23.0软件进行分析,计量资料均用(x±s)表示,将用单因素方差分析(one-wayANOVA)进行分析,经组间比较方法,两两比较采用LSD法。P<0.05认为差异具有统计学意义。
2结果
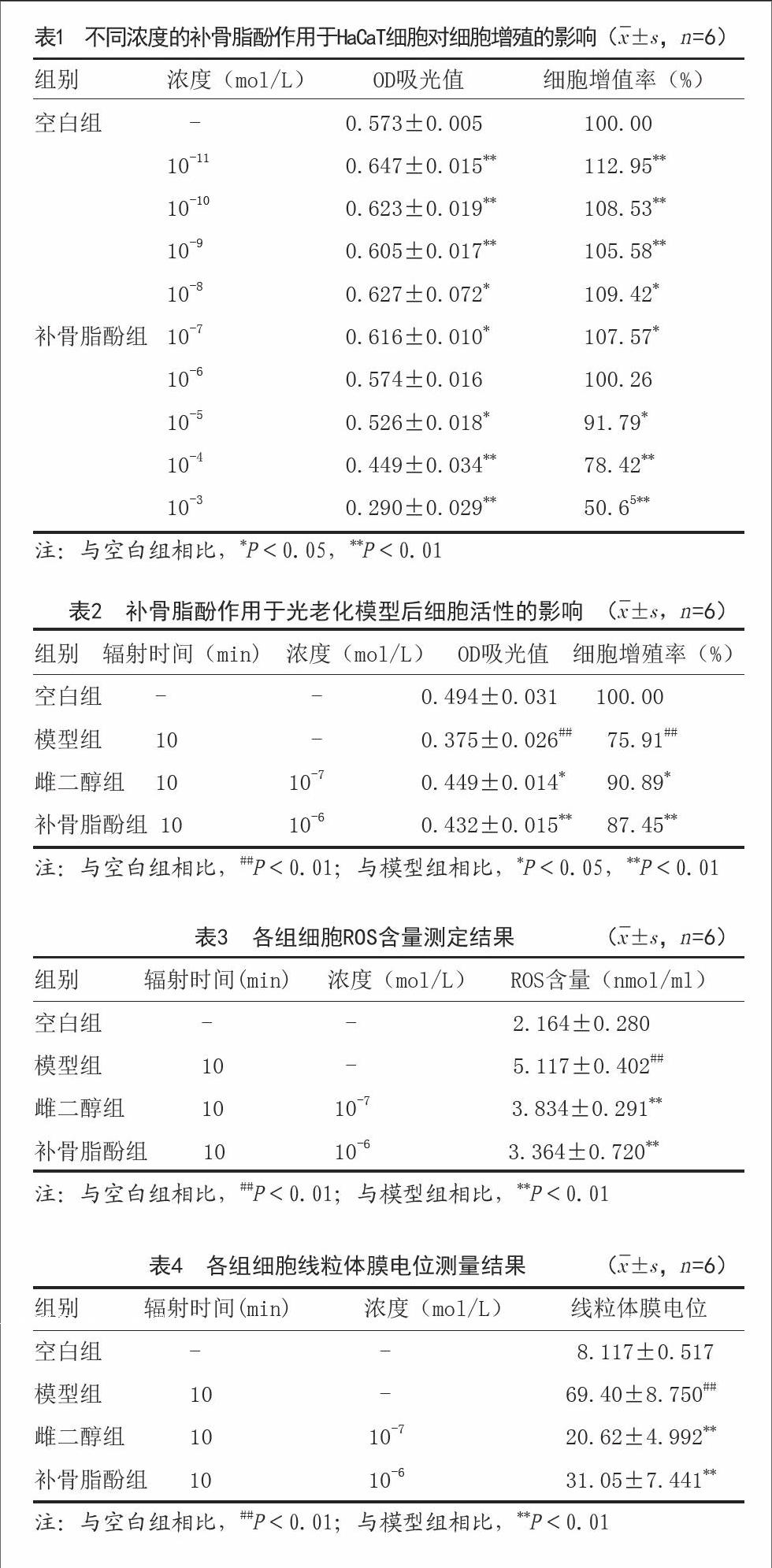
2.1不同浓度补骨脂酚对细胞增殖率的影响:如表1所示,与空白组相比,10-11、10-10、10-9、10-8、10-7mol/L补骨脂酚组细胞增殖率明显上升(P<0.01),10-5、10-4、10-3mol/L补骨脂酚组细胞增殖率明显降低(P<0.05),10-6mol/L补骨脂酚组细胞增值率无明显变化,差异无统计学意义(P>0.05)。
2.2补骨脂酚作用于光老化模型对细胞活性的影响:如表2所示,与空白组相比,模型组细胞增殖率明显下降(P<0.01);与模型组相比,10-7mol/L雌二醇组及10-6mol/L补骨脂酚组细胞增殖率明显上升,差异具有统计学意义(P<0.05)。
2.3 ROS含量测定结果:如表3所示,与空白组相比,模型组ROS含量明显升高(P<0.01);与模型组相比,10-7mol/L雌二醇组及10-6mol/L补骨脂酚组ROS含量明显降低,差异有统计学意义(P<0.05)。
2.4线粒体膜电位测量结果:如表4所示,与空白组相比,模型组线粒体膜电位明显增加(P<0.01);与模型组相比,10-7mol/L雌二醇组及10-6mol/L补骨脂酚组线粒体膜电位明显降低,差异具有统计学意义(P<0.05)。
2.5各组细胞中caspase-3 mRNA表达量测定结果:如表5所示,与空白组相比,模型组中easpase-3 mRNA表达量升高(P<0.01);与模型组相比,10-7mol/L雌二醇组及10-6mol/L补骨脂酚组caspase-3 mRNA表达量均降低,差异具有统计学意义(P<0.01)。
2.6各组细胞中p53、easpase-3蛋白表达量测定结果:如表6及图1所示,与空白组相比,模型组中P53及caspase-3蛋白表达量明显上升(P<0.01);与模型组相比,10-7mol/L雌二醇组及10-6mol/L补骨脂酚组中p53及easpase-3蛋白表达量明显下降,差异具有统计学意义(P<0.01)。
3讨论
皮肤易遭受紫外线辐射诱发细胞凋亡。因角质形成细胞位于皮肤表皮层,紫外线主要诱导角质形成细胞凋亡。其中p53可以被UVB诱导的DNA损伤所激活,细胞凋亡主要是由多个信号通路调控的死亡过程,p53蛋白在细胞的凋亡过程中占有重要的作用。p53蛋白和半胱氨酸盐酸天冬氨酸蛋白-3(caspase-3)是促凋亡基因,在DNA损伤、细胞凋亡、癌基因的激活等方面发挥重要作用。caspase-3蛋白酶作为细胞凋亡的终极指标,是参与细胞凋亡的关键酶,其中,caspase-3可以促使各种凋亡因子被激活,一旦被激活,可以使细胞内蛋白质降解,最终导致细胞死亡。p53参与调控线粒体凋亡通路,主要是通过对线粒体膜上Bcl-2家族蛋白成员的转录调控,使线粒体膜电位发生改变,最终导致激活caspases-3以及启动细胞凋亡级联反应。
为了阐明补骨脂酚对HacaT细胞凋亡保护作用的机制,本研究继续观察补骨脂酚对凋亡相关基因(p53、caspase-3)表达的影响,结果显示,与空白对照组相比,模型组凋亡相关基因p53、caspase-3表达水平显著升高,该结果与模型组HaCaT细胞凋亡水平显著升高相一致。而给予10-7mol/L雌二醇及10-6mol/L补骨脂酚治疗后,与模型组相比,治疗组HaCaT细胞凋亡水平显著降低。
综上所述,本研究结果证实补骨脂酚对HaCaT细胞凋亡保护作用的机制可能是通过调节p53及caspase-3相关基因的表达。通过本实验研究发现,可以将补骨脂酚应用于天然化妆品中,为预防紫外线对皮肤的破坏奠定一定的理论基础。endprint
