腺苷受体A1亚型对视网膜色素上皮细胞免疫调节功能的影响
2018-02-02孔繁强周树民张伟陈松
孔繁强,周树民,张伟,陈松
腺苷(adenosine)为三磷酸腺苷(ATP)的代谢产物,部分研究认为其对机体有免疫调节作用[1-2]。细胞损伤或缺氧时会大量释放ATP到细胞外,促进局部炎症反应的进展[3];位于细胞膜上的2种磷酸酶CD39和CD73可迅速转化ATP为腺苷[4-5]。腺苷与其受体(adenosine receptors,ARs)相结合,可发挥局部免疫调节作用[6-7]。笔者此前研究显示,视网膜色素上皮(retinal pigment epithelium,RPE)细胞是眼底组织中唯一高表达CD73的细胞,提示RPE在产生和利用腺苷上具有重要的作用[8]。ARs系统由ARA1、ARA2A、ARA2B及ARA3这4种亚型组成,它们在不同组织、细胞中的表达、分布及对腺苷的亲和力显著不同[9-10]。而这些不同亚型的ARs在功能上也截然不同。因此,了解不同种类的细胞是通过何种AR来利用腺苷是研究腺苷局部免疫调节作用的前提。本文旨在探讨ARPE-19细胞中高亲和力的腺苷受体亚型及其潜在功能。
1 资料与方法
1.1 一般资料 人ARPE-19细胞购自中国医学科学院细胞库;RNA提取试剂Trizol、人单核细胞趋化因子1(MCP-1)及C-X-C配体 10(CXCL10/IP-10)酶联免疫吸附试验(enzyme linked immune sorbent assay,ELISA)试剂盒购自Invitrogen公司;白细胞介素(IL)-6、IL-10、转化生长因子 β(TGF-β)的ELISA试剂盒购自R&D公司;逆转录试剂及定量PCR试剂盒购自大连宝生物公司;小鼠抗人ARA1、ARA2A、ARA2B、ARA3单克隆抗体购自Santa Cruz公司;HRP标记的兔抗小鼠IgG购自武汉博士德公司;放射性H3标记的腺苷(H3-adenosine)购自北京原子能研究所;ARA1拮抗剂DPCPX、ARA2A拮抗剂SCH58261、ARA2B拮抗剂MRS1754、ARA3拮抗剂MRS1220及ARA1激动剂CCPA购自美国Tocris公司。
1.2 腺苷受体在RPE中的表达 体外培养ARPE-19细胞于6孔细胞培养板中,至80%融合后取3孔细胞充分洗涤,加入Trizol试剂(1 mL/孔),室温静置5 min后刮除细胞,置于Trizol试剂中,参照试剂说明书进行RNA提取。取1.0µg总RNA逆转录合成cDNA;取少量cDNA,real-time PCR测定4种腺苷受体基因的表达以gapdh为内参照。上、下游引物序列见表1。另3孔细胞参照文献[11]用于膜蛋白提取及Western blot检测:取适量膜蛋白(1µg/样本)行SDS-PAGE分离,半干法转移凝胶中的蛋白条带至硝酸纤维素膜(NC膜)上。剪取单个样本的NC膜,经5%脱脂奶粉溶液封闭后,分别用抗ARA1、ARA2A、ARA2B及ARA3的抗体孵育,4℃过夜。充分洗涤后置于含HRP-兔抗小鼠IgG抗体的溶液中,室温放置1 h。充分洗涤后将NC膜置于ECL发光底物中,室温放置10 min,于暗盒中对X-ray胶片曝光、显影及定影。

Tab.1 Primers used for real-time PCR表1 定量PCR引物序列
1.3 放射性配体结合实验 体外培养ARPE-19细胞至80%融合,按1×105个/mL密度接种至96孔细胞培养板。待细胞充分贴壁后加入不同剂量(0、5、10、30、100、200、400、800及 1 200 nmol/孔)H3-adenosine,每个剂量设立 3个复孔。37℃温育1 h,抽吸转移细胞至GF/C膜。先后以去离子水及95%乙醇充分洗涤、晾干后加入闪烁计数液,以液体闪烁计数器计数放射强度后计算出细胞所结合的腺苷量,绘制H3-adenosine结合曲线;Scat chart作图计算出RPE对腺苷的最大结合容量(Bmax),以单位数量细胞(1×104细胞)理论上所能结合的最多腺苷量(单位:fmol)来表示。
1.4 不同亚型腺苷受体对RPE结合腺苷的贡献 将ARPE-19细胞接种至96孔细胞培养板,平均分为5组(A~E),每组含27孔细胞,分别给予1.3中所示的9种H3-adenosine浓度,每个浓度设3个复孔。其中,A组为对照组;B~E组所有细胞分别给予ARA1拮抗剂DPCPX(终浓度50 nmol/L)、ARA2A 拮抗剂 SCH58261(100 nmol/L)、ARA2B 拮抗剂 MRS1754(100 nmol/L)及 ARA3 拮抗剂 MRS1220(5µmol/L),37℃温育30 min。此后,按照1.3中所述方式计算每组细胞结合腺苷的Bmax(n=3)。
1.5 ARA1信号通路对ARPE-19分泌细胞因子及趋化因子的影响 体外培养ARPE-19细胞至80%融合,随机分为对照RPE组(不予ARA1激动剂)及CCPA干预RPE组(CCPA 100 nmol/L),每组6孔细胞。2组细胞均给予TNF-α 10µg/L及IFN-γ 1 000 U/mL联合干预。48 h后收集上清,ELISA法检测 IL-6、IL-10、TGF-β、MCP-1、IP-10 含量。
1.6 统计学方法 采用SPSS 17.0统计软件进行统计处理。符合正态分布的计量数据以均数±标准差(±s)表示。2 组比较采用t检验,多组比较采用方差分析,组间多重比较用LSD-t检验,以P<0.05为差异有统计学意义。
2 结果
2.1 APRE-19细胞中4种腺苷受体的表达 在体外培养的APRE-19细胞中既可以检测到4种腺苷受体基因mRNA的表达,也可通过Western blot定性探测到这4种腺苷受体分子在APRE-19细胞膜上的存在,见图1。

Fig.1 mRNA expression and membrane distribution of adenosine receptor subtypes in ARPE-19图1 4种腺苷受体的mRNA表达及其在ARPE-19细胞膜上表达情况
2.2 RPE腺苷结合曲线及最大腺苷结合容量 RPE结合腺苷的饱和曲线显示,随着H3-adenosine加入剂量的增加,RPE对腺苷的结合逐渐达到饱和;Scat chard作图结果示RPE对腺苷的Bmax=2.04 fmol,见图2。
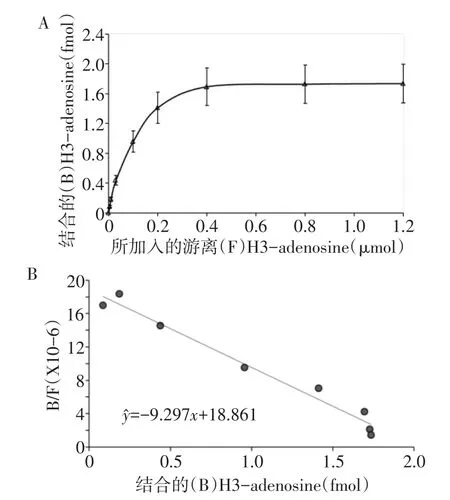
Fig.2 Results of adenosine binding assay图2 RPE腺苷结合实验结果
2.3 不同腺苷受体亚型对ARPE-19细胞结合腺苷的影响A~E组的Bmax(单位:fmol)分别为2.04±0.31、0.44±0.06、1.82±0.28、2.01±0.42 及 2.06±0.44(F=13.195,P<0.01)。其中B组较其他各组Bmax均降低(P<0.01),其他各组间比较差异均无统计学意义。
2.4 细胞因子及趋化因子的ELISA结果比较 与对照RPE组比较,CCPA干预RPE组IL-6、MCP-1及IP-10的含量降低、IL-10的含量增加(P<0.01)。2组TGF-β的含量差异无统计学意义,见表2。
Tab.2 Effects of ARA1 signaling on RPE’s production of cytokines and chemokines表2 ARA1信号通路对RPE分泌细胞因子、趋化因子的影响(n=6,ng/L,±s)

Tab.2 Effects of ARA1 signaling on RPE’s production of cytokines and chemokines表2 ARA1信号通路对RPE分泌细胞因子、趋化因子的影响(n=6,ng/L,±s)
**P<0.01
3 讨论
腺苷及其受体构成了非常复杂的免疫调节系统,在肿瘤免疫及炎症免疫中均具有巨大的潜在应用价值[12]。该系统的复杂性表现在多个方面,如受体多样性、分布广泛性[13-14]及表达高度可变性[15]。正是基于腺苷-腺苷受体系统的复杂性,本文对其进行研究时只能由单一细胞或单一组织入手再逐渐展开。本研究中real-time PCR及Western blot结果证实,4种腺苷受体亚型在ARPE-19细胞中均有表达。但是,受体的表达往往会受到负反馈调节机制的调节,即某一强烈的受体信号可以反馈性抑制该受体的表达,导致受体的表达水平及其功能可能不是正相关,甚至是负相关的[16]。与受体的表达水平不同,受体对配体的结合情况与其功能多是正相关的[17],因此,在研究受体表达的同时也要研究其配体结合情况。不同腺苷受体亚型对RPE结合腺苷的影响结果显示,ARA1拮抗剂干预组较其他各组Bmax均降低,而其他各组间差异均无统计学意义,表明ARA1是ARPE-19细胞中结合腺苷能力最强的受体亚型。研究显示,以TNF-α及IFN-γ联合干预的方式可充分激活ARPE-19细胞,使其大量分泌细胞因子及趋化因子[18]。本研究显示,在TNF-α及IFN-γ联合干预的ARPE-19细胞培养基中检测到细胞因子IL-6、IL-10、TGF-β及趋化因子MCP-1、IP-10的存在,给予ARA1的激动剂可显著性抑制ARPE-19细胞产生 MCP-1、IP-10、IL-6这些促炎因子,而促进抑炎因子IL-10的产生,提示ARA1通路与RPE细胞的免疫抑制功能密切相关。
综上所述,4种腺苷受体亚型在ARPE-19细胞中均有表达,放射性配体结合实验分析显示ARA1在ARPE-19细胞中具有相对较强的腺苷结合能力,该受体信号介导潜在的免疫抑制功能。
[1]Elliott MR,Chekeni FB,Trampont PC,et al.Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance[J].Nature,2009,461(7261):282-286.doi:10.1038/nature08296.
[2]Stagg J,Smyth MJ.Extracellular adenosine triphosphate and adenosine in cancer[J].Oncogene,2010,29(39):5346-5358.doi:10.1038/onc.2010.292.
[3]PandolfiJB,FerraroAA,SananezI,etal.ATP-Induced inflammation drives tissue-resident th17 cells in metabolically unhealthy obesity[J].J Immunol,2016,196(8):3287-3296.doi:10.4049/jimmunol.1502506.
[4]Saze Z,Schuler PJ,Hong CS,et al.Adenosine production by human B cells and B cell-mediated suppression of activated T cells[J].Blood,2013,122(1):9-18.doi:10.1182/blood-2013-02-482406.
[5]Cobbold SP,Adams E,Nolan KF,et al.Connecting the mechanisms of T-cell regulation:dendritic cells as the missing link[J].Immunol Rev,2010,236:203-218.doi:10.1111/j.1600-065X.2010.00913.x.
[6]Liu Y,Zou H,Zhao P,et al.Activation of the adenosine A2A receptor attenuates experimental autoimmune encephalomyelitis and is associated with increased intracellular calcium levels[J].Neuroscience,2016,330:150-161.doi:10.1016/j.neuroscience.2016.05.028.
[7]Li N,Mu L,Wang J,et al.Activation of the adenosine A2A receptor attenuates experimental autoimmune myasthenia gravis severity[J].Eur J Immunol,2012,42(5):1140-1151.doi:10.1002/eji.201142088.
[8]Chen S,Zhou S,Zang K,et al.CD73 expression in RPE cells is associated with the suppression ofconventionalCD4 cell proliferation[J].Exp Eye Res,2014,127:26-36.doi:10.1016/j.exer.2014.05.008.
[9]Allard D,Allard B,Gaudreau PO,et al.CD73-adenosine:a nextgeneration target in immuno-oncology[J].Immunotherapy,2016,8(2):145-163.doi:10.2217/imt.15.106.
[10]Sawynok J.Adenosine and ATP receptors[J].Handb Exp Pharmacol,2007,(177):309-328.
[11]Rico-Lattes I,Gouzy MF,Andre-Barres C,et al.Synthetic neoglycolipids for biological applications:correlation between their structures and their interactions with membrane proteins[J].Biochimie,1998,80(5/6):483-487.
[12]Young A,Ngiow SF,Barkauskas DS,et al.Co-inhibition of CD73 and A2AR adenosine signaling improves anti-tumor immune responses[J].Cancer Cell,2016,30(3):391-403.doi:10.1016/j.ccell.2016.06.025.
[13]Merighi S,Borea PA,Varani K,et al.Deregulation of adenosine receptors in psoriatic epidermis:an option for therapeutic treatment[J].J Invest Dermatol,2017,137(1):11-13.doi:10.1016/j.jid.2016.08.001.
[14]Zha D,Wu X,Gao P.Adiponectin and its receptors in diabetic kidney disease:molecular mechanisms and clinical potential[J].Endocrinology,2017,158(7):2022-2034.doi:10.1210/en.2016-1765.
[15]Sepúlveda C,Palomo I,Fuentes E.Role of adenosine A2b receptor overexpression in tumor progression[J].Life Sci,2016,166:92-99.doi:10.1016/j.lfs.2016.10.008.
[16]Quicke KM,Diamond MS,Suthar MS.Negative regulators of the RIG-I-like receptor signaling pathway[J].Eur J Immunol,2017,47(4):615-628.doi:10.1002/eji.201646484.
[17]Sengupta D,Joshi M,Athale CA,et al.What can simulations tell us about GPCRs:Integrating the scales[J].Methods Cell Biol,2016,132:429-452.doi:10.1016/bs.mcb.2015.11.007.
[18]Juel HB,Faber C,Udsen MS,et al.Chemokine expression in retinal pigment epithelial ARPE-19 cells in response to coculture with activated T cells[J].Invest Ophthalmol Vis Sci,2012,53(13):8472-8480.doi:10.1167/iovs.12-9963.
