热应激对肉仔鸡肝AMPKα1及脂肪代谢相关分子基因表达的影响
2018-01-26周华金胡希怡杨家昶丁祥文宋志刚
周华金,胡希怡,杨家昶,丁祥文,王 玉,宋志刚
(山东农业大学动物科技学院/动物医学院,泰安 271018)
热应激降低家禽生长性能,增加经济损失[1]。肉鸡因其体温高,无汗腺以及代谢旺盛等特点致使其自身蒸发散热能力有限,因而对热应激特别敏感[2]。热应激导致肉鸡采食量降低,继而影响脂肪代谢[3]。肝是禽类脂肪酸从头合成的主要场所,热应激可调节家禽肝脂肪代谢[4]。急性热应激提高肉仔鸡血清中甘油三酯、高密度脂蛋白胆固醇和低密度脂蛋白胆固醇含量,激活脂肪酸合成相关酶,增加脂肪沉积[5]。热应激提高了肝中乙酰辅酶A羧化酶α(ACACA)、酰基辅酶A合成酶(ACSF3)以及硬脂酰辅酶A脱氢酶(SCD)等脂肪代谢相关酶的基因表达量,影响脂肪代谢[6]。
肝细胞以胆固醇为原料合成胆汁酸,是肝清除胆固醇的主要方式[7]。胆汁酸分子具有特殊的表面活性,能催化脂类物质成为油水混合物的液滴,可扩大脂类物质同脂肪酶的接触面,促进脂类的消化和吸收[8]。同时,胆汁酸作为营养信号,可激活特定受体,在营养物质转运、消化和代谢中起重要作用[9]。胆汁酸信号通路在哺乳动物上引起广泛关注,不仅可以调节脂肪代谢和肠道吸收,还可作为新陈代谢调节者和分子标签,影响整个机体脂肪、葡萄糖和能量代谢[10-11]。
固醇调节元件结合蛋白-1c(Sterol regulatory element binding protein- 1c,SREBP-1c)是胆固醇和脂肪酸生物合成中的关键转录因子,优先参与脂肪酸合成相关基因的调控,可直接刺激编码脂肪酸合成酶的基因转录[12-13]。脂肪酸转运蛋白1(Fatty acid transport protein1,FATP-1)促进脂肪细胞吸收脂肪酸,在脂肪细胞的分化中发挥重要作用[14]。载脂蛋白B(Apolipoprotein B,APOB)是肝中转运内源性甘油三酯和胆固醇的功能蛋白,在鸡肝、肾组织中有较高表达,与脂肪的沉积相关[15]。肝X受体(Liver X receptor,LXR)和法尼酯X受体(Farnesoid X receptor,FXR)在胆汁酸和胆固醇代谢过程中起重要的调节作用,通过对靶基因表达的调节来维持脂肪代谢的平衡。胆固醇7-羟化酶(Cholesterol 7-alpha hydroxyl-lase,CYP7A1)是胆固醇转化为胆汁酸过程中主要的限速酶[16]。AMP活化蛋白激酶(AMP-activated protein kinase,AMPK)参与调节细胞代谢和整个机体的新陈代谢活动,作为能量感受器维持内环境的稳定。AMPK参与脂肪的代谢调节,外周AMPK的磷酸化降低脂肪酸合成酶的转录活性,促进脂肪酸氧化[17]。
有关热应激持续时间对肝脂肪代谢影响的研究不多,未见关于AMPK在应激调控肝脂肪代谢中作用的报道。本试验通过探究长、短期热应激对肉仔鸡肝AMPKα1及脂肪代谢相关分子基因表达的影响,以期扩展热应激影响肉仔鸡肝脂肪代谢的理论,为生产中防控肉仔鸡热应激提供科学参考。
1 材料与方法
1.1 试验设计
体重相近的1日龄雄性AA肉仔鸡(Arbor Acres) 120只(购自泰安大宝孵化场),严格控制饲养环境。试验开始时饲养温度35 ℃,根据肉鸡日龄每周降低2~3 ℃,直到降到23 ℃。环境湿度为40%~50%,温度和湿度随肉鸡生长情况做相应调整。光照程序按照23 h光照、1 h黑暗执行。前期日粮粗蛋白含量为21%、代谢能为12.55 MJ·kg-1,后期日粮粗蛋白含量为19%、代谢能为12.97 MJ·kg-1。饲养阶段自由采食和饮水。28日龄选取体重相近的健康肉鸡96只,随机分为4组(每组4个重复,每个重复6只鸡):24 h热应激组和24 h对照组、72 h热应激组和72 h对照组。28日龄08:00开始试验,统计各组鸡体重、余料量,记录加料量,热应激处理组温度为(33±1)℃ ,对照组(23±1)℃。29和31日龄上午08:00每个组在饲喂前称重,每次称取鸡只的体重和剩余料量,计算热应激24和72 h的采食量和体增重,每个重复选取2只鸡,翅静脉采集血液后4 ℃、400 g离心10 min,取上清放置于-20 ℃保存,而后屠宰,采集肝样品,用冰冷的生理盐水漂洗新鲜的肝样品后,液氮速冻,放于-80 ℃保存。
1.2 血浆指标测定
血浆中葡萄糖(GLU)、尿酸(UA)、总胆固醇(TCHO)、总胆汁酸 (TBA)、甘油三脂(TG) 利用比色法采用分光光度计进行测定,试剂盒购自南京建成试剂有限公司。
1.3 RT-PCR对基因表达的测定
肝总RNA用Trizol法提取,利用琼脂糖凝胶电泳和生物分光光度计(Biophotometer plusone,德国 Eppendorf)分别检测总RNA的质量和浓度。按照TaKaRa RNA PCR 反转录试剂盒操作说明进行反转录。反应体系:500 ng总RNA, 5 mmol·L-1MgCl2, 1 μL RT buffer, 1 mmol·L-1dNTP, 2.5 U 逆转录酶(AMV), 0.7 mmol·L-1Oligod (T) 和10 U Ribonuclease inhibitor,加DEPC水至10 μL。42 ℃反应40 min, 99 ℃灭活5 min,降低到5 ℃反应 5 min。cDNA合成以后进行荧光定量PCR。引物序列见表1,由上海生工生物技术有限公司合成。通过对混合样品进行标准曲线测定确定引物的质量和最佳的稀释浓度。反应体系为20 μL∶10 μL SYBR Premix Ex TaqTM (2×),上游引物和下游引物(10 μmol·L-1)各0.4 μL, 0.4 μL ROX Reference Dye Ⅱ(50×), 2 μL cDNA模板和6.8 μL去离子水。两步法荧光定量PCR反应条件:第一步95 ℃变性 10 s, 1个循环,第二步95 ℃变性5 s,60 ℃退火34 s, 40个循环。
表1引物序列
Table1Gene-specificprimersofrelatedgenes
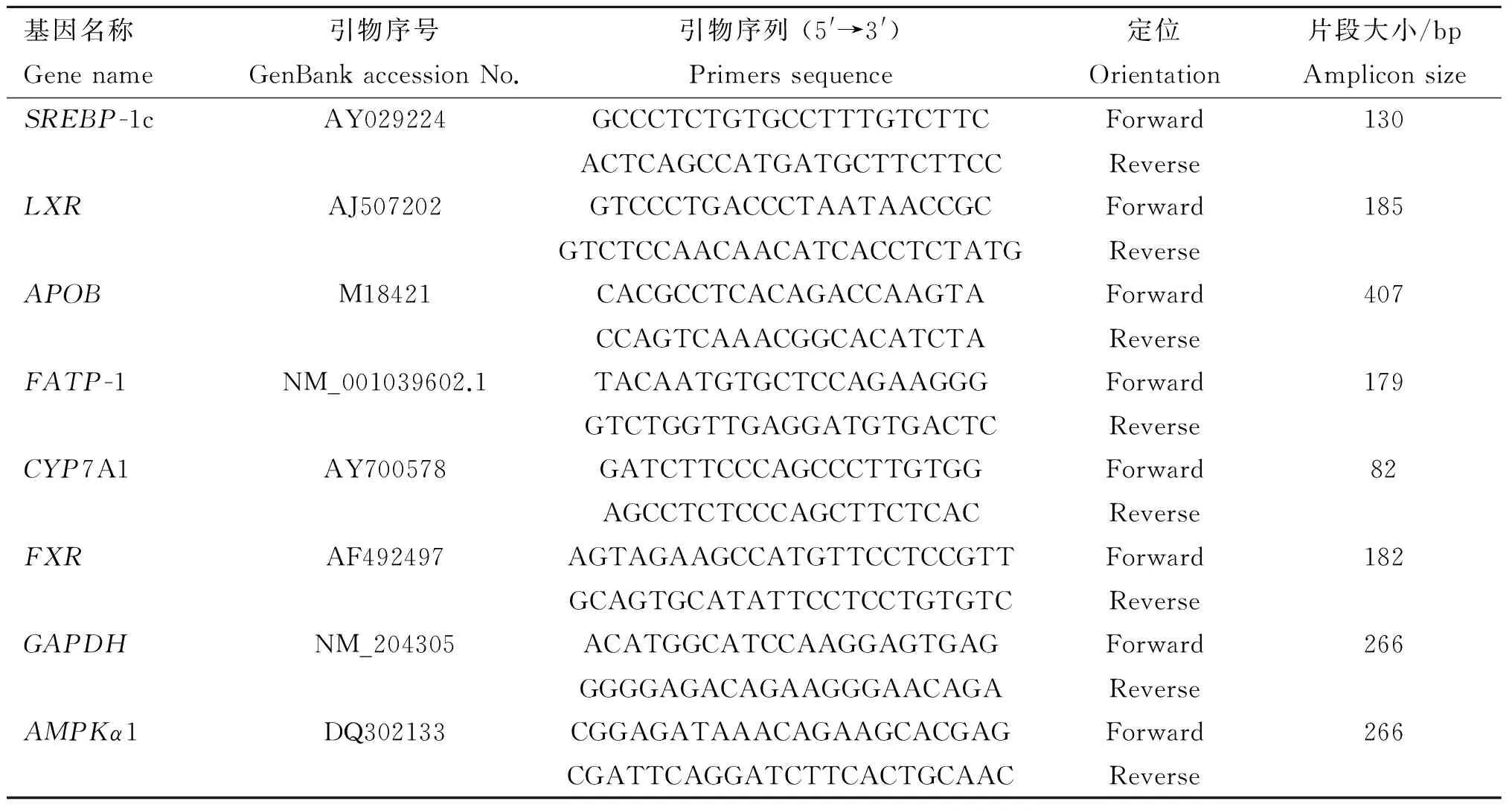
基因名称Genename引物序号GenBankaccessionNo.引物序列(5'→3')Primerssequence定位Orientation片段大小/bpAmpliconsizeSREBP-1cAY029224GCCCTCTGTGCCTTTGTCTTCACTCAGCCATGATGCTTCTTCCForwardReverse130LXRAJ507202GTCCCTGACCCTAATAACCGCGTCTCCAACAACATCACCTCTATGForwardReverse185APOBM18421CACGCCTCACAGACCAAGTACCAGTCAAACGGCACATCTAForwardReverse407FATP-1NM_001039602.1TACAATGTGCTCCAGAAGGGGTCTGGTTGAGGATGTGACTCForwardReverse179CYP7A1AY700578GATCTTCCCAGCCCTTGTGGAGCCTCTCCCAGCTTCTCACForwardReverse82FXRAF492497AGTAGAAGCCATGTTCCTCCGTTGCAGTGCATATTCCTCCTGTGTCForwardReverse182GAPDHNM_204305ACATGGCATCCAAGGAGTGAGGGGGAGACAGAAGGGAACAGAForwardReverse266AMPKα1DQ302133CGGAGATAAACAGAAGCACGAGCGATTCAGGATCTTCACTGCAACForwardReverse266
参照文献[18]的方法用2-ΔΔCT法定量目标基因相对表达量,以GAPDH作为参照基因进行校准
According to reference [18], the relative expression of target genes were determined by 2-ΔΔCTmethod, usingGAPDHas the reference gene for calibration
1.4 数据分析
数据采用SAS (Version 8e,SAS Institute,1998)统计软件ANOVA法进行单因子方差分析,试验数据用“平均值±标准误(Mean±SE)”表示,P<0.05表示处理效应差异显著。
2 结 果
2.1 热应激对肉仔鸡生长性能的影响
由图1可知,24 与72 h热应激均显著降低肉仔鸡平均体增重(P<0.05);由图2可知,24 与72 h热应激均显著降低肉仔鸡平均采食量(P<0.05)。
2.2 热应激对肉仔鸡血液生化指标的影响
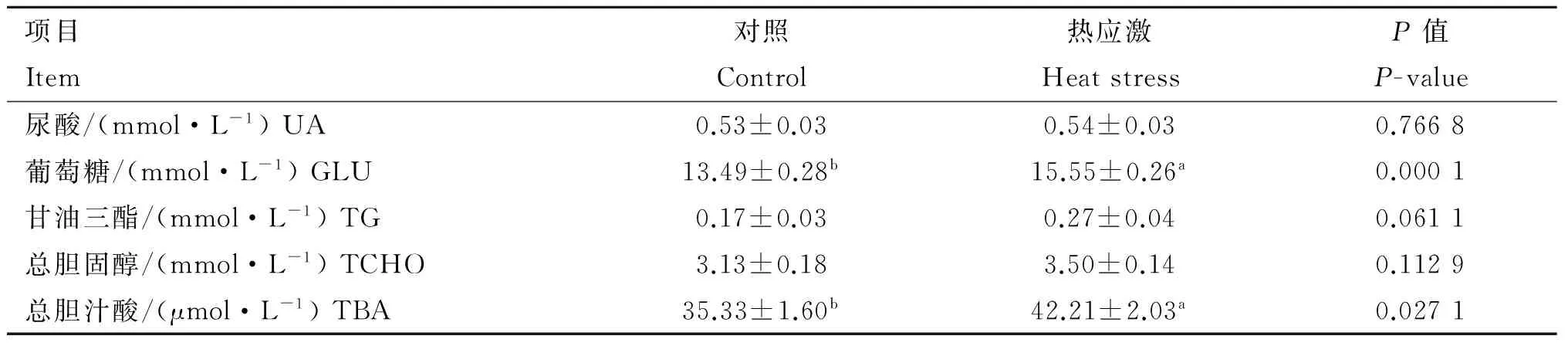
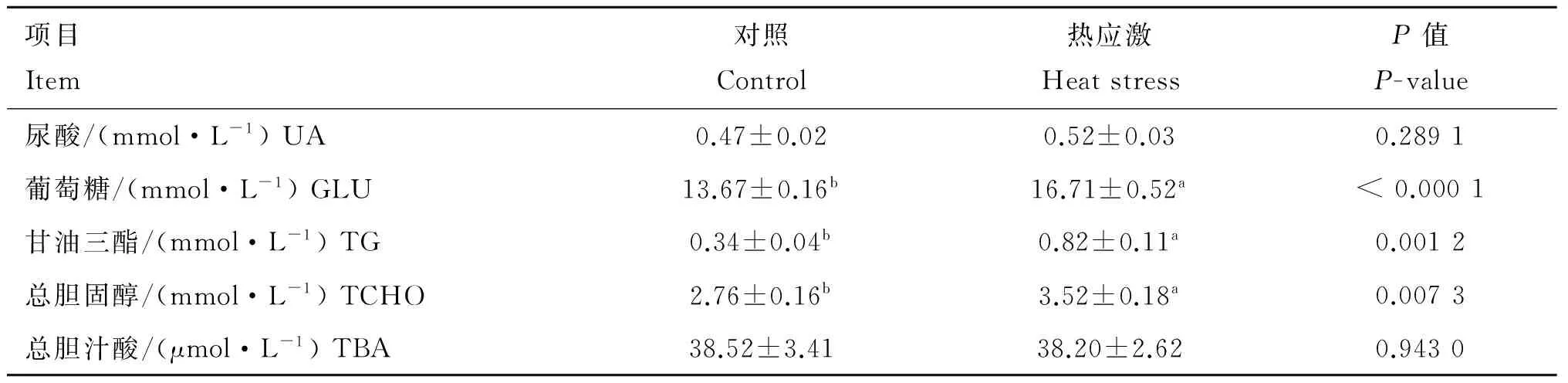
由表2可知,与对照组相比,24 h热应激显著高肉仔鸡血浆中葡萄糖(GLU)和总胆汁酸(TBA)含量(P<0.05);由表3可知,与对照组相比,72 h热应激显著提高肉仔鸡血浆中葡萄糖(GLU)、甘油三脂(TG)和总胆固醇(TCHO)含量(P<0.05)。

柱形图上所标字母相异表示差异显著(P<0.05),未标字母表示差异不显著(P>0.05)。下图同Different letters on the column charts indicate significant differences between treatments (P<0.05), no letters on the column charts mean no significant difference between treatments (P>0.05). The following figures are the same图1 热应激24 和72 h对肉仔鸡体增重的影响Fig.1 Effects of heat stress for 24 and 72 h on the body weight gain of broiler chickens

图2 热应激24和72 h对肉仔鸡采食量的影响Fig.2 Effects of heat stress for 24 and 72 h on feed intake of broiler chickens
2.3 热应激对肉仔鸡脂肪代谢相关基因表达的影响
由图3可知,与对照组相比,24 h热应激对肉仔鸡肝中FATP-1、SREBP-1c、APOB和LXR基因表达均无显著影响(P>0.05) (图3A);72 h热应激显著上调肉仔鸡肝中FATP-1、SREBP-1c、APOB和LXR基因的相对表达量(P<0.05)(图3B)。
2.4 热应激对肉仔鸡肝CYP7A1信号通路相关基因表达的影响
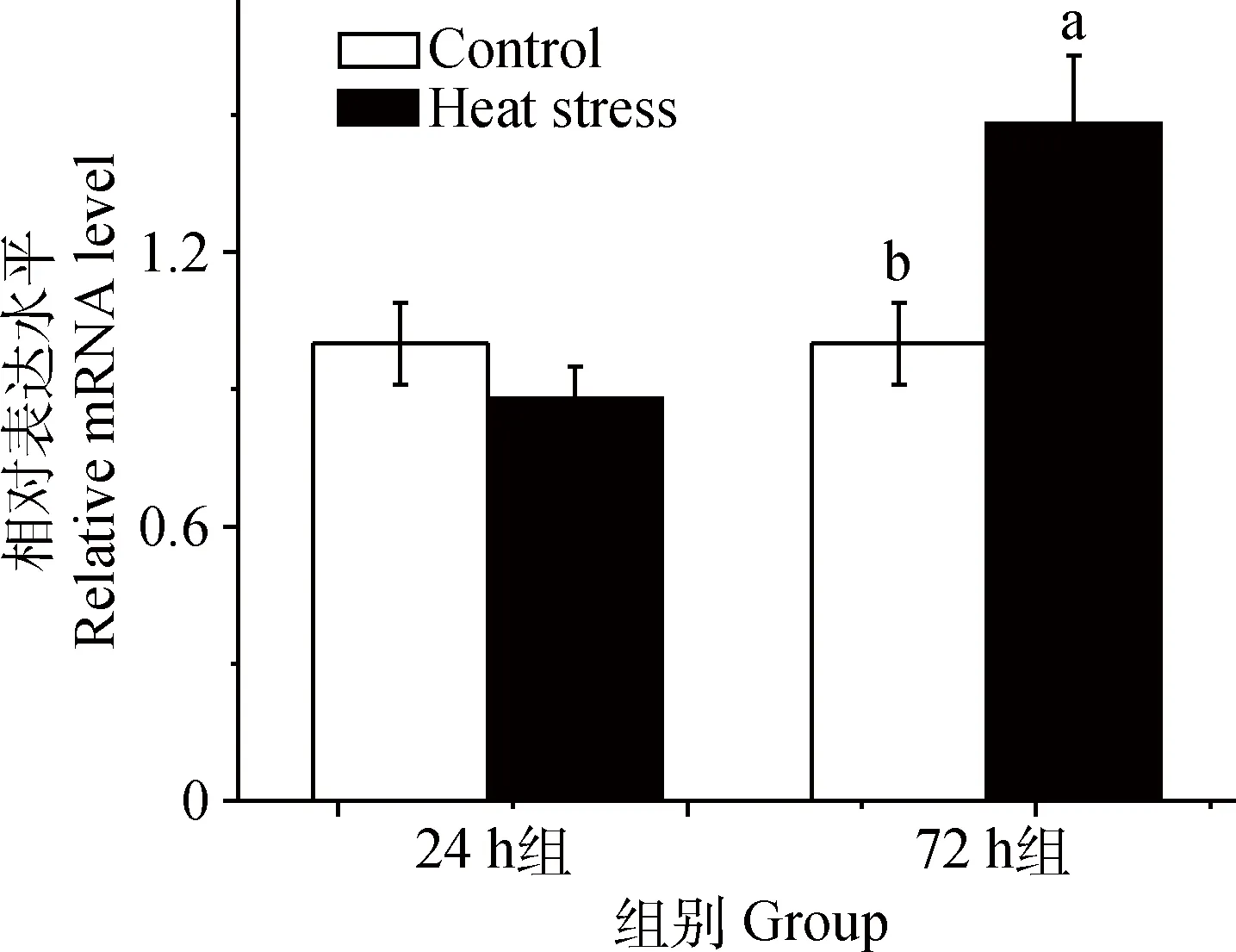
由图4可知,与对照组相比,24 h热应激对肉仔鸡肝中FXR和CYP7A1基因表达无显著性影响(P>0.05)(图4A);72 h热应激对肉仔鸡肝中FXR和CYP7A1基因表达有增加的趋势(0.05 由图5可知,与对照组相比,24 h热应激对肉仔鸡肝中AMPKα1 基因表达没有造成显著影响(P>0.05);72 h热应激显著上调肉仔鸡肝中AMPKα1基因表达(P<0.05)。 表2热应激24h对肉仔鸡血液生化指标的影响 Table2Effectsof24-hourheatstressonplasmametabolitesinbroilerchickens 项目Item对照Control热应激HeatstressP值P-value尿酸/(mmol·L-1)UA0.53±0.030.54±0.030.7668葡萄糖/(mmol·L-1)GLU13.49±0.28b15.55±0.26a0.0001甘油三酯/(mmol·L-1)TG0.17±0.030.27±0.040.0611总胆固醇/(mmol·L-1)TCHO3.13±0.183.50±0.140.1129总胆汁酸/(μmol·L-1)TBA35.33±1.60b42.21±2.03a0.0271 同行数据后所标字母相异表示差异显著(P<0.05),未标字母表示差异不显著(P>0.05)。下同 Different letters in the same row means significant difference between treatments (P<0.05), no letters in the same row means not significant difference between treatments (P>0.05).The same as below 表3热应激72h对肉仔鸡血液生化指标的影响 Table3Effectsof72-hourheatstressonplasmametabolitesinbroilerchickens 项目Item对照Control热应激HeatstressP值P-value尿酸/(mmol·L-1)UA0.47±0.020.52±0.030.2891葡萄糖/(mmol·L-1)GLU13.67±0.16b16.71±0.52a<0.0001甘油三酯/(mmol·L-1)TG0.34±0.04b0.82±0.11a0.0012总胆固醇/(mmol·L-1)TCHO2.76±0.16b3.52±0.18a0.0073总胆汁酸/(μmol·L-1)TBA38.52±3.4138.20±2.620.9430 图3 24 和72 h热应激对肉仔鸡肝脂肪代谢相关基因表达的影响Fig.3 Effects of heat stress for 24(A) and 72 h(B) on the mRNA expression of FATP-1, SREBP-1c, APOB and LXR in livers of broiler chickens 图4 24 和72 h热应激对肉仔鸡肝CYP7A1信号通路相关基因表达的影响Fig.4 Effect of heat stress for 24(A) and 72 h(B) on the genes expression in CYP7A1 signal pathway in livers of broiler chickens 图5 24和72 h热应激对肉仔鸡肝AMPKα1基因表达的影响Fig.5 Effect of heat stress for 24 and 72 h on gene expression of AMPKα1 in livers of broiler chickens 高温可直接引起家禽采食量的降低。本研究结果显示,热应激显著降低肉仔鸡平均体增重和采食量,这与前期研究结果一致[19-21]。已有研究表明,热应激诱导下丘脑-垂体-肾上腺皮质轴(Hypothalamic-pituitary-adrenal, HPA)的激活,肾上腺皮质大量释放皮质酮影响家禽采食行为是造成生产性能下降的主要原因[22]。 与相近体重的哺乳动物相比较,家禽血浆中含有较高的血糖浓度[23]。本研究结果显示,24 和72 h热应激均显著提高血浆中葡萄糖含量,此外,72 h热应激显著提高了血浆中甘油三脂和胆固醇含量。相关研究显示,机体在应激条件下通过促进体内葡萄糖的动员和生产提高血糖水平以维持稳态[24]。短期应激下,HPA轴通过与肾上腺素、胰岛素、胰高血糖素和交感神经系统的互作,提高血糖水平,满足机体重要器官的能量供给[20]。而在长期应激时,HPA轴糖皮质激素的负反馈调节障碍,会持续促进糖皮质激素分泌,使脂肪和肌肉组织的脂肪动员和肝糖异生加强,导致游离脂肪酸和血糖水平升高[25]。本研究结果表明,长期热应激时,机体肝脂肪代谢受到显著影响。 对于鸟类而言,肝是脂肪生成的主要场所[26]。家禽中甘油三脂主要在肝中合成后输出,在皮下脂肪、肝及肌肉组织等器官中贮存[27-28]。FATP-1是重要的转运蛋白之一,参与脂肪酸跨膜转运和脂肪沉积[29]。报道证明,FATP-1不但直接调节脂肪酸跨膜转运,而且催化细胞内脂肪酸转化为酰基辅酶A,优先合成细胞内甘油三脂[30-31]。有研究表明,FATP-1敲除后显著降低脂肪细胞甘油三脂的含量[31]。SREBPs为膜转录因子家族,能够参与调节脂类合成基因的表达。SREBP-1c在高脂肪合成组织中高度表达,例如肝和脂肪组织[32]。LXR激活的SREBP-1c可通过转录激活脂肪合成过程中的相关基因,激活脂肪合成通路[33]。APOB在决定肝细胞合成和分泌VLDL的能力中起重要作用[29,34]。APOB是形成极低密度脂蛋白的必要成分,翻译过程中和翻译前修饰对VLDL合成、分泌、肝脂肪平衡很重要[35]。哺乳动物及家禽上,LXRs在脂类和胆固醇代谢上起重要调节作用,例如胆汁酸和脂肪酸的合成[36-37]。此外,LXRα不仅直接调控脂肪生成转录因子的表达,而且直接调控脂肪合成酶的表达,加强肝脂肪酸的合成[38]。研究表明,饲喂高胆固醇日粮的小鼠缺乏LXRα,显著增加肝胆固醇的沉积[39]。本试验结果表明,短期热应激没有影响肝中脂肪调控因子基因表达,长期热应激显著上调肝中FATP-1、SREBP-1C、APOB和LXR脂肪调控因子基因的表达,说明短期热应激没有影响肝脂肪代谢,长期热应激则显著调动了能量的重新分配,使肝脂肪合成增加。 肝是机体胆固醇代谢的主要部位,游离脂肪酸分泌到胆汁或者进一步转化为胆汁酸,该过程主要受微粒体酶CYP7A1调节[40]。CYP7A1主要参与肝由胆固醇合成胆汁酸和胆固醇的过程[41]。使用LXR激活剂TO-901317处理家禽肝细胞,显著性上调SREBP1、FAS、CYP7A1基因表达[27]。FXR作为胆汁酸的感受器,在肝细胞中激活后,调节胆汁酸的合成、排出和肠道中重新吸收,从而维持体内胆固醇的稳态。在小鼠上,FXR抑制胆汁酸生成,而LXR促进胆汁酸生成。细胞中高浓度的胆汁酸是有毒的,因此,FXR对维持机体胆汁酸水平有重要作用[42]。小鼠FXR缺乏导致肝生成胆汁酸含量增加,干扰脂肪代谢,导致肝变性、炎症和纤维化[43-44]。本试验结果表明,短期应激提高了肉仔鸡血浆中的胆汁酸含量,但没有影响肝中FXR、CYP7A1基因表达,说明短期热应激时血浆中胆汁酸含量增加可能跟肝中胆汁酸合成途径无关;而长期热应激时,血浆中胆汁酸含量未受影响,但是肝中CYP7A1和FXR基因表达有升高的趋势,这是否是机体对抗热应激的一种负反馈反应,值得进一步研究。 AMPK在调控整个机体能量代谢上起重要作用,脂肪细胞生成的激素,例如瘦素和脂联素也会影响AMPK,瘦素可激活骨骼肌中AMPK增加脂肪酸氧化,同时脂联素可以激活肝中的AMPK,增加糖的利用和脂肪酸氧化,抑制肝中葡萄糖的生成[45]。AMPK是主要的细胞能量状态感受器,AMP与ATP之比增加会激活AMPK[46]。作为能量感受器和代谢传感器,AMPK可影响鸟类肝脂肪代谢,在短期的调节过程中,AMPK磷酸化抑制ACCα,继而抑制丙二酰辅酶A和脂肪酸合成,AMPK控制肝葡萄糖和脂肪代谢,影响整个机体的能量利用[47-48]。外周AMPK激活后会关闭机体ATP生成的代谢过程,例如脂肪组织和肝中游离脂肪酸氧化[49-50]。本研究表明,24 h热应激对肝AMPKα1基因的表达无显著影响,而72 h热应激则显著提高肝AMPKα1基因表达,这一变化模式跟肝中脂肪代谢相关分子的变化一致,说明AMPK信号通路可能在应激影响肝脂肪代谢的机制中起着重要作用。 热应激显著降低肉仔鸡生产性能,短期热应激对肉仔鸡肝脂肪代谢影响有限,长期热应激则显著改变肉仔鸡肝脂肪代谢,AMPK可能参与热应激对肝脂肪代谢的调控。 [1] SONG Z G, LIU L, SHEIKHAHMADI A, et al. Effect of heat exposure on gene expression of feed intake regulatory peptides in laying hens[J].JBiomedBiotechnol, 2012, 2012: 484869. [2] NICOL C. Behaviour as an indicator of animal welfare[M]//Webster J. Management and Welfare of Farm Animals: The UFAW Farm Handbook. Chichester, UK: Blackwell Wiley, 2011: 31-67. [3] SUN X L, ZHANG H C, SHEIKHAHMADI A, et al. Effects of heat stress on the gene expression of nutrient transporters in the jejunum of broiler chickens (Gallusgallusdomesticus)[J].IntJBiometeorol, 2015, 59(2): 127-135. [4] GERAERT P A, PADILHA J C, GUILLAUMIN S. Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: Biological and endocrinological variables[J].BritJNutr, 1996, 75(2): 205-216. [5] 刘 梅. 急性热应激对肉仔鸡生长性能及脂肪代谢的影响[J]. 动物营养学报, 2011, 23(5): 862-868. LIU M. Effects of acute heat stress on growth performance and lipid metabolism of broilers [J].ChineseJournalofAnimalNutrition, 2011, 23(5): 862-868. (in Chinese) [6] JASTREBSKI S F, LAMONT S J, SCHMIDT C J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis[J].PLoSOne, 2017, 12(7): e0181900. [7] 王会敏, 王正平, 董旻岳. 胆汁酸代谢与调控研究进展[J]. 国际消化病杂志, 2010, 30(2): 79-82. WANG H M, WANG Z P, DONG M Y. Research progress about metabolism and regulation of bile acids [J].InternationalJournalofDigestiveDiseases, 2010, 30(2): 79-82. (in Chinese) [9] LI T G, CHIANG J Y L. Bile acid signaling in metabolic disease and drug therapy[J].PharmacolRev, 2014, 66(4): 948-983. [10] PRAWITT J, CRON S, STAELS B. Bile acid metabolism and the pathogenesis of type 2 diabetes[J].CurrDiabRep, 2011, 11: 160. [11] THOMAS C, PELLICCIARI R, PRUZANSKI M, et al. Targeting bile-acid signalling for metabolic diseases [J].NatRevDrugDiscovery, 2008, 7: 678-693. [12] KHESHT F A, HASSANABADI A. Effects of sterol regulatory element-binding protein (SREBP) in chickens[J].LipidsHealthDis, 2012, 11: 20. [13] 朱玉萍,周 平,李蛟龙,等.低蛋白氨基酸平衡日粮添加半胱胺对生长猪肉质和相关基因表达的影响[J].畜牧兽医学报,2017,48(4):660-668. ZHU Y P, ZHOU P, LI J L, et al. Effects of low protein level diets supplemented with essential amino acids and Cysteamine on meat quality and related genes expression of growing pigs[J].ActaVeterinariaetZootechnicaSinica, 2017, 48(4): 660-668.(in Chinese) [14] 齐仁立, 黄金秀, 杨飞云, 等. 脂肪酸转运蛋白家族及其介导的脂肪酸跨膜转运[J]. 动物营养学报, 2013, 25(5): 905-911. QI R L, HUANG J X, YANG F Y, et al. Fatty acid transport protein family and their mediation in transmembrane transportation of fatty acids[J].ChineseJournalofAnimalNutrition, 2013, 25(5): 905-911. (in Chinese) [15] ZHANG S, SHI H, LI H. Cloning and tissue expression characterization of the chickenAPOBgene[J].AnimBiotechnol, 2007, 18(4): 243-250. [16] PIEKARSKI A, DECUYPERE E, BUYSE J, et al. Chenodeoxycholic acid reduces feed intake and modulates the expression of hypothalamic neuropeptides and hepatic lipogenic genes in broiler chickens[J].GenCompEndocrinol, 2016, 229: 74-83. [18] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitativeI PCR and the 2-ΔΔCTmethod[J].Methods, 2001, 25(4): 402-408. [19] LIN H, BUYSE J, DU R, et al. Response of rectal temperature of broiler chickens to thermal environment factors[J].ArchivFurGeflugelkunde, 2004, 68(3): 126-131. [20] LIN H, DECUYPERE E, BUSE J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallusgallusdomesticus): 2. Short-term effect[J].CompBiochemPhysiolBiochemMolBiol, 2004, 139(4): 745-751. [21] LIN H, ZHANG H F, JIAO H C, et al. Thermoregulation responses of broiler chickens to humidity at different ambient temperatures. I. One week of age[J].PoultrySci, 2005, 84(8): 1166-1172. [22] QUINTEIRO-FILHO W M, RIBEIRO A, FERRAZDE-PAULA V, et al. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens [J].PoultrySci, 2010, 89(9): 1905-1914. [23] BRANU E J, SWEAZEA K L. Glucose regulation in birds[J].CompBiochemPhysiolBiochemMolBiol, 2008, 151(1): 1-9. [24] VIRDEN W S, KIDD M T. Physiological stress in broilers: ramifications on nutrient digestibility and responses[J].JApplPoultRes, 2009, 18(2): 338-347. [25] LIN H, DECUYPERE E, BUYSE J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallusgallusdomesticus): 1. Chronic exposure[J].CompBiochemPhysiolBiochemMolBiol, 2004, 139(4): 737-744. [26] GREENE E, KHALDI S, ISHOLA P, et al. Heat and oxidative stress alter the expression of orexin and its related receptors in avian liver cells[J].CompBiochemPhysiolAMolIntegrPhysiol, 2015, 191: 18-24. [27] EVANS A J.Invitrolipogenesis in the liver and adipose tissues of the female Aylesbury duck at different ages[J].BrPoultSci, 1972, 13(6): 595-602. [28] HIRSCH D, STAHL A, LODISH H F. A family of fatty acid transporters conserved from mycobacterium to man[J].ProcNatlAcadSciUSA, 1998, 95(15): 8625-8629. [29] DAVIS R A, BOOGAERTS J R, BORCHARDT R A, et al. Intrahepatic assembly of very low density lipoproteins. Varied synthetic response of individual apolipoproteins to fasting[J].JBiolChem, 1985, 260(26): 14137-14144. [30] HATCH G M, SMITH A J, XU F Y, et al. FATP1 channels exogenous FA into 1,2,3-triacyl-sn-glycerol and down-regulates sphingomyelin and cholesterol metabolism in growing 293 cells[J].JLipidRes, 2002, 43(9): 1380-1389. [31] LOBO S, WICZER B M, SMITH A J, et al. Fatty acid metabolism in adipocytes: functional analysis of fatty acid transport proteins 1 and 4[J].JLipidRes, 2007, 48(3): 609-620. [32] COMMERFORD S R, PENG L, DUBÉ J J, et al.Invivoregulation of SREBP-1c in skeletal muscle: effects of nutritional status, glucose, insulin, and leptin[J].AmJPhysiolRegulIntegrCompolPhysiol, 2004, 287(1): R218- R227. [33] CHU K, MIYAZAKI M, MAN W C, et al. Stearoyl-coenzyme A desaturase 1 deficiency protects against hypertriglyceridemia and increases plasma high-density lipoprotein cholesterol induced by liver X receptor activation[J].MolCellBiol, 2006, 26(18): 6786-6798. [34] BORÉN J, WETTESTEN M, RUSTAEUS S, et al. The assembly and secretion of APOB-100-containing lipoproteins[J].BiochemSocTrans, 1993, 21(2): 487-493. [35] WANG S Y, PARK S, KODLI V K, et al. Identification of protein disulfide isomerase 1 as a key isomerase for disulfide bond formation in apolipoprotein B100[J].MolBiolCell, 2014, 26(4): 594-604. [37] SATO K, KAMADA T. Regulation of bile acid, cholesterol, and fatty acid synthesis in chicken primary hepatocytes by different concentrations of T0901317, an agonist of liver X receptors[J].CompBiochemPhysiolAMolIntegrPhysiol, 2011, 158(2): 201-206. [38] DEMEURE O, DUBY C, DESERT C, et al. Liver X receptor α regulates fatty acid synthase expression in chicken[J].PoultrySci, 2009, 88(12): 2628-2635. [39] PEET D J, TURLEY S D, MA W Z, et al. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXRα[J].Cell, 1998, 93(5): 693-704. [40] RUDLIG M, ANGELIN B, STAHLE L, et al. Regulation of hepatic low-density lipoprotein receptor, 3-hydroxy-3-methylglutaryl coenzyme A reductase, and cholesterol 7α-hydroxylase mRNAs in human liver[J].JClinEndocrinolMetabol, 2002, 87(9): 4307-4313. [41] BJÖRKHEM I, ARAYA Z, RUDLING M, et al. Differences in the regulation of the classical and the alternative pathway for bile acid synthesis in human liver. No coordinate regulation of CYP7A1 and CYP27A1[J].JBiolChem, 2002, 277(30): 26804-26807. [42] CALKIN A C, TONTONOZ P. Transcriptional integration of metabolism by the nuclear sterol-activated receptors LXR and FXR[J].NatRevMolCellBiol, 2012, 13(4): 213-224. [43] KIM I, AHN S -H, INAGAKI T, et al. Differential regulation of bile acid homeostasis by the farnesoid X receptor in liver and intestine[J].JLipidRes, 2007, 48(12): 2664-2672. [44] SINAL C J, TOHKIN M, MIYATA M, et al. Targeted disruption of the nuclear receptor FXR/BAR impairs bile acid and lipid homeostasis[J].Cell, 2000, 102(6): 731-744. [45] ANDERSSON U, FILIPSSON K, ABBOTT C R, et al. AMP-activated protein kinase plays a role in the control of food intake[J].JBiolChem, 2004, 279(13): 12005-12008. [46] ALEXANDER A, WALKER C L. The role of LKB1 and AMPK in cellular responses to stress and damage[J].FEBSLett, 2011, 585(7): 952-957. [47] VIOLLET B, FORETZ M, GUIGAS B, et al. Activation of AMP-activated protein kinase in the liver: a new strategy for the management of metabolic hepatic disorders[J].JPhysiol, 2006, 574(1): 41-53. [48] VIOLLET B, GUIGAS B J, HÉBRARD S, et al. AMP-activated protein kinase in the regulation of hepatic energy metabolism: from physiology to therapeutic perspectives[J].ActaPhysiol, 2009, 196(1): 81-98. [49] ZHANG B B, ZHOU G C, LI C. AMPK: an emerging drug target for diabetes and the metabolic syndrome[J].CellMetab, 2009, 9(5): 407-416. [50] RUDERMAN N B, SAHA A K, KRAEGEN E W. Minireview: malonyl CoA, AMP-activated protein kinase, and adiposity[J].Endocrinology, 2004, 144(12): 5166-5171.2.5 热应激对肉仔鸡肝AMPK的影响


3 讨 论
3.1 热应激对肉仔鸡生长性能的影响
3.2 热应激对肉仔鸡肝脂肪调控因子的影响
3.3 热应激对肉仔鸡肝CYP7A1信号通路的影响
3.4 热应激对肉仔鸡肝AMPK的影响
4 结 论
