SOX5影响小鼠皮肤黑色素细胞MITF-M表达和黑色素生成
2018-01-26常露程赵兵令李经纬李莉鑫王海东
常露程,赵兵令,刘 颖,白 云,李经纬,李莉鑫,王海东
(山西农业大学动物科技学院,太谷 030801)
SOX基因家族是拥有一个或多个高迁移率组转录因子SRY相关的HMG盒,参与神经调节、骨组织的发育等多种早期胚胎发育过程[1]。SOX家族根据HMG的保守程度又分为8组:A~H组,而SRY sex determining region Y-box 5 (SOX5)是SOXD中的一个基因[2]。SOX5在软骨细胞、神经细胞、以及黑色素细胞中都有重要的调节作用[3-5],在黑色素瘤细胞中SOX5通过调节小眼畸形相关转录因子(Microphthalmia-associated transcription factor,MITF)来调控黑色素瘤细胞的增殖[6]。MITF与转录因子EB、TFE3和TFEG一起构成了MIT家族[7],而MITF是MIT家族中唯一对正常黑色素细胞发育有重要作用的因子[8]。MITF基因是多启动子结构,至少有9种不同的启动子-外显子单元指导MITF的启动[9],而只有M启动子(MITF-M)在黑素细胞中选择性表达[10]。MITF-M是黑色素细胞特异性调节因子,在黑色素瘤和色素沉着过度疾病中具有关键作用[11-13]。MITF-M直接调控相关色素基因的转录,包括酪氨酸酶(Tyrosinase,TYR)[14]、酪氨酸酶相关蛋白(Tyrosinase-related protein 1,TYRP1)[15]、Tyrosinase-related protein 2 (TYRP2)、前黑素体蛋白(Premelanosome protein,PMEL-17)[16](也称PMEL)和Ocular albanism 1 (OA1)[17-18]等。TYR、TYRP1和TYRP2 3种主要色素沉着相关酶被认为是MITF转录的靶基因且已被人所熟知[7,19]。PMEL依赖性MITF表达,其蛋白质和mRNA水平受MITF的转录调控[16,20]。并且PMEL是负责在黑素体内色素细胞功能所必需,缺乏或突变PMEL表达的动物均显示不同程度的色素沉着不足[21]。T.Z.Chen等[22]提出OA1可能通过调节MITF的水平以及黑素体的数量、大小、能动性和成熟,参与毛色的形成。OA1在调节MITF表达中的作用参与α-MSH-MITF信号传导途径[23],并且OA1功能的丧失将大大降低MITF的表达,尽管其水平仍足以维持色素细胞的存活和分化状态[24]。SOX5在不同毛色小鼠皮肤中定位以及表达是否存在一定的线性关系从而参与毛色的形成,以及在小鼠黑色素细胞中SOX5是如何调控黑色素的合成,目前未见相关报道。通过不同的生物学方法对SOX5进行检测,从而确定SOX5与毛色形成的相关性以及为研究SOX5调控黑色素合成路径奠定基础。
1 材料与方法
1.1 试剂与材料
MELM(2201) (SCIENCELL)、SOX5、TYR、TYRP1和TYRP2兔抗多克隆抗体(Abcam公司)、MITF兔抗多克隆抗体(武汉三鹰公司)、OA1兔抗多克隆抗体(Santa公司)、X-tremeGENE HP DNA Transfection Reagent(上海宇博生物公司)、C57BL/6品系小鼠、C57BL/6小鼠黑色素细胞5代、慢病毒载体pLV.Des3d.P/Puro(本实验室提供)。
小鼠品系为C57BL/6,由山西农业大学国家级动物医学实验教学示范中心动物室提供,随机选取出生后12 d的黑色、棕色、灰色小鼠各3 只,背部刮毛每只小鼠各取3块皮肤组织,其中2块组织用于总蛋白质及总RNA的提取。其余1 块在Bouin’s液中固定,用于小鼠皮肤毛囊组织切片制作。
1.2 RNA提取和qRT-PCR检测
Trizol法提取不同毛色小鼠皮肤和每组细胞的总RNA,测定其浓度后反转录。在NCBI上检索小鼠SOX5、MITF-M、TYR、TYRP1、TYRP2、PMEL、OA1序列,利用Premier 5.0软件设计PCR扩增引物,送北京华大基因公司合成。引物序列和产物长度见表1,退火温度为58 ℃。
按照SYBR®Premix Ex TapTMⅡ试剂盒说明书进行荧光定量PCR,通过2-ΔΔCT法计算目的mRNA相对表达量变化。
1.3 蛋白提取和Western blotting检测
根据碧云天裂解液试剂盒说明书提取不同毛色小鼠皮肤组织和各组黑色素细胞的总蛋白,每个样品总蛋白上样量为200 μg,待SDS-PAGE电泳结束后,转移至NC膜;NC膜经5 %脱脂奶粉室温封闭1 h;加入SOX5(1∶500),MITF、TYR、TYRP1、TYRP2(1∶1 000),OA1(1∶400)以及β-Actin(1∶2 000)一抗4 ℃过夜孵育;次日NC膜用TBST摇洗3次(每次10 min);加入HRP标记的二抗(1∶10 000)37 ℃恒温水平摇床孵育NC膜1 h。NC膜用TBST摇洗6次(每次5 min),使用ECL试剂盒显色后进行胶片曝光,扫描获取目的图像,用Image-ProPlus 6.0软件对目的基因和β-actin免疫印迹结果进行分析。蛋白含量=条带面积×平均灰度;目的蛋白半定量值=目的蛋白含量/β-actin蛋白含量。
表1目的基因引物序列及PCR扩增产物
Table1SequenceofprimerandproductssizeofPCR
1.4 免疫荧光检测
将石蜡组织切片经梯度酒精复水,滴加3% H2O2室温静置(10 min),PBS摇洗3次(每次3 min);滴加5%山羊血清,室温封闭10 min,弃掉切片上的血清,并滴加1∶50倍SOX5兔抗多克隆抗体,阴性对照滴加抗体稀释液;4 ℃过夜;次日37 ℃复温30 min,用PBS摇洗2 次(每次3 min);滴加1∶200倍FITC标记羊抗兔IgG,放入37 ℃温箱孵育30 min,用PBS摇洗4 次(每次5 min);梯度脱水,水溶性封片剂封片、荧光显微镜下观察。
1.5 小鼠SOX5、MITF-M克隆载体和真核表达载体构建
1.5.1 克隆载体构建 首先,Enzyme Solution 5 μL;目的基因PCR胶回收产物4 μL;T-Vector pMD19 1 μL。16 ℃连接14~16 h;随后转化、涂板;次日进行蓝白斑筛选,37 ℃水平摇床200 r·min-1培养12~16 h,提取质粒后,送华大基因公司测序,确定克隆载体是否构建成功。
1.5.2 真核表达载体构建 克隆载体双酶切,回收的目的基因产物5 μL;表达载体产物4 μL;T4 DNA Ligase 1 μL。混匀,PCR仪中16 ℃过夜连接;其余操作同1.5.1。
1.6 黑色素细胞复苏、转染以及黑色素含量测定
小鼠黑色素细胞复苏、培养,待细胞达融合到75%~80%时进行基因转染。首先,将200 μL(无血清双抗)培养基稀释7.5 μL转染试剂,混匀后室温静置5 min。其次,将5.0 μg DNA组加入混合液中,混匀后室温静置15 min。最后,将混合物滴加到含有1 mL新鲜培养基的培养板(6 孔板)中,每孔补至2 mL,37 ℃培养48~56 h。
每组黑色素细胞用0.25%胰酶消化后收集,PBS冲洗2~3次,细胞计数后,用0.2 mol·L-1NaOH溶解黑色素细胞,85 ℃ 5 min充分溶解黑色素颗粒,475 nm波长下进行吸光值检测,每组至少重复3次。
1.7 数据分析
所有数据均用SPSS19.0 软件进行单因素方差分析,结果均用“平均值±标准误(Mean±SE)”表示,所有图柱均用GraphPad PrismTM(GraphPad Software,Inc.California,USA)处理。
2 结 果
2.1 SOX5在不同毛色小鼠皮肤中定位
免疫荧光结果显示,在不同毛色小鼠皮肤中均有SOX5蛋白的表达,且其主要表达部位为毛囊外根鞘,黑色和灰色小鼠皮肤中荧光强度明显高于棕色小鼠皮肤(图1)。由此可以看出,SOX5可以在不同毛色小鼠皮肤定位和表达。

a、b、c.黑、灰、棕小鼠皮肤SOX5阳性组; a-、b-、c-.黑、灰、棕小鼠皮肤SOX5阴性对照组;1.外根鞘;2.内根鞘a, b, c.Black, gray, brown mice skin SOX5 positive groups; a-, b-, c-.Black, gray, brown mice skin SOX5 negative control groups; 1.Outer root sheath; 2. Inner root sheath图1 不同毛色小鼠皮肤组织中 SOX5的免疫荧光图Fig.1 Immunofluorescence results of SOX5 in mouse skin tissues with different coat colors
2.2 SOX5在不同毛色小鼠皮肤中表达
1%琼脂糖凝胶电泳后显示,SOX5条带清晰且单一,条带大小为157 bp (图2A)。切胶后送华大基因公司测序,序列比对后正确,表明SOX5在不同毛色小鼠皮肤中可正常表达。qRT-PCR结果表明:SOX5在黑色和灰色小鼠皮肤中mRNA相对表达量是棕色的8.12倍(P<0.01)和2.87倍(P<0.01)
(图2B)。蛋白免疫印迹结果表明:在不同毛色小鼠皮肤的总蛋白可以与SOX5抗体发生免疫学阳性反应(图2C),通过对蛋白条带分析可知,SOX5在黑色和灰色小鼠皮肤中蛋白水平是棕色的2.46倍(P<0.01)和1.91倍(P<0.01) (图2D)。由此可以看出,SOX5在不同毛色小鼠皮肤中表达量存在差异性,说明SOX5对毛色的形成有一定影响。

**.P<0.01。下同。A.不同毛色小鼠皮肤SOX5 PCR产物,M.DL2000 DNA marker; B.不同毛色小鼠皮肤SOX5的qRT-PCR分析结果;C.SOX5在不同毛色小鼠皮肤的蛋白印迹;D.SOX5在不同毛色小鼠皮肤中蛋白相对水平**.P<0.01. The same as below.A.SOX5 PCR products in different colors of mouse skin; M.DL2000 DNA marker;B. qRT-PCR analysis of SOX5 in different colors of mouse skin; C.Immunoblot results of SOX5 protein in different colors of mouse skin;D.Relative expression level of SOX5 protein in different colors of mouse skin图2 不同毛色小鼠皮肤中SOX5表达量分析Fig.2 Analysis of the expression of SOX5 in different colors of mouse skin
2.3 小鼠SOX5与MITF-M核酸序列获取和真核表达载体构建
在NCBI中获取SOX5和MITF-M的CDS区,成功构建真核表达载体。为确保载体连接的准确性,提取质粒进行了测序。在NCBI对测序结果进行比对,序列为小鼠SOX5和MITF-M的CDS区,大小为2 292和1 260 bp(图略),与SOX5和MITF-M序列完全一致。
2.4 过表达SOX5后黑色素细胞的形态特征
小鼠黑色素细胞复苏、培养,待细胞达融合到75%~80%时,进行基因转染,此时细胞密集且无其它细胞污染。空载组(Vector-GFP)、试验组(Vector-GFP-SOX5)与正常组(Control)相比形态未发生明显改变 (图3)。

Control.正常黑色素细胞;Vector-GFP.转染空载体的黑色素细胞;Vector- GFP-SOX5.转染SOX5的黑色素细胞。下同Control.Normal melanocytes;Vector-GFP.The melanocytes transfected with empty vector;Vector-GFP-SOX5.The melanocytes transfected with SOX5.The same as below 图3 SOX5转染后黑色素细胞的形态Fig.3 Morphology of melanocytes transfected by the SOX5
2.5 黑色素细胞中SOX5的转染效率
过表达SOX5后经检测空载组(Vector-GFP)与正常组(Control)中SOX5表达量无显著差异,试验组(Vector-GFP-SOX5)中SOX5 mRNA升高18.80倍 (P<0.01) (图4A); 蛋白升高1.69倍 (P<0.05)(图4B,C)。
2.6 SOX5与MITF-M表达的相互影响
过表达SOX5后结果显示:与空载组(Vector-GFP)对比,试验组(Vector-GFP-SOX5)的MITF-MmRNA升高1.50倍(P<0.05) (图5A);蛋白升高1.95倍(P<0.01) (图5B,C)。为了进一步说明SOX5与MITF-M的关系,过表达MITF-M后使SOX5的mRNA降低1.34倍(P<0.05)(图5D);蛋白降低2.05倍(P<0.05)(图5E,F)。表明,过表达SOX5促进MITF-M的表达,而过表达MITF-M抑制SOX5的表达。
2.7 SOX5对黑色素细胞色素生成基因及色素含量的影响
过表达SOX5后结果显示:与空载组(Vector-GFP)对比,试验组(Vector-GFP-SOX5)的TYRmRNA升高3.13倍(P<0.01),蛋白升高1.43倍(P<0.05);TYRP1 mRNA升高3.41倍(P<0.01),蛋白升高1.56倍(P<0.05);TYRP2 mRNA升高1.31倍(P<0.05),蛋白升高1.20倍(P<0.05) (图6A,B,C)。通过检测黑色素含量可知空载组(Vector-GFP)与正常组(Control)无显著差异,试验组(Vector-GFP-SOX5)黑色素含量明显增加,相对于空载组黑色素含量增加1.47倍(P<0.01)(图6D)。表明,SOX5表达量的变化对黑色素的合成有影响。
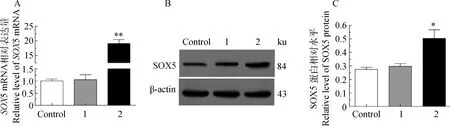
*.P<0.05。1.Vector-GFP;2.Vector-GFP-SOX5。下同。A. SOX5 mRNA的表达差异;B.蛋白阳性免疫印迹;C.蛋白相对水平分析*.P<0.05.1.Vector-GFP;2.Vector-GFP-SOX5.The same as below.A. SOX5 mRNA expression in melanocytes transfected with Vector-GFP-SOX5; B.The positively blotting signal in melanocytes transfected with Vector-GFP-SOX5; C. Relative level of SOX5 protein in melanocytes transfected with Vector-GFP-SOX5图4 SOX 5转染后黑色素细胞中SOX 5 mRNA和蛋白水平Fig.4 SOX5 mRNA and protein levels in melanocytes transfected by the SOX5
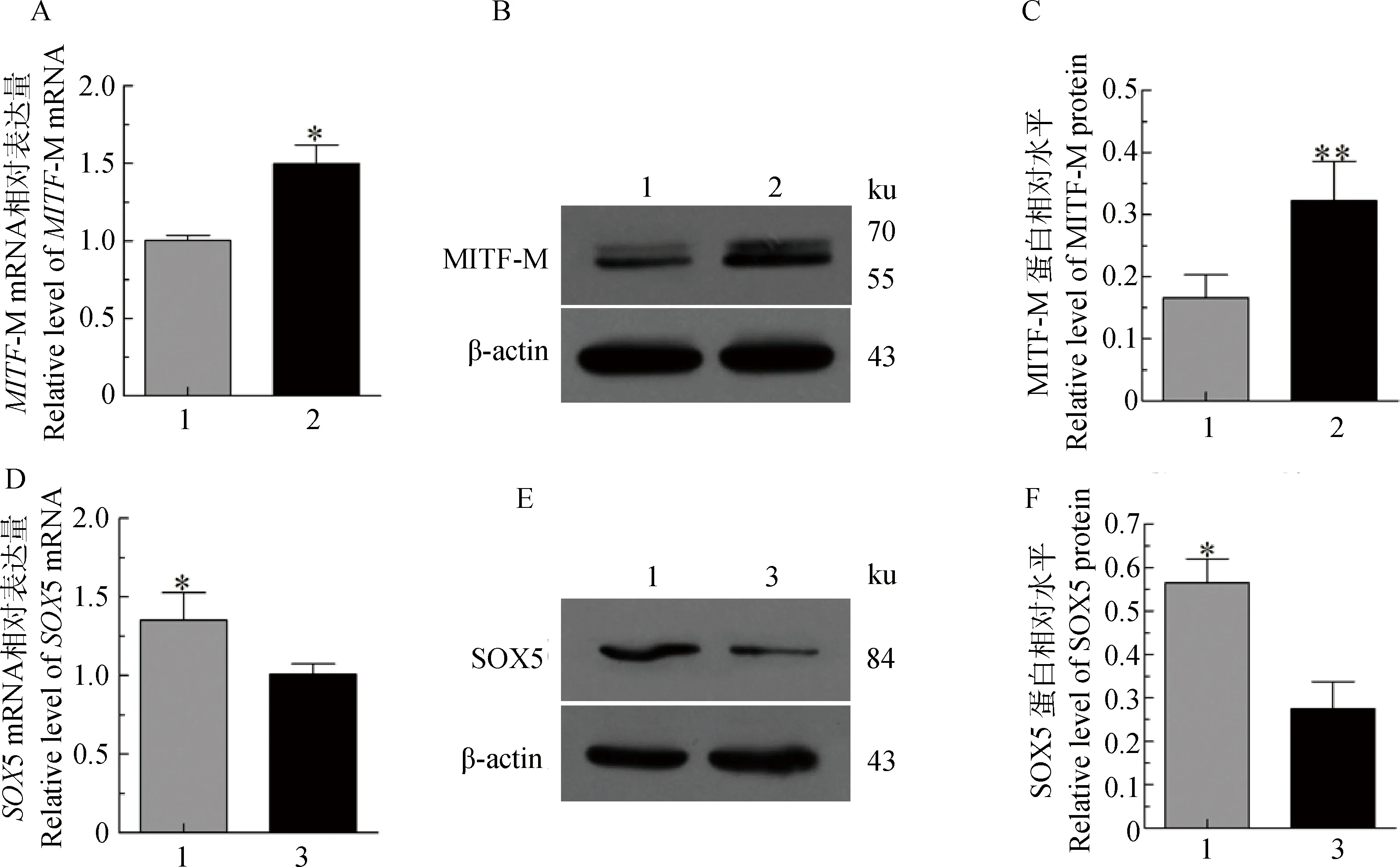
1.Vector-GFP; 2.Vector-GFP-SOX5; 3.Vector-GFP-MITF-M。A、D.MITF-M和SOX5 mRNA的表达差异;B、E.MITF-M和SOX5蛋白阳性免疫印迹;C、F.MITF-M和SOX5蛋白相对水平分析1.Vector-GFP;2.Vector-GFP-SOX5;3.Vector-GFP-MITF-M.A,D.MITF-M and SOX5 mRNA expression in melanocytes transfected with Vector-GFP-SOX5 and Vector-GFP-MITF-M; B,E.The positively blotting signal in melanocytes transfected with Vector-GFP-SOX5 and GFP-MITF-M; C,F.Relative level of MITF-M and SOX5 proteins in melanocytes transfected with Vector-GFP-SOX5 and Vector-GFP-MITF-M图5 检测转染后黑色素细胞中SOX5和MITF-M的表达Fig.5 Analysis of SOX5 and MITF-M expression in melanocytes after transfection
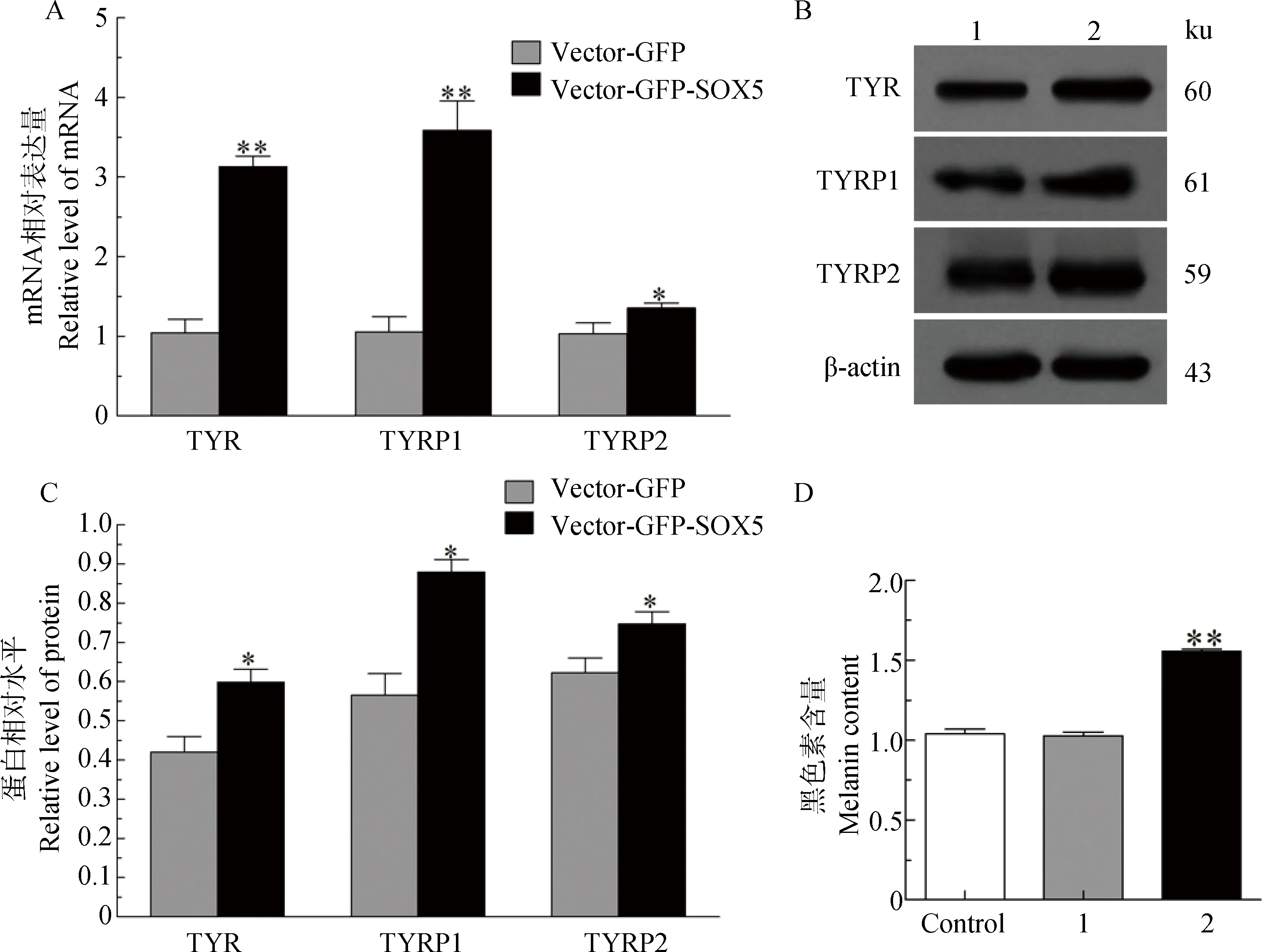
A.TYR、TYRP1、TYRP2 mRNA的表达差异; B.蛋白阳性免疫印迹; C.TYR、TYRP1和TYRP2蛋白相对水平分析; D.黑色素含量分析A.TYR,TYRP1 and TYRP2 mRNA expression in melanocytes transfected with Vector-GFP-SOX5; B.The positively blotting signal in melanocytes transfected with Vector-GFP-SOX5; C.Relative levels of TYR,TYRP1 and TYRP2 protein in melanocytes transfected with Vector-GFP-SOX5; D.Melanin content analysis in Control, Vector-GFP and Vector-GFP-SOX5图6 SOX5对黑色素生成的影响Fig.6 Effect of SOX5 on melanin formation
2.8 SOX5对PMEL和OA1的影响
过表达SOX5后结果显示:与空载组(Vector-GFP)对比,试验组(Vector-GFP-SOX5)的PMELmRNA升高4.8倍(P<0.01) (图7A)(PMEL不属于主线基因加上经费原因,没有进行相关蛋白分析);OA1 mRNA升高2.1倍(P<0.05),OA1蛋白升高1.29倍(P<0.05) (图7B,C,D)。由此可以看出,过表达SOX5可以使PMEL和OA1的表达升高。
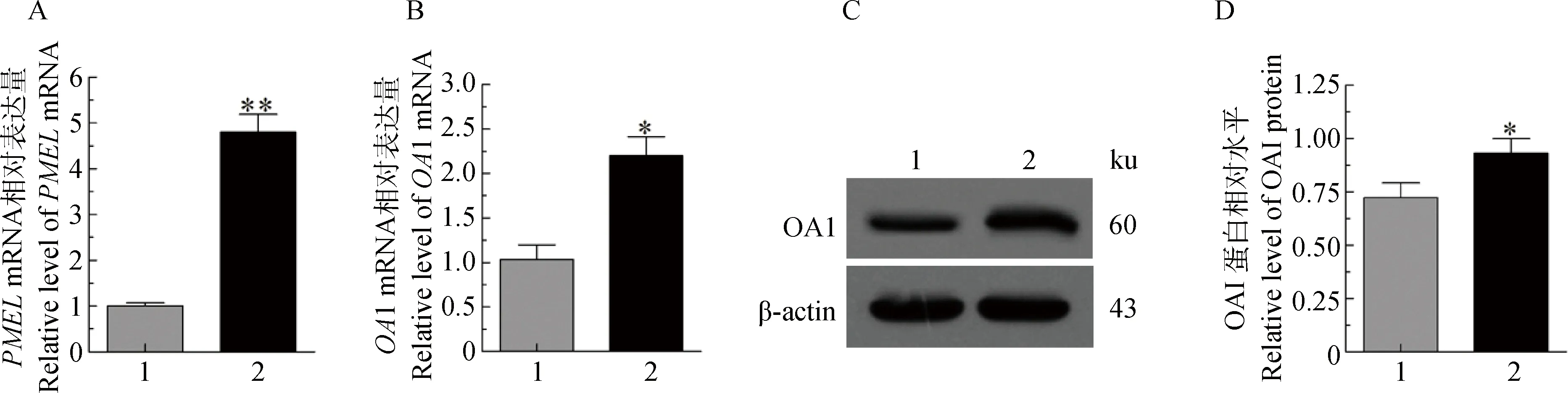
A、B. PMEL和OA1 mRNA的表达差异; C.OA1蛋白阳性免疫印迹; D. OA1蛋白相对水平分析A,B.PMEL and OA1 mRNA expression in melanocytes transfected with Vector-GFP-SOX5;C.The positively blotting signal of OA1 in melanocytes transfected with Vector-GFP-SOX5;D.Relative level of OA1 protein in melanocytes transfected with Vector-GFP-SOX5图7 SOX5对PMEL和OA1的影响Fig.7 Effect of SOX5 on PMEL and OA1
3 讨 论
哺乳动物背毛颜色的多样性取决于黑色素中的真黑素(棕色/黑色)和褐黑素(黄色/红色)的相对含量[25]。P.C.Wang等[26]在黑色素细胞中发现microRNA-21a-5p在调节黑色素生成的功能中是通过靶向SOX5实现的。以及在人的黑色素瘤细胞中,SOX5通过MITF调节人黑素瘤细胞的平衡。本试验探究SOX5在不同毛色小鼠皮肤表达量是否存在差异性,结果表明,SOX5在不同毛色小鼠皮肤表达量不同,SOX5在黑色小鼠皮肤的表达量>灰色>棕色。本试验结果为进一步研究其对黑色素的生成以及如何参与毛色形成奠定了基础。
本试验通过过表达SOX5后明显升高MITF-M的表达。与此不同的是T.Kordab等[6]发现的升高SOX5抑制了MITF的表达。两者存在差异性可能由于SOX5过高或过低激活相应的调控路径对MITF-M表达产生不同的结果。为了进一步了解SOX5与MITF-M关系,结果显示过表达MITF-M对SOX5表达存在负反馈作用。由此可以看出,SOX5可以调控MITF-M表达进而影响色素生成。黑素体是黑色素合成的场所[27],黑素体蛋白PMEL发生在黑素体I期和Ⅱ期作为原纤维的结构基础,在斑马鱼视网膜色素上皮细胞PMEL和OA1两者均可调节黑素体数量、形状以及能动性[28],并且过表达OA1可以明显升高PMEL[22]。本试验过表达SOX5后,PMEL和 OA1表达量升高,由此可知,PMEL与 OA1受SOX5通过MITF-M来调节。本试验通过过表达SOX5,使TYR、TYRP1以及TYRP2表达量升高,最终引起黑色素含量升高;与杨玉静等[29]研究相吻合。由此可以得出,SOX5可以影响色素生成,进而参与毛色的形成。
4 结 论
SOX5在不同毛色小鼠皮肤中的表达存在差异性,可以通过MITF-M调控色素相关基因来影响色素生成,并且MITF-M对SOX5有负调控作用,这为SOX5参与毛色形成潜在机制提供依据。
[1] 储沨婷. SOX蛋白在软骨内成骨中的作用[J]. 国外医学: 口腔医学分册, 2006, 33(2): 136-138.
CHU F T. SOX protein in cartilage osteogenesis in the role of[J].ForeignMedicine:JournalofStomatology, 2006, 33(2): 136-138. (in Chinese)
[2] LEFEBVRE V. The SOXD transcription factors-SOX5, SOX6, and SOX13-are key cell fate modulators[J].IntJBiochemCellBiol, 2010, 42(3): 429-432.
[3] SMITS P, DY P, MITRA S, et al.SOX5 andSOX6 are needed to develop and maintain source, columnar, and hypertrophic chondrocytes in the cartilage growth plate[J].JCellBiol, 2004, 164(5): 747-758.
[4] MARTINEZ-MORALES P L, QUIROGA A C, BARBAS J A, et al. SOX5 controls cell cycle progression in neural progenitors by interfering with the WNT-β-catenin pathway[J].EMBORep, 2010, 11(6): 466-472.
[5] STOLT C C, LOMMES P, HILLGRTNER S, et al. The transcription factor SOX5 modulates SOX10 function during melanocyte development[J].NucleicAcidsRes, 2008, 36(17): 5427-5440.
[6] KORDAΒ T, WEBER C E M, OSWALD M, et al. SOX5 is involved in balancedMITFregulation in human melanoma cells[J].BMCMedGenomics, 2016, 9: 10.
[7] HEMESATH T J, STEINGRMSSON E, MCGILL G, et al. microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family[J].GenesDev, 1994, 8(22): 2770-2780.
[8] LEVY C, KHALED M, FISHER D E. MITF: master regulator of melanocyte development and melanoma oncogene[J].TrendsMolMed, 2006, 12(9): 406-414.
[9] HERSHEY C L, FISHER D E. Genomic analysis of theMicrophthalmialocus and identification of the MITF-J/Mitf-J isoform[J].Gene, 2005, 347(1): 73-82.
[10] FUSE N, YASUMOTO K, SUZUKI H, et al. Identification of a melanocyte-type promoter of the microphthalmia-associated transcription factor gene[J].BiochemBiophysResCommun, 1996, 219(3): 702-707.
[11] ROH E, YUN C Y, YUN J Y, et al. cAMP-binding site of PKA as a molecular target of bisabolangelone against melanocyte-specific hyperpigmented disorder[J].JInvestDermatol, 2013, 133(4): 1072-1079.
[12] HARTMAN M L, CZYZ M. Pro-survival role of MITF in melanoma[J].JInvestDermatol, 2015, 135(2): 352-358.
[13] WELLBROCK C, AROZARENA I. Microphthalmia-associated transcription factor in melanoma development and MAP-kinase pathway targeted therapy[J].PigmentCellMelanomaRes, 2015, 28(4): 390-406.
[14] YASUMOTO K, YOKOYAMA K, TAKAHASHI K, et al. Functional analysis of microphthalmia-associated transcription factor in pigment cell-specific transcription of the human tyrosinase family genes[J].JBiolChem, 1997, 272(1): 503-509.
[15] FANG D, TSUJI Y, SETALURI V. Selective down-regulation of tyrosinase family geneTYRP1 by inhibition of the activity of melanocyte transcription factor, MITF[J].NucleicAcidsRes, 2002, 30(14): 3096-3106.
[16] DU J Y, MILLER A J, WIDLUND H R, et al. MLANA/MART1 and SILV/PMEL17/GP100 are transcriptionally regulated by MITF in melanocytes and melanoma[J].AmJPathol, 2003, 163(1): 333-343.
[17] MCKAY B S, SCHWARTZ S G. Pigmentation and macular degeneration: is there a role for GPR143?[J].JOculPharmacolTher, 2016, 32(1): 3-4.
[18] GIORDANO F, SIMOES S, RAPOSO G. The ocular albinism type 1 (OA1) GPCR is ubiquitinated and its traffic requires endosomal sorting complex responsible for transport (ESCRT) function[J].ProcNatlAcadSciUSA, 2011, 108(29): 11906-11911.
[19] BENTLEY N J, EISEN T, GODING C R. Melanocyte-specific expression of the human tyrosinase promoter: activation by the microphthalmia gene product and role of the initiator[J].MolCellBiol, 1994, 14(12): 7996-8006.
[20] BAXTER L L, PAVAN W J.Pmel17 expression isMitf-dependent and reveals cranial melanoblast migration during murine development[J].GeneExprPatterns, 2003, 3(6): 703-707.
[21] WATT B, VAN NIEL G, RAPOSO G, et al. PMEL: a pigment cell-specific model for functional amyloid formation[J].PigmentCellMelanomaRes, 2013, 26(3): 300-315.
[22] CHEN T Z, WANG H D, LIU Y, et al.OcularAlbinismType1 regulates melanogenesis in mouse melanocytes[J].IntJMolSci, 2016, 17(10): 1596.
[23] FALLETTA P, BAGNATO P, BONO M, et al. Melanosome-autonomous regulation of size and number: the OA1 receptor sustains PMEL expression[J].PigmentCellMelanomaRes, 2014, 27(4): 565-579.
[25] THODY A J, HIGGINS E M, WAKAMATSU K, et al. Pheomelanin as well as eumelanin is present in human epidermis[J].JInvestDermatol, 1991, 97(2): 340-344.
[26] WANG P C, ZHAO Y Y, FAN R W, et al. microRNA-21a-5p functions on the regulation of melanogenesis by targeting Sox5 in mouse skin melanocytes[J].IntJMolSci, 2016, 17(7): 959.
[27] GERSTENBLITH M R, SHI J X, LANDI M T. Genome-wide association studies of pigmentation and skin cancer: a review and meta-analysis[J].PigmentCellMelanomaRes, 2010, 23(5): 587-606.
[28] BURGOYNE T, O′CONNOR M N, SEABRA M C, et al. Regulation of melanosome number, shape and movement in the zebrafish retinal pigment epithelium by OA1 and PMEL[J].JCellSci, 2015, 128(7): 1400-1407.
[29] 杨玉静, 张丹瑾, 聂瑞强, 等. 绵羊MITF-M在黑素细胞中过表达后的功能分析[J]. 中国农业科学, 2016, 49(21): 4214-4221.
YANG Y J, ZHANG D J, NIE R Q, et al. The function analysis of over-expression of oar MITF-M in melanocytes[J].ScientiaAgriculturaSinica, 2016, 49(21): 4214-4221. (in Chinese)
