山羊DCT基因启动子活性区及其转录因子调控探究
2018-01-26刘春杨张乐超周荣艳李兰会李祥龙
刘春杨,张乐超,王 麒,周荣艳,李兰会*,李祥龙
(1.河北农业大学动物科技学院,保定 071000; 2.河北科技师范学院,秦皇岛 066004)
定位于黑素体膜上的跨膜蛋白—多巴色素异构酶 (Dopachrometautomerase,DCT),也称作酪氨酸酶相关蛋白2 (TYRP2),参与黑色素的生物合成过程[1-2],1992 年被确认为酪氨酸酶相关蛋白家族的第3个成员,具有多巴色素异构酶(DT)活性,催化多巴色素转变为5, 6-二羟基吲哚羧酸( DHICA),具有加速黑色素生成的作用。黑色素细胞中的DCT基因控制着5, 6-二羟基吲哚羧酸与5,6-二羟基吲哚(DHI)的比例,因此推测,DCT可能对黑色素合成早期阶段的酪氨酸酶催化活性有调节作用,对动物毛色形成发挥重要作用[3-4]。而且,由于中间产物DHI在体内比DHICA有更高的细胞毒性,DCT能使含有羧酸前体的DHICA迅速掺入生物合成的黑色素内,对减少中间产物的细胞毒性有重要意义[5-6]。据前人研究,DCT基因单碱基变异的小鼠,与野生型黑色小鼠相比,DCT活性降低了3~4倍,从而产生了深灰或深棕色小鼠[7]。DCT基因突变的小鼠黑色素细胞过早死亡,可能是由于 DCT 缺失而产生的细胞毒性中间体所致[8]。也有研究发现,小鼠敲除DCT第一外显子后导致DCT基因mRNA和蛋白质表达量缺失,影响了黑色素细胞的活性和色素沉着,形成深灰色被毛[9]。
鉴于DCT基因表达在动物毛色和生命活动中的重要作用,其表达水平受转录因子调控[10-11],因此,研究DCT基因启动子转录调控机理对于揭示动物毛色形成具有重要作用。SOX10是调控DCT基因表达的一个非常重要的转录因子[12],体外试验表明,SOX10能够激活DCT基因启动转录[13-14]。MITF也是黑色素合成过程中重要的转录因子,有研究认为,DCT基因的表达是依靠MITF与SOX10两种转录因子协同激活的[15-16],也有研究认为,DCT可能存在不依赖于MITF的转录调控[17-19]。人视网膜色素上皮细胞的体外试验发现,转录因子OTX2能够结合到DCT基因的启动子区,抑制内源OTX2的表达导致DCT蛋白含量的减少,说明OTX2能够激活DCT基因启动子[20]。
本试验克隆山羊DCT基因5′UTR区序列,并构建DCT基因系列缺失片段的pGL3-Basic重组质粒,利用点突变方法构建转录因子SOX10、MITF和OTX2结合位点突变的DCT基因系列重组质粒,以Luciferase报告基因系统检测重组质粒的启动活性变化,明确该基因启动子核心调控区和转录因子对DCT基因的表达调控,以期为山羊DCT基因的表达调控提供理论依据。
1 材料与方法
1.1 试验材料
pGL3-Basic 与 pRL-TK 载体、双荧光素酶报告基因检测试剂盒为美国 Promega 公司产品;脂质体Lipofectamine®2000 Reagent为美国Invitrogen公司产品;Opti-MEM培养基为美国Gibco公司产品;胎牛血清、DMEM培养基为美国Hyclone公司产品;TransStartTaqDNA 聚合酶、PCR产物纯化试剂盒、点突变试剂盒为北京全式金公司产品;感受态细胞DH5α、胶回收试剂盒为上海生工生物工程有限公司产品;闪电克隆试剂盒为北京博奥龙免疫技术有限公司产品;内切酶为大连宝生物工程有限公司产品;无内毒素质粒提取试剂盒为北京天根公司产品。引物合成由上海生工生物工程有限公司完成,测序由北京六合华大基因公司完成。
绩效管理工作的有效推进可以很大程度上激发工作人员的工作积极性,并保证全部管理活动能够在绩效管理体系建设的过程中实现更大的价值。
1.2 试验方法
1.2.4 重组质粒的构建 将回收的山羊DCT基因系列缺失片段与线性化载体进行连接重组,反应体系:目的片段400 ng,载体20 ng,2×Lightening Cloning Master Mix 5 μL,ddH2O补足至10 μL,50 ℃水浴1 h。将反应液转化到DH5α感受态细胞,鉴定为阳性的单菌落送公司测序,确定为阳性克隆,进一步进行重组质粒的大量提取。
根据点突变试剂盒说明书,应用PCR方法实现碱基突变。PCR反应体系:原质粒载体DNA 20~50 ng,上下游引物(10 μmol·L-1)各1 μL,2×TransStart®FastPfu PCR SuperMix 25 μL,ddH2O补足至50 μL。反应程序:94 ℃预变性5 min;94 ℃变性20 s,退火温度退火20 s,72 ℃延伸2 min,共35个循环;72 ℃终延伸10 min;4 ℃终止反应。1%琼脂糖凝胶电泳检测,若有目的条带,加1 μL DMT酶于10 μL PCR产物中,37 ℃消化孵育1 h。将消化产物转化到50 μL DMT感受态细胞中,涂布到含氨苄的培养皿,挑取单菌落,送至华大测序。成功实现突变的载体进行质粒的大量提取,为转染做准备。
利用特异引物(表1)扩增山羊DCT基因上游序列5′端缺失片段,分别命名为P1、P2、P3、P4和P5。根据第一轮启动子预测片段活性值,以P3片段序列为模板,设计一系列3′端缺失序列引物,分别命名为P6、P7、P8、P9、P10和P11。
表1山羊DCT基因启动子扩增引物
Table1PrimersamplifyinggoatDCTgenepromoterswithdifferentlength
小写字母为与载体的重叠区序列;下划线标记为所用酶切位点;大写字母为引物特异性序列
Lowercase letters are overlap regions with the vectors; underlined letters are restriction enzyme digestion sites; uppercase letters are the specifical primer sequences
1.2.2 山羊DCT基因启动子系列缺失片段的克隆 PCR反应体系:10×PCR Buffer 5 μL,dNTP(2.5 mmol·L-1)4 μL,上、下游引物(10 μmol·L-1)各1 μL,DNA模板2.5 μL,TransStartTaq酶(2.5 U·μL-1)0.5 μL,最后ddH2O补足至50 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,退火温度退火30 s,72 ℃延伸0.5~2 min,共35个循环;72 ℃终延伸10 min;4 ℃终止反应。利用1%琼脂糖凝胶对 PCR 产物进行电泳检测,若条带单一且为目的条带,直接进行PCR产物纯化,若条带复杂但含有目的条带,将PCR产物进行切胶纯化回收处理。
1.2.3 载体质粒的线性化 根据设计引物时所加酶切位点,利用SacI和XhoI两种限制性内切酶将载体pGL3-Basic进行双酶切处理,实现载体的线性化。载体双酶切体系:载体质粒1 500 ng,10×M Buffer 6 μL,SacI和XhoI各1.5 μL,H2O补足至60 μL,37 ℃反应3~4 h,将反应液进行切胶回收。
基于对青少年自我价值感及道德判断能力与价值观关系的研究,在自我价值感中,自我观与人际情感之间具有正向关系,其他则为负向。换言之,自我观越重的学生,其自我价值感越强,并以此更加关注自身的成长,形成良性循环。与此同时,群体观与自我价值感却存在着互相关联,自我观强导致群体观低、人际价值感高。
1.2.1DCT基因启动子系列缺失片段引物设计 以山羊DCT基因序列(NCBI收录号:NC_030819.1)为模板,利用在线引物设计程序NEBuilder(http://nebuilder.neb.com/)设计特异引物,在上游引物5′端引入SacI的酶切位点及部分与表达载体的重叠区,下游引物5′端引入XhoI的酶切位点及部分与表达载体的重叠区。
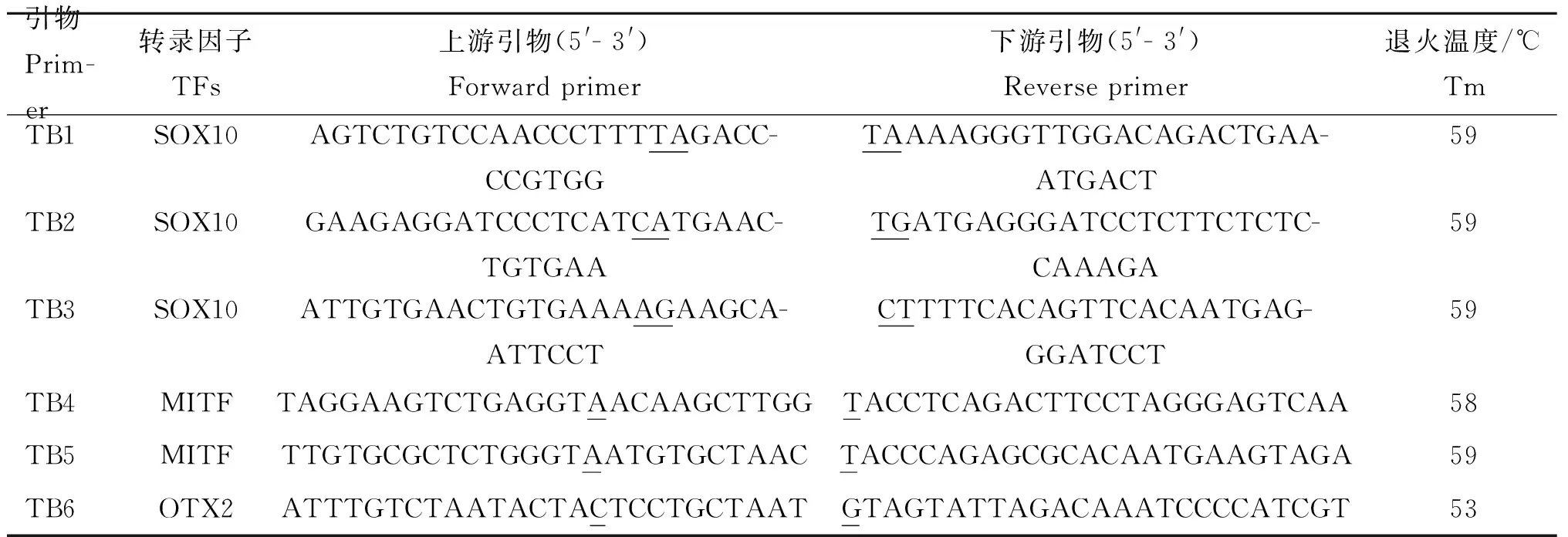
1.2.5 转录因子结合位点点突变载体的构建 利用转录因子在线分析软件JASPAR(http://jaspar.genereg.net/cgi-bin/jaspar_db.pl)对启动子活性值最高的片段进行转录因子结合位点分析。根据不改变DCT基因其它转录因子结合位点的原则,按照点突变说明书,对启动子活性值最高片段设计突变转录因子SOX10、MITF和OTX2结合位点的引物,命名为TB1、TB2、TB3、TB4、TB5和TB6(表2)。TB1将-846和-845 bp位置GT突变为TA,TB2将-617 和-616 bp位置GT突变为CA,而TB3将-604和-603 bp位置CA突变为AG,从而使三者的SOX10结合位点消失。TB4将-410 bp的C突变为A,TB5将-310 bp的C突变为A,使MITF结合位点消失。TB6将-277 bp的A突变成C,使转录因子OTX2结合位点消失。
表2转录因子结合位点突变引物
Table2Primersofpointmutationintranscriptionfactorbindingsite
引物Prim-er转录因子TFs上游引物(5'-3')Forwardprimer下游引物(5'-3')Reverseprimer退火温度/℃TmTB1SOX10AGTCTGTCCAACCCTTTTAGACC-CCGTGGTAAAAGGGTTGGACAGACTGAA-ATGACT59TB2SOX10GAAGAGGATCCCTCATCATGAAC-TGTGAATGATGAGGGATCCTCTTCTCTC-CAAAGA59TB3SOX10ATTGTGAACTGTGAAAAGAAGCA-ATTCCTCTTTTCACAGTTCACAATGAG-GGATCCT59TB4MITFTAGGAAGTCTGAGGTAACAAGCTTGGTACCTCAGACTTCCTAGGGAGTCAA58TB5MITFTTGTGCGCTCTGGGTAATGTGCTAACTACCCAGAGCGCACAATGAAGTAGA59TB6OTX2ATTTGTCTAATACTACTCCTGCTAATGTAGTATTAGACAAATCCCCATCGT53
下划线处为突变碱基
Underlined letters are mutant bases
从“宏大叙事”转向“微小叙事”—从英雄、模范的典型事迹转向平民百姓的生活事件;从隐身叙事转到自传叙事—从讲述别人的故事到自己“亲历”的故事;从“社会剧本”转到“生活剧本”—把学生当下身边发生的道德事件作为教材的出发点,发现细微小事和典型事例中隐藏的道德智慧。
风险投资在近年“共享经济”的飞速发展中被人所熟知,是投资中介向特别有潜能的高新技术产业投入风险资本后,实现利益共享、风险共担的一种投资方式。大学生创业遇到的最大问题就是资金不足。以Dormi为例,在校创业阶段,有政府提供的项目资金与学校举办相关活动的奖金。在转型进入更大市场时,依靠前期盈利与创始团队家庭支持是不够的,必须寻求社会资金的帮助。对于风险投资公司来说,他们也希望寻找到有发展潜力的中小型项目,通过早期“资本换股权”,从而在未来公司成熟后以更高价格将股权卖出获得收益[2]。
颠覆潮流,打破“长”规,全新梅赛德斯-奔驰长轴距A级轿车以实力向世人揭示—这才是新生代豪华轿车该有的样子!此次上市车型包括全新A 180 L轿车及运动轿车、全新A 200 L轿车及运动轿车,以及全新A 200 L运动轿车先型特别版共 5款车型,厂商建议零售价格区间为人民币21.69万元至29.99万元。
2 结 果
2.1 山羊DCT基因启动子区调控元件分析
NCBI数据库中下载山羊DCT基因(NC_030819.1),并与人和鼠DCT基因启动子序列相比,山羊DCT基因的-648~+73 bp(外显子1第1个碱基记为+1)区域与人的相似度达81%,-472~-28 bp区域与鼠DCT基因启动子序列相似度达78%。
对目的序列进行启动子特征分析,发现多种调控元件(图1):包含6个TATA box,分别位于-382~-366 bp、-337~-321 bp、-205~-189 bp、-187~-172 bp、-125~-109 bp和+774~+790 bp;位于-138~-123 bp的GC box;位于-188~-175 bp 的CAAT box;位于-313~-303 bp的M box和位于-620~-588 bp 的32 bp元件。

图1 山羊DCT基因序列特征图Fig.1 Sequence feature of goat DCT gene
利用在线程序Meth Primer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi) 对山羊DCT基因-2 000~+1 000 bp序列进行CpG岛预测分析,参数设置为Island size ≥100,Obs/Exp ≥0.6,GC Percent ≥50.0。在3 000 bp的序列中,共存在2处CpG岛,分别位于该基因的-93~+7 bp和+309~+603 bp(图1)。
1.2.6 山羊DCT基因启动子活性检测 A375细胞接种于含10%胎牛血清的DMEM培养基,在 5% CO2和 37 ℃条件下培养。转染前1 d以每孔1.5×105个细胞的密度接种于24孔培养板,以不含血清及抗生素的培养基进行培养。当细胞融合度达到90%,根据Lipofectamine®2000 Reagent脂质体转染说明开始转染。脂质体与质粒总量比例为2∶1,pGL3-promoter质粒与pRL-TK质粒共转染,转染比例为19∶1。转染后 48 h 裂解细胞,利用双荧光素酶检测试剂盒,GloMax-MultiJr 单管型多功能检测仪检测荧光素酶活性。
利用Promoter 2.0预测山羊DCT基因-801、-501和+200 bp 3个位置为启动子位置,分值分别为0.657、0.692和0.621。
2.2 山羊DCT基因启动子缺失片段载体的构建
5′和3′系列缺失序列片段的PCR产物1%琼脂糖凝胶电泳检测见图2,可见目的条带单一,亮度清晰,无杂带。图2中P1、P2、P3、P4和P5片段分别长2 059、1 598、1 222、775和436 bp,P6、P7、P8、P9、P10和P11片段分别长1 024、778、728、690、650和356 bp。
将PCR产物纯化回收,与线性化表达载体混合,加入重组酶,使目的片段与线性化表达载体连接,构建重组质粒,阳性克隆送华大公司测序,测序结果与预期结果一致,证明重组载体构建成功。
(4)借助社会专业力量,对企业进行流程进行梳理,提升核心竞争力。国家有一整套完善的《企业内部控制规范》属理论范畴,指引企业内部风险控制。但是,企业内控成本与企业实际工作的有效结合,成本效益原则的尺度不好掌握。往往造成的结果是高成本全覆盖,有事大家都有责,没人担当,没有责任人,也没有经济成果,无为就无责。企业应以实际经营工作及流程为出发点,聘请社会专业力量,对企业流程进行梳理,缩短管理链条,根据重要性原则和关键节点实施有效控制,有法满足的实施替代内控复核程序,提升管理效率,找到成本与效益的最佳结合点,实现国有企业的价值最大化。
2.3 重组质粒启动子活性分析
虽然P9片段的基本调控元件与P10片段相同,但P9却表现出极显著高于P10的启动子活性(P<0.01),说明-232~-192 bp间存在重要调控元件,1个TATA box(-205~-189 bp),增加DCT基因的转录活性。P8片段增加1个TATA box(-187~-172 bp)和1个CAAT框(-188~-175 bp),其活性表现为最高,比阴性对照高出20.26倍,极显著高于其它所有片段(P<0.01),推测-192~-154 bp间的CAAT box为DCT基因的关键调控元件,发挥增强子作用。

M. DNA 相对分子质量标准;P1~P11. 系列缺失片段M. DNA marker; P1-P11. The series of deleted fragments图2 DCT基因系列缺失片段PCR产物凝胶检测图Fig.2 Agarose gel electropherogram of PCR products of the serial deleted fragments of DCT gene
不同片段显示出不同的荧光活性,说明启动子不同区域存在不同作用的重要调控元件。由P5、P4至P3片段长度增加,启动子活性逐渐增加推测,山羊DCT基因核心启动子区可能位于-990~-204 bp范围内,与Promoter 2.0预测的-801和-501 bp启动子位置结论一致;-204~+232 bp区域内可能存在负调控元件,与该区域内的CpG岛(-93~+7 bp)和GC box(-138~-123 bp)有关,或正调控元件不能发挥启动作用。
1.2.7 数据分析 利用SPSS 17.0软件的ANOVA对系列缺失片段活性进行分析,采用LSD多重比较对每个检测片段的启动子活性作两两比较。对点突变载体利用独立样本t检验比较突变前后活性变化。P<0.01表示差异极显著。
由验证结果可知,所有的观测变量与其对应的具体因素之间都具有显著性关系。再通过表4可发现,智慧城市感知质量、感知价值以及发展水平都与智慧城市建设市民满意度呈显著正相关;智慧城市市民抱怨、智慧城市预期与智慧城市建设市民满意度负相关是显著的;智慧城市感知价值、智慧城市发展水平、智慧城市预期都与智慧城市感知质量正相关是显著的;智慧城市感知价值、智慧城市发展水平与智慧城市预期呈正相关且是显著的;智慧城市感知价值、智慧城市发展水平与智慧城市市民抱怨呈负相关且是显著的;智慧城市预期与智慧城市市民抱怨呈正相关但不显著。在图2中,用实线代表该影响路径具有显著性,用虚线代表该影响路径不具有显著性。
以-990~+232 bp的P3片段序列为模板,构建系列3′端缺失序列报告基因载体P6~P11,5′端固定在-881 bp位置,3′端依次缩短。如图1所示,片段P6(-881~+143 bp)包含32 bp元件、M box、CAAT box、GC box、CpG岛和5个TATA box;片段P7(-881~-104 bp)在P6基础上缺少CpG岛;P8(-881~-154 bp)在P7基础上缺少GC box和TATA box;P9(-881~-192 bp)在P8基础上缺少1个CAAT box和2个TATA box;P10(-881~-232 bp)和P9所包含调控元件相同;片段P11(-881~-526 bp)只包含32 bp元件。
图3显示,片段P6~P10活性值均极显著高于阴性对照组(P<0.01),P11活性值与阴性对照差异不显著(P>0.01),说明包含预测启动子位置-801 bp的P11片段仅有32 bp元件并不能启动DCT基因的表达,必须与其他调控元件结合共同发挥启动活性。P10片段增加了2个TATA box和1个M box,其启动活性极显著高于P11(P<0.01)。这不仅验证了5′系列缺失片段活性检测结果,并进一步精确核心启动子区为-881~-232 bp区,说明包含2个预测启动子位置-801 和-501 bp的P10,具有32 bp元件(-620~-588 bp)与2个TATA box(-382~-366 bp与-337~-321 bp)和1个M box(-313~-303 bp)组合发挥了基本转录活性。
表达载体pGL3-Basic与5′系列缺失片段重组,获得报告基因载体pGL3-P1~pGL3-P5,瞬时转染A375细胞,细胞状态良好,转染后48 h收获并裂解细胞,检测萤火虫荧光素酶荧光值(F)和海肾荧光素酶荧光值(R),计算相对荧光素酶活性,即F/R值。结果显示(图3),P1(-1 827~+232)、P2(-1 366~+232)、P3(-990~+232)和P4(-543~+232)片段活性均极显著高于阴性对照组(P<0.01),P3活性极显著高于其它片段(P< 0.01),但P5(-204~+232)活性与阴性对照差异不显著(P>0.01)。
片段P7和P6增加了GC box(-138~-123 bp)、CpG岛(-93~+7 bp)和1个TATA box,其活性极显著低于P8和P9(P<0.01),说明-154~+143 bp区域存在的GC box和CpG岛发挥了负调控作用,与5′系列缺失片段发现的-204~+232 bp的负调控结果一致。
2.4 转录因子结合位点点突变报告基因载体构建及活性分析
以P8报告基因载体质粒为模板,使用点突变试剂盒构建点突变报告基因片段TB1~TB6,PCR产物为环状,大小为5 525 bp,琼脂糖凝胶电泳检测结果如图4所示。
课题组设计调查表,对各地市食药检机构的人员数量、编制、职称、学历情况进行调查,于2015年8月5日通过广西食品药品监督管理局发放给8家地市级食品药品检验检测机构填报,2015年9月5日前收回问卷。本次调查共发放问卷8份,回收8份,由课题组对数据进行核实和汇总。

不同大写字母代表差异极显著(P<0.01),相同大写字母代表差异不显著(P>0.01)Columns with different upper case letters mean highly significant difference (P<0.01); Columns with the same upper case letter mean no significant difference (P>0.01)图3 系列缺失片段荧光素酶相对活性值Fig.3 Luciferase relative activity of serial deleted fragments
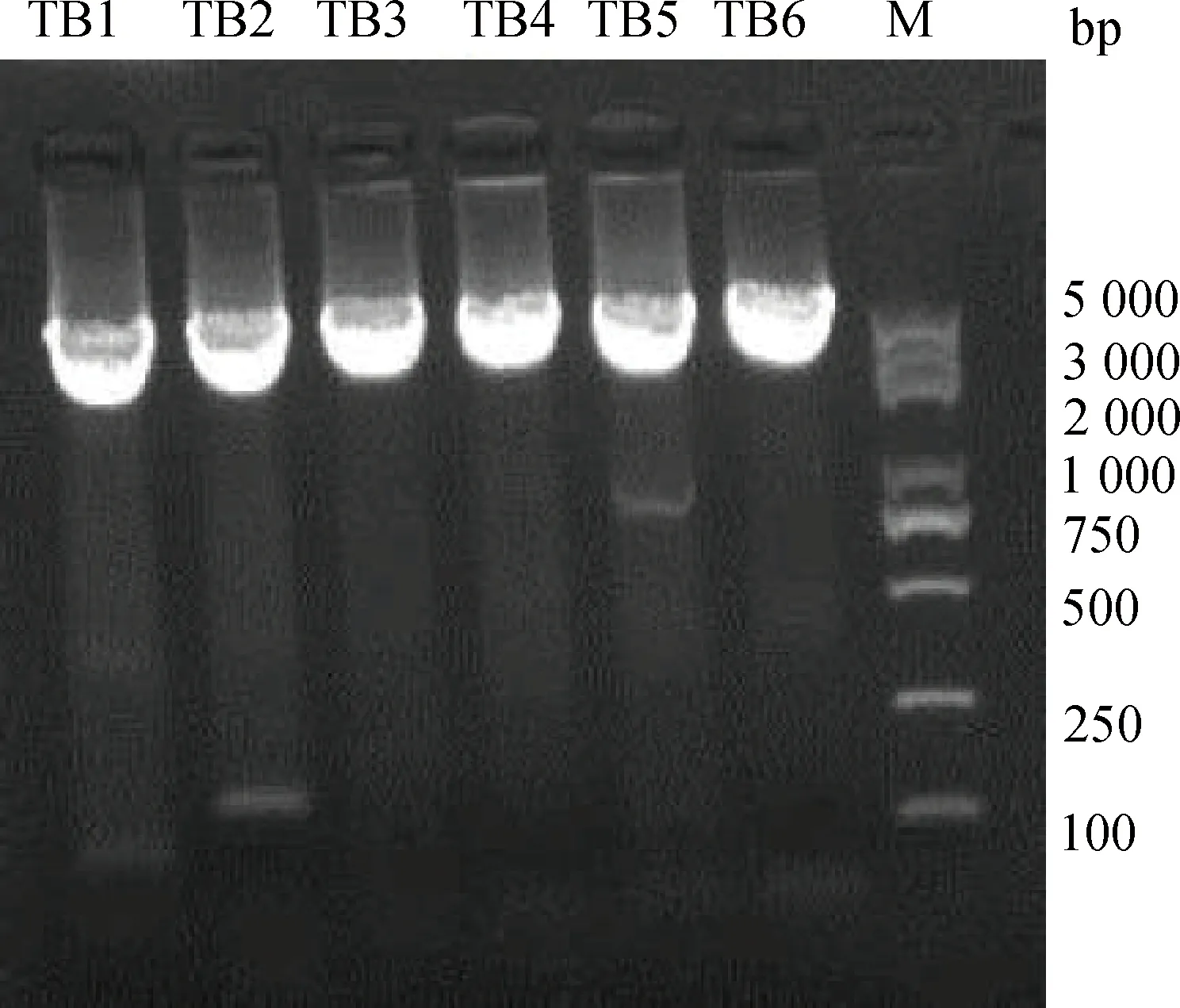
TB1~TB6. 转录因子结合位点点突变报告基因片段;M. DNA 相对分子质量标准TB1-TB6. Reporter gene fragments with point mutation of transcription factor binding site; M. DNA marker 图4 转录因子结合位点点突变片段扩增结果Fig.4 Amplification result of point mutation fragments for transcription factor binding sites
图5显示,使SOX10结合位点消失的突变载体TB1(-846和-845 bp)、TB2(-617和-616 bp)和TB3(-604和-603 bp)活性较突变前活性极显著降低(P<0.01),推测转录因子SOX10与DCT基因在核心启动子区域的结合对转录表达起到正调控作用。而引起MITF和OTX2结合位点消失的突变载体TB4(-410 bp)、TB5(-310 bp)和TB6(-277 bp)启动子活性较突变前极显著增强(P<0.01),推测转录因子MITF和OTX2对山羊DCT基因的转录表达起到负调控的作用或DCT基因存在不依赖于MITF和OTX2的其他转录调控机制。
3 讨 论
3.1 利用山羊DCT基因5′和3′系列缺失片段活性检测核心启动子区
启动子的检测方法有多种,最常见的就是报告基因的方法,基本方式就是将生物信息学预测与试验相结合,根据预测结果有目的地设计试验,更快地找到核心启动子区[21-25]。本试验在DCT基因序列中发现了CpG岛、TATA box、GC box、CAAT box、M box和32 bp元件,这些都是启动子附近的标志性元件。而且,通过与GeneCopoeia上公布的人和小鼠DCT基因启动子序列进行比对,发现高度相似性序列也覆盖了这些元件。根据这些序列特征,再参考在线启动子预测结果,首先构建了一系列5′端序列缺失报告基因载体,最长片段为2 059 bp。结果显示,位于-990~+232 bp位置的片段启动子活性值最高,不同缺失片段之间启动子活性都存在极显著差异,根据活性变化情况可推测出-990~-204 bp区域内可能存在正调控元件,在-1 366~-990 bp区域内可能存在负调控元件。结合对基因序列特征的分析,发现片段P4的活性比P3活性显著降低,在P3片段序列中包含32 bp元件,而P4片段中则没有,此元件已被证实是DCT基因在色素细胞表达中所必需的[5],因此可推测,32 bp元件对山羊DCT基因的转录起正调控作用。
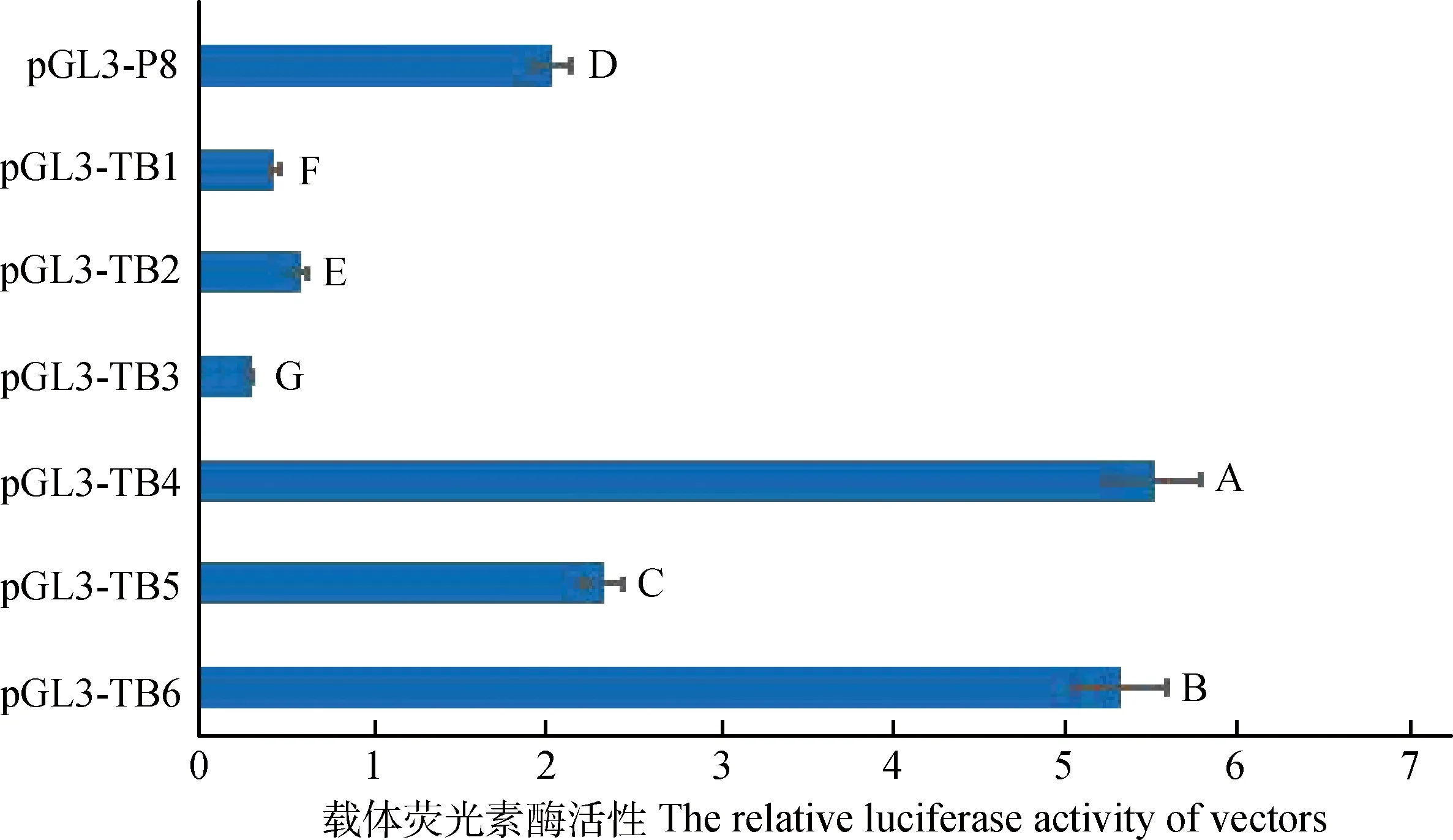
图5 点突变报告基因载体与模板P8载体荧光素酶相对活性Fig.5 Luciferase relative activity of mutational and original reporter gene vectors and fragment P8
根据5′系列启动子缺失片段活性检测结果,在最高活性-990~+232 bp片段的基础上,构建了一系列3′端序列缺失报告基因载体,最长片段为1 024 bp。启动子活性检测发现,-881~-154 bp片段的启动子活性最高,该序列包括32 bp元件、M box和2个TATA box,推测这些调控元件组合可维持山羊DCT基因的基本转录。不同缺失片段启动子活性之间都存在显著差异,根据活性变化情况可推测核心启动子区为-881~-232 bp区,该区域包含2个预测启动子位置,具有32 bp元件、2个TATA box和1个M box,这些元件组合发挥了基本转录活性。-192~-154 bp间的CAAT box为DCT基因的关键调控元件,发挥增强子作用。-154~+143 bp区域存在的GC box和CpG岛,对DCT基因的转录发挥负调控作用。
3.2 利用点突变DCT基因启动子活性片段检测转录因子调控作用
本试验在前人对不同物种DCT基因的研究基础上,筛选出了3种与DCT基因转录表达相关的转录因子:SOX10、MITF和OTX2。通过点突变的方法引起转录因子SOX10结合位点消失后的3个片段活性均显著降低,即SOX10能够激活DCT基因的表达,这与前人研究结果一致[12-13]。另外,在5′端系列缺失报告基因载体中,片段P3的启动子活性要显著高于P4,片段P10的启动子活性要显著高于P11,而片段P3和P10序列中都包含了转录因子SOX10的3处结合位点,片段P4和P11则没有SOX10的结合位点,这一结果进一步说明了转录因子SOX10有促进山羊DCT基因转录的作用。
而目前关于转录因子MITF对DCT基因的调控作用尚存在一些争议,有研究认为,MITF能够协同转录因子SOX10刺激并激活DCT基因启动子的转录[14-15],也有研究认为,DCT基因的转录调控不依赖于MITF,或者MITF对DCT基因发挥的是负调控作用[16-18]。本试验共突变了2处转录因子MITF的结合位点,结果显示,突变后片段的启动子活性都显著增强,但在首轮启动子缺失片段中,片段P4序列包含了转录因子MITF的2个结合位点,而片段P5不包含,但片段P4的活性却显著高于片段P5,与MITF结合位点点突变的结果相矛盾,因此产生了2种推测结果:第一,转录因子MITF抑制DCT基因的表达,但由于片段P4序列中含有其它正调控元件,而这些正调控原件发挥了主要作用;第二,DCT基因可能存在其它不依赖MITF的转录调控机制。转录因子OTX2对DCT基因调控作用的研究目前还不是很多,有研究表明,在人视网膜色素上皮细胞中转录因子OTX2能够激活DCT基因的表达,但目前还没有在黑色素细胞中的研究证明[20]。本试验通过定点突变改变转录因子OTX2的结合位点,结果显示,突变后片段启动子活性显著上升,说明转录因子OTX2可能对山羊DCT基因的转录发挥负调控作用。
本研究使用多种实验动物模型综合评价注射用雷贝拉唑钠的药理学作用。胃溃疡模型有多种,目前比较常用的大鼠胃溃疡模型主要有应激性溃疡(水浸应激模型)、化学因素型胃溃疡模型(吲哚美辛法、醋酸性)及幽门结扎型大鼠胃溃疡模型等,随着分子生物学技术的发展,相关技术也应用到溃疡模型制备的领域中,如基因敲除技术、蛋白缺失技术等[11]。
4 结 论
本试验成功构建了5个5′端和6个3′端序列缺失启动子报告基因载体,确定山羊DCT基因启动子核心区位于-881~-232 bp区域;-192~-154 bp区域内可能存在正调控元件,发挥增强子作用;-154~+143 bp区域存在的GC box和CpG岛对DCT基因的转录发挥负调控作用。转录因子SOX10对山羊DCT基因转录起正调控作用,而转录因子MITF和OTX2对山羊DCT基因的转录调控作用尚需继续深入研究。
[1] 舒 文, 毛华明. 黑色素的研究进展[J]. 国外畜牧学: 猪与禽, 2003, 23(2): 31-34.
SHU W, MAO H M. Advances in melanin[J].AnimalScienceAbroad-PigsandPoultry, 2003, 23(2): 31-34. (in Chinese)
[2] 刘甲斐, 仇雪梅. 黑色素及其相关基因的研究进展[J]. 生物技术通报, 2007(4): 55-58.
LIU J F, QIU X M. Melanin and its relevant gene research progress[J].BiotechnologyBulletin, 2007(4): 55-58. (in Chinese)
[3] 刘伟兰, 李祥龙, 周荣艳, 等. 不同物种TYRP2基因完整编码区生物信息学分析[J]. 河南农业科学, 2011, 40(10): 144-148.
LIU W L, LI X L, ZHOU R Y, et al. Bioinformatics analysis of complete coding regions of TYRP2 gene among 21 species[J].JournalofHenanAgriculturalSciences, 2011, 40(10): 144-148. (in Chinese)
[4] 白春雨, 高玉花, 庞全海. 影响动物毛色的基因[J]. 国外畜牧学: 猪与禽, 2008, 28(5): 71-73.
BAI C Y, GAO Y H, PANG Q H. Gene that affect the coat color of animals[J].AnimalScienceAbroad-PigsandPoultry, 2008, 28(5): 71-73. (in Chinese)
[5] YOKOYAMA K, YASUMOTO K, SUZUKI H, et al. Cloning of the human DOPAchrome tautomerase/tyrosinase-related protein 2 gene and identification of two regulatory regions required for its pigment cell-specific expression[J].JBiolChem, 1994, 269(43): 27080-27087.
[6] 郭 敏, 张大山, 李祥龙. TYR基因及其与动物毛色关系的研究进展[J]. 黑龙江畜牧兽医, 2017(1): 83-85.
GUO M, ZHANG D S, LI X L. Advances in TYR gene and the relationship with the coat color of animals[J].HeilongjiangAnimalScienceandVeterinaryMedicine, 2017(1): 83-85. (in Chinese)
[7] JACKSON I J, CHAMBERS D M, TSUKAMOTO K, et al. A second tyrosinase-related protein, TRP-2, maps to and is mutated at the mouse slaty locus[J].EMBOJ, 1992, 11(2): 527-535.
[8] 于秀菊, 范阔海. 毛囊的黑色素沉着[J]. 畜牧兽医科技信息, 2009(4): 16-17.
YU X J, FAN K H. melanin pigmentation in hair follicle[J].ChineseJournalofAnimalHusbandryandVeterinaryMedicine, 2009(4): 16-17. (in Chinese)
[9] GUYONNEAU L, MURISIER F, ROSSIER A, et al. Melanocytes and pigmentation are affected in dopachrome tautomerase knockout mice[J].MolCellBiol, 2004, 24(8): 3396-3403.
[10] 刘相莲, 乔 芳, 黄德玉, 等. 转录因子研究方法进展[J]. 生理科学进展, 2017, 48(1): 73-76.
LIU X L, QIAO F, HUANG D Y, et al. Advances in the Research method of transcription factor[J].ProgressinPhysiologicalSciences, 2017, 48(1): 73-76. (in Chinese)
[11] 侯 琳, 钱敏平, 朱云平, 等. 转录因子结合位点生物信息学研究进展[J]. 遗传, 2009, 31(4): 365-373.
HOU L, QIAN M P, ZHU Y P, et al. Advances on bioinformatic research in transcription factor binding sites[J].Hereditas, 2009, 31(4): 365-373. (in Chinese)
[12] JALILIAN N, TABATABAIEFAR M A, ALIMADADI H, et al.SOX10 mutation causes Waardenburg syndrome associated with distinctive phenotypic features in an Iranian family: A clue for phenotype-directed genetic analysis[J].IntJPediatrOtorhinolaryngol, 2017, 96(5): 122-126.
[13] POTTERF S B, MOLLAAGHABABA R, HOU L, et al. Analysis of SOX10 function in neural crest-derived melanocyte development: SOX10-dependent transcriptional control of dopachrome tautomerase[J].DevBiol, 2001, 237(2): 245-257.
[14] LUDWIG A, REHBERG S, WEGNER M. Melanocyte-specific expression of dopachrome tautomerase is dependent on synergistic gene activation by the Sox10 and Mitf transcription factors[J].FEBSLett, 2004, 556(1-3): 236-244.
[15] JIAO Z X, MOLLAAGHABABA R, PAVAN W J, et al. Direct interaction of Sox10 with the promoter of murineDopachrometautomerase(Dct) and synergistic activation ofDctexpression with Mitf[J].PigmCellRes, 2004, 17(4): 352-362.
[16] BERTOLOTTO C, BUSCR, ABBE P, et al. Differentcis-acting elements are involved in the regulation of TRP1 and TRP2 promoter activities by cyclic AMP: pivotal role of M boxes (GTCATGTGCT) and of microphthalmia[J].MolCellBiol, 1998, 18(2): 694-702.
[17] 黎 钊, 王 平, 洪为松. 正常人黑素细胞MITF对酪氨酸酶相关蛋白的转录调控研究[J]. 医学研究杂志, 2013, 42(3): 58-62.
LI Z, WANG P, HONG W S. Effect of MITF transcriptional control on family of tyrosinase-related proteins in normal human melanocytes[J].JournalofMedicalResearch, 2013, 42(3): 58-62. (in Chinese)
[18] 刘 栋, 朱文元. MITF与黑素细胞的发育、分化和功能调节[J]. 细胞生物学杂志, 2002, 24(6): 346-351.
LIU D, ZHU W Y. The development, differentiation and regulation function of melanocyte with MITF gene[J].ChineseJournalofCellBiology, 2002, 24(6): 346-351. (in Chinese)
[19] YASUMOTO K, YOKOYAMA K, TAKAHASHI K, et al. Functional analysis of microphthalmia-associated transcription factor in pigment cell-specific transcription of the human tyrosinase family genes[J].JBiolChem, 1997, 272(1): 503-509.
[20] TAKEDA K, YOKOYAMA S, YASUMOTO K, et al. OTX2 regulates expression of DOPAchrome tautomerase in human retinal pigment epithelium[J].BiochemBiophResCommun, 2003, 300(4): 908-914.
[21] 汤 方, 涂慧珍. 真核启动子研究进展[J]. 林业科技开发, 2015, 29(2): 7-12.
TANG F, TU H Z. Advances in eukaryotic promoter[J].ChinaForestryScienceandTechnology, 2015, 29(2): 7-12. (in Chinese)
[22] 王秋岩, 何淑雅, 马 云, 等. 启动子分析方法的研究进展[J]. 现代生物医学进展, 2015, 15(14): 2794-2800.
WANG Q Y, HE S Y, MA Y, et al. Advances in analytical methods of promoter[J].ProgressinModernBiomedicine, 2015, 15(14): 2794-2800. (in Chinese)
[23] 姚一龙, 李伯江, 李齐发. 牦牛FKBP6基因启动子区克隆、鉴定与分析[J]. 畜牧兽医学报, 2017, 48(1): 52-59.
YAO Y L, LI B J, LI Q F. Cloning and identification of promoter of Yak FKBP6 gene[J].ActaVeterinariaetZootechnicaSinica, 2017, 48(1): 52-59. (in Chinese)
[24] 李丽莎, 彭永东, 郑晓宁, 等. 山羊PMEL基因启动子活性及转录调控元件分析[J]. 畜牧兽医学报, 2017, 48(5): 826-835.
LI L S, PENG Y D, ZHENG X N, et al. Analysis of the promoter activity and transcriptional regulatory elements of goat PMEL gene[J].ActaVeterinariaetZootechnicaSinica, 2017, 48(5): 826-835. (in Chinese)
[25] 胡慧艳, 贾 青, 侯胜奎, 等. 猪DKK1基因启动子区的克隆及其活性分析[J]. 畜牧兽医学报, 2017, 48(6): 1150-1157.
HU H Y, JIA Q, HOU S K, et al. Cloning and activity analysis of the promoter region of swineDKK1 gene[J].ActaVeterinariaetZootechnicaSinica, 2017, 48(6): 1150-1157.
