Nogo-B通过Rac1 GTPase激活途径调控巨噬细胞的胞葬功能
2018-01-23姜春玲蒋小娟肖乾慧毛永巧
张 英,周 莉,姜春玲*,蒋小娟,肖乾慧,毛永巧
(1.北京大学深圳医院,广东 深圳518036;2 .四川大学华西医院,四川 成都610041)
细胞凋亡是机体清除感染或受损细胞,维持内稳态的重要机制。但凋亡细胞内含有大量自身抗原与警报素,若未被及时清除,会导致继发性坏死,引起组织损伤甚至自身免疫性疾病[1]。巨噬细胞密切参与凋亡细胞的清除,其将凋亡细胞吞噬、移除的过程,称为胞葬功能(Efferocytosis)[2]。胞葬功能对维持机体生长发育、内环境稳定等至关重要[3]。
Nogo-B是一类主要定位于真核细胞内质网及细胞膜的跨膜蛋白,其高表达于单核/巨噬细胞中,对巨噬细胞功能具有一定调节作用[4]。研究表明Nogo-B 基因缺陷可导致单核/巨噬细胞的迁移、延展能力降低以及肌动蛋白细胞骨架重组功能障碍[4]。
肌动蛋白细胞骨架重组是巨噬细胞完成吞噬的必经步骤[5]。Rho/Rac GTPase是介导胞葬功能时肌动蛋白细胞骨架重组与凋亡细胞吞入的重要蛋白酶。激活Rac1可促进肌动蛋白细胞骨架重组,完成凋亡细胞吞入[6]。Schanda等研究发现,在单核/巨噬细胞系统中,Nogo-B与RhoA/Rac1存在共定位关系[7],因此二者可能存在功能上相互关联。
基于以上认识,本实验中我们采用siRNA转染下调巨噬细胞Nogo-B表达,通过观察其对凋亡细胞吞噬指数及在凋亡细胞刺激下Rac1 GTPase活性的变化,探讨Nogo-B是否对巨噬细胞的胞葬功能具有调控作用及其可能机制。
1 材料与方法
1.1细胞株及试剂
J774A.1小鼠巨噬细胞与HL60早幼粒细胞株(美国伊利诺伊大学芝加哥分校麻醉与药理实验室胡国昌老师惠赠);胎牛血清(Invitrogen,美国)、 DMEM 培养基(Thermo Fisher Scientific,美国)、RPMI-1640 培养基(Gibco,美国);兔抗小鼠 Nogo-B抗体(Abcam,美国),兔抗小鼠GAPDH抗体 (Cell Signaling Technology,美国);RhoA/Racl/Cdc42 活性分析试剂盒(Cytoskeleton,美国);膜联蛋白Ⅴ(AnnexinⅤ)-FITC(含 PI)双染凋亡试剂盒(BD Biosciences,美国);Nogo-B siRNA(Dharmacon,美国);红色与绿色荧光CellTracker(Invitrogen,美国);Lipofectamine RNAiMAX转染试剂(Invitrogen,美国)
1.2细胞培养与分组
J774A.1巨噬细胞用含10%胎牛血清的DMEM培养基置于37℃、5%CO2孵育箱中培养。细胞自然生长状态下汇合50%-70%时分别转入不同浓度(10、20、50、100 nM)Nogo-B siRNA,转染48 h以western印迹法检测转染效果,优化最佳转染浓度。HL60细胞用10%胎牛血清的RPMI-1640培养基置于37℃、5%CO2孵育箱中培养。
分组:将J774A.1巨噬细胞分为siRNA 阴性对照组(对照组)与Nogo-B siRNA 干扰组。对照组与Nogo-B siRNA组分别取生长状态良好的J774A.1巨噬细胞,用细胞刮刮取细胞,反复吹打至单细胞悬液、计数,按每孔5×105个细胞铺于普通6孔板中,待细胞完全贴壁后分别使用阴性对照siRNA与Nogo-B siRNA进行转染,48 h后检测巨噬细胞对凋亡细胞的胞葬功能以及在凋亡细胞刺激下巨噬细胞内Rac1 GTPase活性变化。所有实验于相同条件下重复3次。
1.3紫外线法诱导HL60细胞凋亡
取107个HL60细胞,离心去上清,以无色、无血清RPMI培养基10 ml重悬。置于短波紫外灯下室温照射后(波长254 nm,能量100 J/m2),转移至 37℃,5% CO2培养箱中孵育3 h。使用Annexin V-FITC/PI双染试剂盒染色后,以FACSCanto流式细胞仪(Beckman Coulter,美国)鉴定凋亡细胞比例,其中Annexin V 阳性,PI 阴性为凋亡细胞,本方法可获得90%以上细胞发生凋亡(图1)。
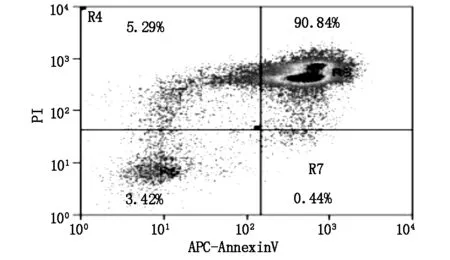
图1 紫外线法诱导细胞凋亡
1.4体外检测巨噬细胞的胞葬功能
实验前1日,将105/well巨噬细胞接种于24孔板。吞噬前分别采用5 μM绿色与2 μM红色荧光celltracker对巨噬细胞与凋亡细胞分别染色30 min及15 min,以PBS洗涤一次以去除多余染料,按照5∶1的比例将凋亡细胞加入已接种巨噬细胞的24孔板中,置于37℃,5%CO2培养箱共同孵育2 h,使巨噬细胞充分吞噬凋亡细胞后,免疫荧光显微镜下观察,计算巨噬细胞对凋亡细胞的吞噬指数(每100个巨噬细胞所吞噬的凋亡细胞总数除以100,取三次计数的平均值)。
1.5Western印迹检测细胞蛋白表达
接种于6孔板中的J774A.1巨噬细胞,加入RIPA裂解液,离心取上清液,BCA法测定各组蛋白含量。加入5X上样缓冲液,100℃水浴变性提取蛋白,并在SDS-聚丙烯酰胺凝胶中进行电泳,经转膜仪将蛋白转移至硝酸纤维素膜后,经5 %脱脂奶粉室温封闭2 h。分别加入Nogo-B抗体(1∶1 000)、GAPDH抗体(1∶2 000)、4℃过夜,次日使用PBST洗膜后,加入辣根过氧化物酶标记的二抗(1∶2 000)室温孵育1 h,再次洗膜。蛋白条带以Odyssey Fcimager (LI-COR Biosciences,美国)化学发光/荧光检测数字成像系统进行检测并拍照,应用ImageJ 软件分析条带光密度。
1.6谷胱甘肽巯基转移酶-拉下实验(GST-pulldownassay)检测Rac1GTPase活性
按照RhoA/Racl/Cdc42 活性分析试剂盒操作说明进行。该试剂盒中含有被标记Rho GTPase下游效应蛋白的琼脂糖珠,该琼脂糖珠可特异性结合处于活性状态的Rho GTPase,将其从琼脂糖珠洗脱下后,通过Western印迹法可检测活性Rac1 GTPase的含量。以12%SDS-PAGE行Western印迹分析(同前述)。
1.7统计学处理
2 结果
2.1Nogo-B对巨噬细胞胞葬功能的调控作用
首先采用Nogo-B siRNA(50 nM)转染、下调J774A.1巨噬细胞Nogo-B 表达,以绿色与红色荧光celltracker对巨噬细胞与凋亡细胞进行染色,随后共同孵育、吞噬2 h,免疫荧光显微镜下观察结果。我们发现,Nogo-B下调导致巨噬细胞对凋亡细胞的胞葬功能明显受损,吞噬指数由对照组的(48.9±3.5)%显著降低至(17.8±2.1)%(P<0.01)(图2),提示Nogo-B对巨噬细胞的胞葬功能具有调控作用。
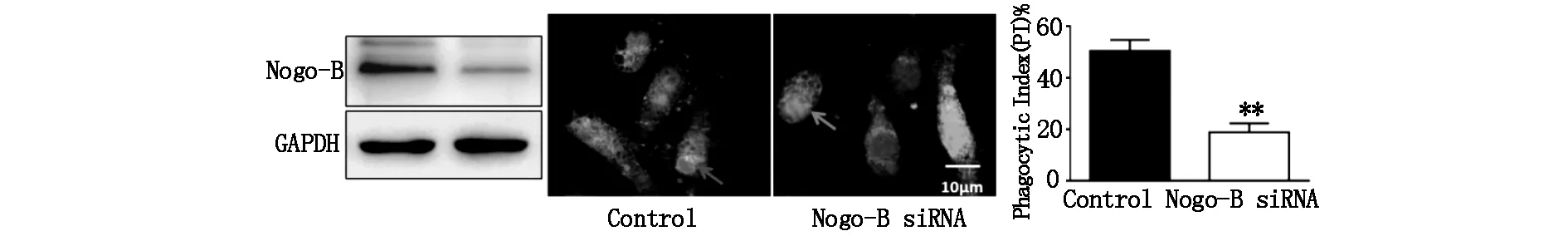
图2 下调Nogo-B表达抑制巨噬细胞的胞葬功能 **P<0.01
2.2Nogo-B对巨噬细胞RaclGTPase活性的影响
为进一步明确Nogo-B如何调控巨噬细胞的胞葬功能,我们使用siRNA转染下调巨噬细胞Nogo-B表达后,给予凋亡细胞刺激,分别于刺激5、10及15 min,采集巨噬细胞提取蛋白,采用pulldown技术检测Rac1 GTPase活性变化。结果发现,对照组巨噬细胞在凋亡细胞刺激下Rac1 GTPase被逐渐激活,于刺激后的5至15 min其活性呈递增趋势;而使用siRNA下调Nogo-B表达后的巨噬细胞,在凋亡细胞刺激下Rac1活性无显著变化,即出现Rac1 GTPase激活障碍(图3)。肌动蛋白细胞骨架重组是巨噬细胞完成吞噬的必经步骤,而Rac1 GTPase是完成细胞骨架重组的重要分子,Rac1激活障碍将影响巨噬细胞肌动蛋白细胞骨架重组并可能导致吞噬功能受损。因此,该结果提示Nogo-B 可能通过调控Rac1 GTPase激活途径影响细胞骨架重组,进而调节巨噬细胞的胞葬功能。

*P<0.05,**P<0.01
3 讨论
人体每天有数以亿计的细胞发生凋亡。若凋亡细胞未被及时清除,就会继发性坏死,导致细胞内损伤性物质大量释放、损伤机体。而巨噬细胞可以通过胞葬功能,在损伤性物质释放到周围组织之前将凋亡细胞吞噬并安全移除,因此胞葬功能对维持机体生长发育、内环境稳定等至关重要[3]。我们的研究发现, Nogo-B可以调控巨噬细胞的胞葬功能,采用siRNA转染下调Nogo-B 表达后,巨噬细胞对凋亡细胞的胞葬功能明显受损;Nogo-B 可能通过Rac1 GTPase激活途径,影响细胞骨架重组,发挥对胞葬功能的调节作用。
胞葬功能受机体严密调控,胞葬凋亡细胞的过程包括凋亡细胞的识别、吞入及消化等多个环节[3,8],对任一环节的调控均可能影响胞葬功能。Nogo-B是一类主要定位于真核细胞内质网及细胞膜的跨膜蛋白,其高表达于单核/巨噬细胞中。已有研究发现,Nogo-B对巨噬细胞功能具有一定调节作用[4],Nogo-B 基因缺陷可导致单核/巨噬细胞的迁移、延展能力降低以及肌动蛋白细胞骨架重组功能障碍[4]。而我们的研究进一步发现,Nogo-B对巨噬细胞的胞葬功能也有调控作用,下调Nogo-B后巨噬细胞的胞葬功能严重受损,对凋亡细胞的吞噬指数由对照组的48.9±3.5%显著降低至17.8±2.1%。
那么网状蛋白Nogo-B是如何调节巨噬细胞的胞葬功能呢?已知胞葬过程中当凋亡细胞直接或间接被吞噬细胞识别后,最终导致Rho/Rac GTPase激活[9,10],促进纤丝状肌动蛋白F-actin聚合并由此介导细胞骨架重组完成吞入及消化。Rac1 作为Rac GTPase的重要成员,是影响细胞骨架重组的关键分子,Rac1激活后可通过诱导肌动蛋白聚合、F-actin形成介导肌动蛋白细胞骨架重组[11-13]。肌动蛋白细胞骨架重组是巨噬细胞完成吞噬的必经步骤[5]。而现有研究提示,Nogo-B基因缺陷可导致小鼠骨髓单核/巨噬细胞细胞F-actin聚合及由此介导的肌动蛋白细胞骨架重组功能障碍[4]。此外,有研究进一步发现在单核/巨噬细胞系统中,Nogo-B与RhoA/Rac1存在共定位关系[7],提示二者可能存在功能上相互关联。本研究中我们发现,使用siRNA转染下调巨噬细胞Nogo-B表达后,Rac1 GTPase的激活过程受到抑制。提示,Nogo-B可能通过调控Rac1 GTPase的激活途径以及由此介导的肌动蛋白细胞骨架重组,调控巨噬细胞的胞葬功能。
综上所述,网状蛋白Nogo-B对巨噬细胞的胞葬功能具有调控作用,下调Nogo-B 表达后巨噬细胞的胞葬功能明显受损;Nogo-B 可能通过Rac1 GTPase激活途径,影响细胞骨架重组,发挥对胞葬功能的调节作用。
[1]Elliott MR,Ravichandran KS.Clearance of apoptotic cells:implications in health and disease[J].J Cell Biol,2010,189(7):1059.
[2]deCathelineau AM,Henson PM.The final step in programmed cell death:phagocytes carry apoptotic cells to the grave.Essays Biochem,2003,39:105.
[3]Devitt A,Marshall LJ.The innate immune system and the clearance of apoptotic cells[J].J Leukoc Biol,2011,90:447.
[4]Yu J,Fernandez-Hernando C,Suarez Y,et al.Reticulon 4B (Nogo-B) is necessary for macrophage infiltration and tissue repair[J].Proc Natl Acad Sci USA,2009,106:17511.
[5]May RC,Machesky LM.Phagocytosis and the actin cytoskeleton[J].J Cell Sci,2001,114:1061.
[6]Nakaya M,Tanaka M,Okabe Y,et al.Opposite effects of rho family GTPases on engulfment of apoptotic cells by macrophages[J].J Biol Chem ,2006,281:8836.
[7]Schanda K,Hermann M,Stefanova N,et al.Nogo-B is associated with cytoskeletal structures in human monocyte-derived macrophages[J].BMC Res Notes,2011,4:6.
[8]Hochreiter-Hufford A,Ravichandran KS.Clearing the dead:apoptotic cell sensing,recognition,engulfment,and digestion[J].Cold Spring Harb Perspect Biol,2013,5:a008748.
[9]Leverrier Y,Ridley AJ.Requirement for Rho GTPases and PI 3-kinases during apoptotic cell phagocytosis by macrophages[J].Curr Biol,2001,11:195.
[10]Tosello-Trampont AC,Nakada-Tsukui K,Ravichandran KS.Engulfment of apoptotic cells is negatively regulated by Rho-mediated signaling[J].J Biol Chem,2003,278:49911.
[11]Wells CM,Walmsley M,Ooi S,et al.Rac1-deficient macrophages exhibit defects in cell spreading and membrane ruffling but not migration[J].J Cell Sci,2004,117:1259.
[12]Allen WE,Jones GE,Pollard JW,et al.Rho,Rac and Cdc42 regulate actin organization and cell adhesion in macrophages[J].J Cell Sci,1997,110 ( Pt 6):707.
[13]Wheeler AP,Wells CM,Smith SD,et al.Rac1 and Rac2 regulate macrophage morphology but are not essential for migration[J].J Cell Sci,2006,119:2749.
