扁穗雀麦优良品系数量性状的变异分析及遗传参数评估
2018-01-19孙铭雷雄张新全张成林伍文丹赵文达杨晓鹏马啸
孙铭,雷雄,张新全,张成林,伍文丹,赵文达,杨晓鹏,马啸
(四川农业大学动物科技学院,四川 成都611130)
扁穗雀麦(Bromuscatharticus)为一年生或越年生的冷季型牧草,属于禾本科(Gramineae)雀麦属(Bromus),为异源六倍体且具有兼性闭花授粉的特性,原生于南美洲阿根廷潘帕斯草原,现已在全世界温带地区广为栽培[1-3]。由于其具有产量高、秋冬季生长速度快、适应性强和种子成熟后仍能保持青绿的特点[4-5],越来越受国内外饲草育种研究者的关注。我国扁穗雀麦种质资源主要分布于西南、华中、华北和西北地区,而且在部分地区有逸生资源存在[4],这为基于逸生或野生种质资源选育优良品种提供了便利。此外,冬春季家畜饲草不足一直是我国大多数地区普遍存在的问题,严重限制了我国农区草牧业的发展,因此对本土扁穗雀麦种质资源进行综合评价培育优良品种显得尤为重要。
形态学评价是研究植物遗传多样性和筛选育种材料的重要手段[6-8],与细胞学和分子生物学方法相比操作简单且效率高。已有多位学者对扁穗雀麦的部分表型性状进行了分析,并发现扁穗雀麦群体间和群体内均存在较高的表型变异[2],但对其表型的研究大多数都集中为与产草量相关性状的简单分析[3-4],而涉及遗传力和遗传进度等遗传参数的研究则较少。
广义遗传力和遗传进度是作物数量性状遗传研究中最重要的内容之一。广义遗传力是估测各种不同性状传递给下一代的能力强弱的重要指标[9]。遗传进度则反映了经过选择之后,子代从亲代获得的遗传增量[10]。测定作物主要性状,通过分析各性状的遗传变量,确定杂交组合及有效的育种方法,从而可以提高育种效果,加快新品种、新材料的创新进程。因此,这对于育种进展相对比较缓慢和研究相对滞后的扁穗雀麦具有重要意义。目前国内外的扁穗雀麦品种大多数为野生种质材料经过混合选择获得的品种,品种稳定性相对较差[4]。如果可以对部分具有优良性状的品种或品系进行遗传参数评估,找到一些遗传力较高的杂交亲本,利用杂交手段对具有兼性闭花授粉特性的扁穗雀麦进行品种培育,将对扁穗雀麦品种培育具有重要意义。
本研究旨在通过对目前国内仅有的扁穗雀麦品种“江夏”、“黔南”以及由四川农业大学草学系通过多年田间观测和筛选获得的9个扁穗雀麦新品系进行农艺性状测定,对这些种质材料的21个农艺性状进行变异分析、性状间相关性分析,并重点通过方差分析模型估算其方差成分和广义遗传力、遗传进度等部分遗传参数。
1 材料与方法
1.1 试验地概况
试验地位于四川农业大学教学科研基地(30°08′ N, 103°14′ E),海拔600 m,属北亚热带湿润季风气候区。年均气温16.2 ℃,极端高温37.7 ℃,极端低温-3 ℃,年降水量1774.3 mm,年蒸发量1011.2 mm,相对湿度79%,年日照时数1039.6 h,年无霜期304 d,≥10 ℃年积温5231 ℃。紫色土,土壤有机质含量14.6178 g·kg-1,全氮含量1.9070 g·kg-1,全磷含量0.0554 g·kg-1,全钾含量11.5793 g·kg-1,速效氮含量100.6289 mg·kg-1,速效磷含量4.7286 mg·kg-1,速效钾含量338.2429 mg·kg-1,pH为5.46。
1.2 植物材料
本试验选用的材料中两个为我国的国审品种“黔南”和“江夏”,分别由贵州省草业研究所和湖北省农业科学院畜牧研究所提供,其余材料为四川农业大学草学系从我国西南地区采集的逸生材料中筛选培育的具有植株高、叶片宽大、叶量丰富等饲草特性的新品系(表1)。2015年9月下旬,供试材料在四川农业大学雅安校区草学试验基地进行盆栽育苗,每份材料发芽60粒种子,待幼苗生长至三叶期后移栽至田间。田间试验采用完全随机区组设计(RCBD),每个品种或品系3个重复,每小区3行共12株,株距50 cm,行距50 cm。管理过程中保证了良好的排灌水,并进行了适当的杂草防除。2016年5-6月针对各材料进行数据收集和种子收获。

表 1 供试材料Table 1 Accessions of B. catharticus used in the study
1.3 性状数据测定
本试验中共测定21个农艺性状,其中营养生长性状10个,分别为:株高、茎粗、旗叶长、旗叶叶鞘长、旗叶宽、倒二叶长、倒二叶叶鞘长、倒二叶宽、单株分蘖数、第1节间长;生殖生长性状9个,分别为:花序长、花序宽、单株圆锥花序数目、花序节数、花序轴第1节间长、初级分枝数、单株小穗数、小穗平均长、小花数;产量性状2个,分别为单株干草产量、单株种子产量。所有性状在成熟期每小区随机选5个单株进行测定,方法如表2。
1.4 数据分析
通过利用SAS 9.2[11]和NTSYS 2.1[12]软件来评价供试材料的表型变异,并对方差分析达到显著的性状的方差成分、广义遗传力和遗传进度进行了估算。
描述性统计量:分析统计了供试材料的最大值、最小值、平均值、标准差和变异系数。
相关性分析:对供试材料的21个表型性状进行了Person 相关性分析, 并做了显著性检验。
聚类分析:通过对各性状的平均值进行标准化之后,利用NTSYS 2.1计算了各种质之间的欧氏距离,然后利用UPGMA 法进行了聚类分析。

表2 各性状测定方法Table 2 List of quantitative traits of new strains of B. catharticus
遗传参数估算: 对各性状进行方差分析,对F值测验达到显著的性状进行参数估算,采用了完全随机设计单因素方差分析模型估算了环境方差(σe2)、遗传方差(σg2)、 表型方差(σp2)、遗传变异系数(CVg)和表型变异系数(CVp),来揭示群体间存在的变异,计算模型如表3。


表3 方差分析的模型Table 3 Form of analysis of variance for each quantitative traits
MSa:材料均方 Mean square of accessions;MSe:机误均方Mean square of error;r:重复数Number of replications;n:材料数Number of accessions.

2 结果与分析
2.1 性状的描述性分析
供试11份材料的21个营养生长性状、生殖生长性状和产量性状的统计结果表明:各统计量在不同的性状间存在较大的差异,变异系数介于4.85%~34.37%之间,平均值为13.46%(表4)。在这些性状中单株种子产量、圆锥花序数、分蘖数、旗叶宽和单株干草产量表现出较大的变异值,分别为34.37%、31.42%、21.99%、19.44%和18.81%。变异系数较小的性状分别为平均小穗长(4.85%)、花序节数(5.08%)和小花数(5.16%)。总体而言,不同材料间性状存在较大的变异,其中16个性状的变异系数大于10%,这些具有优异产草量相关性状的材料可以被用来培育扁穗雀麦新品种。
2.2 相关性分析
性状间的相关性分析结果表明:各性状间存在复杂的相关性,其中部分性状间表现出极显著的正相关和负相关(表5)。表现出极显著相关性并具有高相关系数的依次为:倒二叶宽和茎粗(r=0.993,P<0.001),旗叶宽和茎粗(r=0.990,P<0.001),旗叶宽和倒二叶宽(r=0.989,P<0.001)以及圆锥花序数和分蘖数(r=0.900,P<0.001)。株高和旗叶宽、倒二叶宽、茎粗以及分蘖数表现出紧密的相关性,相关系数分别为0.946(P<0.001)、0.932(P<0.001)、0.925(P<0.001)和0.942(P<0.001)。单株干重和株高的相关性最高,相关系数为0.976(P<0.001),其次为旗叶宽(r=0.950,P<0.001)、分蘖数(r=0.948,P<0.001)、倒二叶宽(r=0.934,P<0.001)、茎粗(r=0.920,P<0.001)和花序数(r=0.900,P<0.001)。 此外,旗叶长和倒二叶长也达到了极显著相关性(r=0.951,P<0.001),但它们与单株干重的相关性均未达到显著水平。单株种子产量与花序数的相关性最高(r=0.988,P<0.001),分蘖数次之(r=0.960,P<0.001),接下来为单株干重(r=0.927,P<0.001)和株高(r=0.909,P<0.001)。而极显著的负相关存在于小花数与旗叶叶鞘长(r=-0.857,P<0.01)和第1节间长之间(r=0.758,P<0.001)。
2.3 聚类分析
对11份扁穗雀麦种质材料的21个表型性状的测定值标准化后进行了遗传距离计算和基于非加权组平均法(UPGMA)的聚类分析,结果表明:各材料之间的欧氏距离范围介于0.186(BCS1101和BCS1105之间)~2.148(BCS1106和江夏之间)中间,平均值为0.847(图1)。这些材料在阈值为0.847处被分为两组,其中所有的新品系组成了聚类结果中的第1组,两个国审品种组成了第2组。对聚类分析两组中与产草量和种子产量相关性达到极显著的性状进行方差齐性测试,发现只有小穗数为非齐性,然后对这些性状分别进行了齐性t检验和非齐性t检验(表6)。检验结果发现除小穗数以外,其余性状均达到极显著差异(P<0.005),第1组在产草量相关性状和种子产量相关性状上均优于第2组,即新品系在单株产量相关各性状上总体均优于两个国审品种。此外,聚类分析结果显示,第1组材料表现为植株高,叶片宽,茎秆粗壮,基部第1节间较短,分蘖数多,单株干重和种子产量高等特点,相反,第2组表现为植株较矮,叶片较窄,茎秆较细,基部第1节间较长,分蘖数较少,单株干重和种子产量相对较低等特点。在第1组内,BCS1103的株高和单株产草量最高,分蘖数较多;BCS1106的叶片最宽,分蘖数、圆锥花序数和小穗数最多,单株产草量和单株种子产量最高;BCS1109的基部第1节间,叶鞘最长,叶片最窄,茎秆最细,初级分枝数最多。在第2组内,黔南比江夏表现出更优异的产草相关性状,比如植株更高,叶片更宽,茎秆更粗壮等。

表4 各性状的描述性统计量Table 4 Descriptive statistical analysis of reproductive growth traits, vegetative growth traits and yield traits
2.4 方差分析和遗传参数估算
11份材料的21个性状的单因素方差分析结果显示(表7):15个表型性状在不同的材料间存在极显著差异,包括:株高(PH)、旗叶叶鞘长(PLSL)、倒二叶叶鞘长(PLSL)、旗叶宽(FLW)、倒二叶宽(PLW)、茎粗(SD)、分蘖数(TN)、花序长(PL)、圆锥花序数(NPPP)、小穗数(TNS)、花序节数(NNPP)、初级分枝数(TNPB)、小花数(NFS)、单株干重(DMY)、单株种子产量(SY);2个性状在不同材料间存在显著差异,为第1节间长(LFI)和倒二叶长(PLL)。其余性状在各材料之间的差异未达到显著水平。由此可见,观测的21个性状中大多数的性状在供试材料中存在较大的变异,这将为进行扁穗雀麦育种工作提供便利。


图1 基于表型性状的聚类分析Fig.1 UPGMA dendrogram of the 11 B. catharticus accessions based on 21 phenotypic characters

性状TraitsLevene的变异数相等测试Levene’stestforequalityofvarianceF值Fvalue显著性Significancet检验t-testforequalityofmeant值tvalue自由度df显著性(双尾)Significance(2-tailed)平均差异Meandifference标准误差StandarderrordifferencePH0.220.6476.719.000.00032.464.84FLW0.270.6148.239.000.0005.750.70PLW0.570.4688.559.000.0005.910.69SD0.100.7659.579.000.0002.420.25TN0.400.5414.799.000.00113.182.75NPPP0.210.6593.889.000.00412.763.29TNS13.220.0051.921.050.29719.8010.33DMY0.200.6695.019.000.00173.8514.74SY0.280.6114.169.000.00255.3913.32

表 7 各性状的方差分析Table 7 One-way analysis of variance for 21 traits among 11 B. catharticus accessions
MSa:材料均方 Mean square of accessions;MSe:机误均方Mean square of error;**:显著水平为P<0.01 Significant atP<0.01 level; *:显著水平为P<0.05 Significant atP<0.05 level.
对方差分析中达到极显著和显著水平的性状进行方差成分、表型变异系数、遗传变异系数、广义遗传力、遗传进度、相对遗传进度估算,结果显示(表8):这17个表型性状的遗传方差和遗传变异系数均低于表型方差和表型变异系数,说明扁穗雀麦这些性状的表现不仅仅由遗传因素,还受到生长环境的影响。各性状遗传方差和表型方差的变异幅度比较大,遗传方差从最低的0.19(第1节间长和花序节数)到1197.65(单株干重),而表型方差在各性状略高,最低为0.21(花序节数),最高为1212.26(单株干重)。遗传变异系数的变化范围为0.0383~0.3351,最高变异系数出现在单株种子产量,最低在旗叶长;表型变异系数的变化范围为0.0499~0.3437,最高为单株种子产量,最低为花序节数。参照前人的研究,遗传变异系数和表型变异系数高于0.2视为变异较大,变异系数处于0.1~0.2之间为中等,低于0.1视为变异较小[16]。根据这个标准,表型变异系数和遗传变异系数较高的分别为分蘖数(0.2199和0.2087),圆锥花序数(0.3142和0.2999),单株种子产量(0.3437和0.3351);变异系数中等的为株高(0.1060和0.1027),倒二叶叶鞘长(0.1516和0.1503),旗叶宽(0.1944 和0.1927),倒二叶宽(0.1803和0.1787),茎粗(0.1584和0.1561),小穗数(0.1644和0.1587),初级分枝数(0.1879和0.1862)和单株干重(0.1881和0.1870);变异系数较低的有第1节间长(0.0836和0.0691),旗叶叶鞘长(0.0722和0.0674),旗叶长(0.0508和0.0383),小穗长(0.0648和0.0543),花序节数(0.0499和0.0468)和小花数(0.0552 和0.0523)。
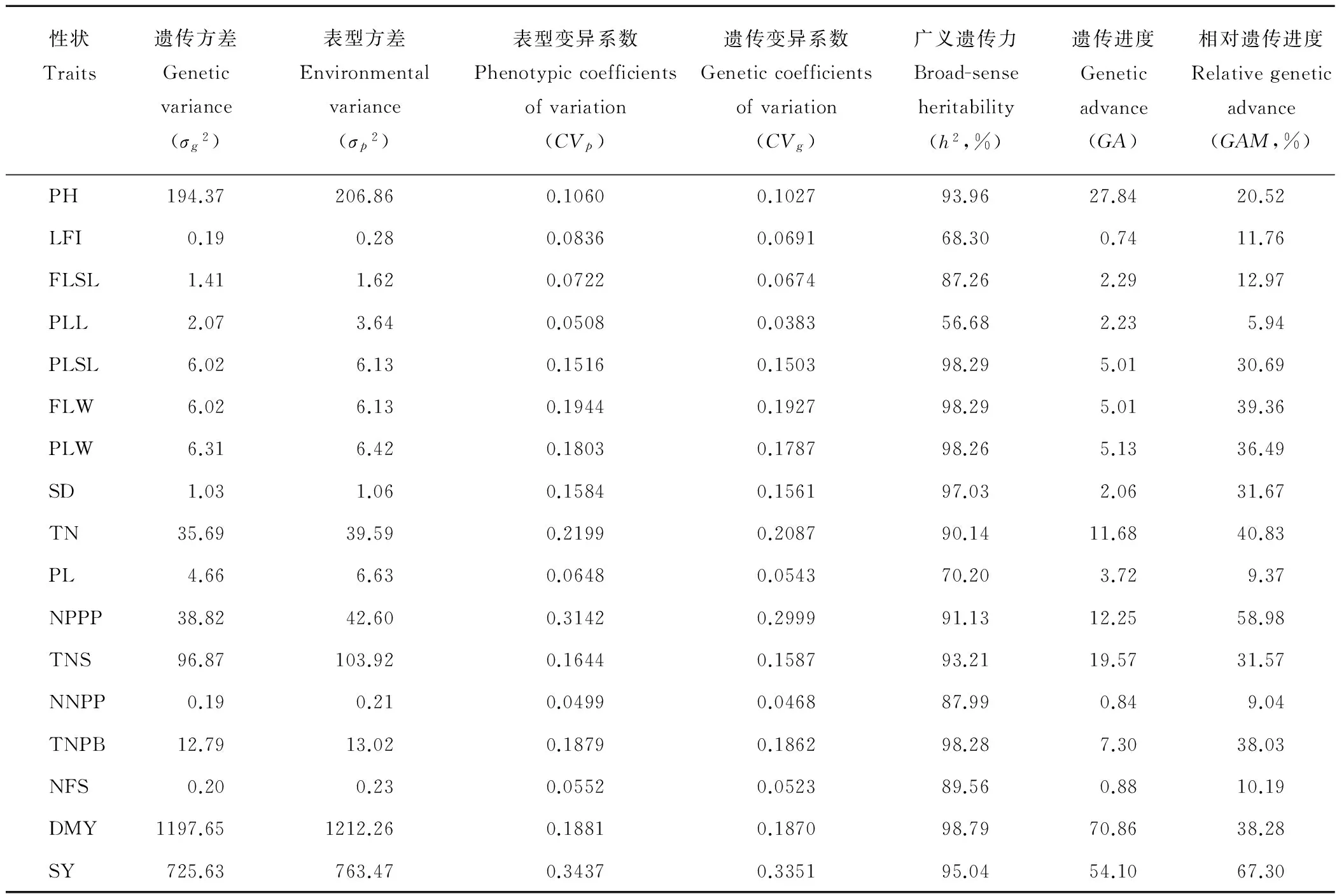
表8 各性状的方差成分、遗传变异系数、表型变异系数、广义遗传力、遗传进度和相对遗传进度Table 8 Variance component, coefficient of variation, heritability, genetic advance and relative genetic advance of 17 traits
广义遗传力和遗传进度是评价性状选择的重要指标,在本研究中各性状广义遗传力变化范围为0.5668~0.9879,平均值为0.8897。根据Singh[17]的研究,遗传力高于80%可以认为处于极高水平的遗传力,在60%~79%为中高水平,40%~59%为中等水平,低于40%为水平较低。因此本研究中所有性状均表现出了中等或者中等以上水平的遗传力,其中14个性状表现出极高的遗传力,2个性状表现出高水平的遗传力,只有1个性状表现出中等水平的遗传力。遗传进度的变化范围为 0.74~70.86,其中单株干重和单株种子重具有较高的遗传进度,第1节间长和小花数较低。相对遗传进度在本研究中也具有较大的变异,其中单株种子产量最高为67.30%,旗叶长最低为5.94%。按照 Johnson等[15]的标准:低于10%为遗传进度低,10%~20%为中等,高于20%为遗传进度高。本研究中具有较高遗传进度的有株高、倒二叶叶鞘长、旗叶宽、倒二叶宽、茎粗、分蘖数、花序数、小穗数、初级分枝数、单株干重和单株种子产量,中等的相对遗传进度表现在第1节间长、旗叶叶鞘长和小花数,低水平的遗传进度表现在旗叶长和花序节数。此外,广义遗传力和相对遗传进度之间存在显著的相关性,相关系数为0.613(P<0.01),而广义遗传力与遗传进度之间的相关性并未达到显著水平(P<0.05)。
3 结论与讨论
植物数量性状的变异,不仅受到基因型的控制,生长环境对其影响也十分重要。以往的扁穗雀麦表型性状的研究主要集中在其产草量相关的性状方面,比如:株高、茎粗、叶片长宽、分蘖数、单株生物量等方面,而很少研究其他形态或农艺性状[2-4]。本研究通过对11份扁穗雀麦品种(系)材料的营养生长、生殖生长以及生产性能相关的21个性状进行了全面考察,发现供试材料的部分农艺性状之间存在着极显著的相关性。其中与产草量达到极显著相关,且相关性系数高于0.9的性状依次为株高(r=0.976),旗叶宽(r=0.950)、分蘖数(r=0.948)、倒二叶宽(r=0.934)、茎粗(r=0.920)和花序数(r=0.900),与单株种子产量达到极显著相关,且相关系数高于0.9的依次为花序数(r=0.988),分蘖数(r=0.960),单株干重(r=0.927)和株高(r=0.909)。相关性规律除叶片长度以外,其余结果与马啸等[4]的研究结果基本一致。故而在扁穗雀麦牧草品种选育时应该重点考虑植株高度、叶片宽度、分蘖数、茎粗和花序数目,而叶片长度可以作为以上指标差异不显著时的参考指标。
广义遗传力是植物数量性状遗传传递的重要指标。广义遗传力大的数量性状,根据表现型选出的优良个体的基因型有较大的可能传递到下一代[18]。本研究中参与广义遗传力估算的17个性状中,82%的性状广义遗传力达到极高水平(广义遗传力大于80%),18%的性状广义遗传力处于中等水平或中高水平(广义遗传力介于40%~79%)。因此,广义遗传力高的性状如:株高、旗叶宽、倒二叶宽、旗叶叶鞘长、倒二叶叶鞘长、茎粗、分蘖数、花序数、小穗数、花序节数、初级分枝数、小花数、单株干重、单株种子产量等不容易受到环境的影响,可以在早代进行严格的选择。广义遗传力中等的性状如第1节间长、花序长和旗叶长则受环境影响相对稍大,可以在早代性状比较稳定后选择更有效。值得注意的是,一些研究表明广义遗传力并不能作为性状选择效果的唯一指标,因为高的遗传力可能被低的遗传进度所取代,而且在一定的选择率下广义遗传力会随着遗传变异系数的大小而发生变化[19-21]。一般情况下,遗传变异系数大,说明遗传变异的范围大,选择机会多,选择效果也会更好[10]。遗传进度则考虑了广义遗传力和遗传变异系数,被认为更可靠的选择指标。本研究中相对遗传进度与遗传变异系数和广义遗传力之间的相关性均达到了极显著水平,这为评估选择效果提供了更加便利的途径,可以通过其中一项指标对选择效果进行初步评估,然后结合其余指标进行综合评价,从而大大提高评估选择效果的效率。根据本研究的结果,综合考虑3种遗传参数,株高、倒二叶叶鞘长、旗叶宽、倒二叶宽、茎粗、分蘖数、花序数、小穗数、初级分枝数、单株干重和单株种子产量均有较高的遗传变异系数、极高的广义遗传力和相对遗传进度,其在育种选择中的效果会比较好。结合性状间相关性分析结果可知,在注重产草量对扁穗雀麦进行选择时,株高、旗叶宽、分蘖数、倒二叶宽、茎粗和花序数的可靠性较大,选择效果较好。在注重种子产量的扁穗雀麦育种选择时,分蘖数、花序数、单株生物量和株高的可靠性较大,效果较好,而小花数和花序节数作为种子产量的重要指标,遗传变异系数和遗传进度相对较低,所以在选择时应该慎重。
本研究中所使用的扁穗雀麦新品系均为从我国西南地区采集的野生或逸生种质经过筛选和连续多年的单株选择而获得的,在生产性能和抗逆性相关性状上具有较多的优势。在本研究的聚类分析和t检验结果中,可以看出通过多年选择、驯化的扁穗雀麦新品系和已有国审品种被明显的分为两大类,新品系与供试品种相比具有植株高、叶片宽、分蘖数多、茎秆粗等特点,在产草性状和种子生产性状方面均占有明显优势,具有成为牧草型扁穗雀麦新品种的潜力。此外,通过本研究对扁穗雀麦新品系和审定品种进行遗传参数评估,可以通过分析各性状的遗传变量,确定杂交组合及有效的育种方法,从而培育出稳定好、产量高的扁穗雀麦杂交品种。
References:
[1] Zhou X, Wang Q, Chen G. Research progress ofBromuscatharticus. Prataculture & Animal Husbandry, 2014, (4): 54-56.
周潇, 王巧, 陈刚. 扁穗雀麦研究进展. 草业与畜牧, 2014, (4): 54-56.
[2] Aulicino M B, Arturi M J. Phenotypic diversity in Argentinian populations ofBromuscatharticus(Poaceae). Genetic and environmental components of quantitative traits. New Zealand Journal of Botany, 2002, 40(2): 223-234.
[3] Hao F, Xu Z, Yan W H,etal. Morphological variations analysis ofBromuscatharticusgermplasm. Acta Agrestia Sinica, 2011, 19(4): 668-673.
郝峰, 徐柱, 闫伟红, 等. 扁穗雀麦种质资源形态变异分析. 草地学报, 2011, 19(4): 668-673.
[4] Ma X, Zhou C J, Zhang C L,etal. Patterns of morphological variation and agronomic traits in a worldwide sample of prairie grass germplasm. Acta Agrestia Sinica, 2015, 23(5): 1048-1056.
马啸, 周朝杰, 张成林, 等. 扁穗雀麦种质资源形态和农艺性状变异的初步分析. 草地学报, 2015, 23(5): 1048-1056.
[5] Luo T Q, Mo B T, Long Z F. A study on the management and utilization of artificial sward ofBromuscatharticusin winter. Prataculture & Animal Husbandry, 2000, (2): 18-23.
罗天琼, 莫本田, 龙忠富. 优良冬季青绿禾草扁穗雀麦在人工草地中的建植及利用研究. 草业与畜牧, 2000, (2): 18-23.
[6] Fu X N, Wang Z, Gao H W,etal. Morphologic variation of importedDactylisglomerata. Acta Agrestia Sinica, 2012, 20(2): 358-362.
富新年, 王赞, 高洪文, 等. 国外引进鸭茅种质形态变异研究. 草地学报, 2012, 20(2): 358-362.
[7] Sun M, Fu K X, Fan Y,etal. Analysis of phenotypic variations in 15 introduced elite germplasm ofLoliummultiflorumLam. Journal of Plant Genetic Resources, 2016, 17(4): 655-662.
孙铭, 符开欣, 范彦, 等. 15份多花黑麦草优良引进种质的表型变异分析. 植物遗传资源学报, 2016, 17(4): 655-662.
[8] Wang Y X, Zhang B. Phenotypic variation ofTrifoliumrepensin Xinjiang. Acta Agrestia Sinica, 2012, 20(6): 1163-1168.
王玉祥, 张博. 新疆野生白三叶表型性状变异研究. 草地学报, 2012, 20(6): 1163-1168.
[9] Mao S X, Feng X Q. The basis of quantitative genetics in cropsⅱ. Heritability and its estimation. Hereditas, 1979, (4): 42-47.
毛盛贤, 冯新芹. 作物数量遗传学基础——二、遗传力及其估算. 遗传, 1979, (4): 42-47.
[10] Chen Y N. Estimation of heritability, genetic advance and genetic correlation of main characters in sugarbeet varieties. Sugar Crops of China, 1983, (4): 9-13.
陈煜南. 甜菜品种主要性状遗传力、遗传进度和遗传相关的估算. 中国糖料, 1983, (4): 9-13.
[11] Publishing S. SAS 9.2 SQL Procedure User’s Guide. Cary, North Carolina: SAS Publishing, 2008: 995-1014.
[12] Rohlf F J. NTSYS-PC: Numerical Taxonomy and Multivariate Analysis System. version 2.2. New York: Setauket, 2000.
[13] Syukur M, Agronomy D O. Estimation of genetic parameter for quantitative characters of pepper (CapsicumannuumL.). Journal of Tropical Crop Science, 2014, 1(1): 4-7.
[14] Allard R W. Principles of Plant Breeding. 6th edition. New York: Willy, 1999.
[15] Johnson H W, Robinson H F, Comstock R E. Estimates of genetic and environmental variability in soybeans. Agronomy Journal, 1955, 47(7):314-318.
[16] Deshmukh S N, Basu M S, Reddy P S. Genetic variability, character association and path coefficients of quantitative traits inVirginiabunchvarieties of groundnut. Revrvore, 2013, 37(1): 175-180.
[17] Singh B. Plant Breeding: Principles and Methods. 6th edition. New Delhi, India: Kalyani Publishers, 2001.
[18] Luo H Y, Zhou Y L, Hu X Y,etal. Genetic study on several important quantitative traits of wheat. Chinese Agricultural Science Bulletin, 2006, 22(7): 139-141.
罗怀勇, 周玉琳, 胡学运, 等. 小麦几个重要数量性状的遗传分析. 中国农学通报, 2006, 22(7): 139-141.
[19] Gao Z R. Quantitative Genetics. Chengdu: Sichuan University Press, 1986.
高之仁. 数量遗传学. 成都: 四川大学出版社, 1986.
[20] Ai S L. Study on heritability and genetic advance of main economic characters in flue-cured tobacco. Chinese Tobacco Science, 1981, (4): 14-17.
艾树理. 烤烟主要经济性状的遗传力及遗传进度的研究. 中国烟草科学, 1981, (4): 14-17.
[21] Liu Y H, Leng R, Zhang Z R,etal. Preliminary study on heritability and hereditary progress of the main quantitative characters in tumorous stem mustard (Brassicajunceavar.tumidaTsen et Lee). Journal of Plant Genetic Resources, 2006, 7(4): 442-444.
刘义华, 冷蓉, 张召荣, 等. 茎瘤芥主要数量性状遗传力和遗传进度的初步研究. 植物遗传资源学报, 2006, 7(4): 442-444.
