低温处理下外源水杨酸和脱落酸与内生真菌互作对醉马草共生体的影响
2018-01-19刘静陈振江李秀璋周景乐柳莉李春杰
刘静,陈振江,李秀璋,周景乐,柳莉,李春杰
(草地农业生态系统国家重点实验室,农业部草牧业创新重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020)
醉马草(Achnatheruminebrians)是禾本科芨芨草属多年生草本植物,是我国西北天然草场主要的烈性毒草之一;主要分布在我国甘肃,内蒙古,青海,西藏等地[1],且调查发现,这几个地方醉马草内生真菌的带菌率都很高,近100%[2-3]。禾草内生真菌是指在禾草体内度过大部分或者全部生命周期,但却不会引起禾草外部显示任何病害症状的一大类真菌[4]。大量研究表明,内生真菌的侵染能显著提高醉马草的耐盐性[5]、耐旱性[6]、抗病性[6-7]和抗虫性[8]等。但有关内生真菌提高宿主抗寒性的报道不多见。
低温是影响植物生长发育主要的非生物因素之一。研究表明低温条件下种子萌发率降低、幼苗生长缓慢[9]和植物的膜系统发生变化,从而影响脂肪酸的组分与膜的流动性和稳定性[10-11]。此外,低温对植物生理生化的影响主要是影响光合作用[12-14]、酶活性[15-17]和细胞中渗透调节物质等[18-19]。近年来有关外源激素与植物抗寒性的报道很多[20-21]。激素种类繁多,对植物的作用也各不相同,抗寒性方面研究较多的激素为脱落酸(abscisic acid, ABA)[22]和水杨酸(salicylic acid, SA)[23]。ABA具有抑制种子萌发、促进胚胎成熟、叶脱落和衰老等生理作用[24],在植物低温、干旱、盐渍等胁迫中有着重要的作用[25-27],植物处于低温条件时,外施ABA可提高可溶性糖的含量、SOD和POD活性,从而提高植物的耐寒水平[28]。陈善娜等[29]用5~10 mol·L-1的ABA喷施低温胁迫2~4 d的水稻(Oryzasativa)两叶期幼苗,结果显示,清除自由基的超氧化物歧化酶活性和过氧化物酶活性增加,丙二醛含量减少,从而说明外源ABA能明显增强光下水稻幼苗自由基清除能力,加强水稻的耐寒性。SA即邻经基苯甲酸,是植物体内普遍存在的一类小分子物质,在植物的生长发育过程中起着重要的调节作用,如参与植物种子萌发、气孔开闭、光合和呼吸等作用[30]。另外,有研究表明,植物在遭受低温胁迫时,外源SA预处理可提高养心菜(Sedumaizoon)和沟叶结缕草(Zoysiamatrella)中的抗氧化酶活性,增强他们对低温胁迫的抵抗[31-32]。然而有关低温条件下,内生真菌和外源激素互作对醉马草生理、生化的影响尚未见报道。因此,本研究以醉马草幼苗为研究对象,喷洒浓度为1.5 mmol·L-1的SA和0.1 mmol·L-1的ABA,研究其对Chl含量、MDA含量、保护性酶活性、渗透调节物质含量的影响,探讨外源激素与内生真菌互作对醉马草幼苗低温下生长的影响,初步为醉马草低温生长的研究提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1种子和外源激素来源 带内生真菌(E+)和不带内生真菌(E-)醉马草种子均采集于兰州大学草地农业科技学院榆中田间试验地(E 104°08′,N 35°56′,海拔1514 m)。
水杨酸(SA)和脱落酸(ABA)由上海中秦化学试剂有限公司和天津光复精细化工研究所分别提供。
1.1.2幼苗的种植 按照李春杰等[33]的检测方法将上述收集到的E+、E-醉马草种子进行进一步检测,验证种子带内生真菌的情况;先用75%的酒精表面消毒5 min,再用1%次氯酸钠消毒10 min,最后用无菌水冲洗干净,在双层滤纸上进行发芽,出芽2 d后将长势均匀的幼苗种植于16 cm (上口径)×12 cm (下口径)×10 cm (高)的花盆中,每盆5株,然后置于25 ℃光暗交替培养箱中进行生长,一个月后开始处理。
1.2 方法
1.2.1激素处理 幼苗生长1个月后,抽样进行内生真菌检测,再次确定带菌情况,参照前期研究基础[34],用浓度为1.5 mmol·L-1的SA和0.1 mmol·L-1的ABA分别喷施于醉马草E+、E-幼苗,分别标记为SA+、SA-、ABA+和ABA-,用无菌水喷施醉马草E+、E-幼苗作为对照,标记为CK+、CK-,每组设8个重复,常温下生长3 d后再喷洒一次,然后将幼苗置于5 ℃光暗交替培养箱中生长,从第0天开始,每隔2 d取一次样测定生理指标,共取样5次。
1.2.2各项生理指标的测定 按照李合生[35]《植物生理生化实验原理与技术》进行生理生化指标的测定,叶绿素(chlorophyll, Chl)含量采用丙酮浸提法、超氧化物歧化酶(superoxide dismutase, SOD)活性采用氮蓝四唑法、过氧化物酶(peroxidase, POD)活性采用愈创木酚法、脯氨酸(proline acid, Pro)含量采用磺基水杨酸法、丙二醛(malondialdehyde, MAD)含量采用硫代巴比妥酸法、可溶性糖(soluble sugar, SS)含量采用蒽酮法。
1.3 数据处理
采用Microsof Excel 2013录入数据并作图,采用SPSS 17.0对外源激素与内生真菌互作对醉马草幼苗叶绿素含量、丙二醛含量、保护性酶活性和渗透调节物质的含量分别进行双因素方差分析(ANOVA),对同一胁迫时间内不同处理间的差异性进行单因素方差分析。
2 结果与分析
2.1 不同外源激素和内生真菌互作对醉马草幼苗Chl含量的影响
随胁迫时间的延长,醉马草幼苗Chl含量整体呈下降的趋势,且与CK比较,外源激素处理下Chl含量高于CK。CK处理下,E+的Chl含量在胁迫第6天和第8天时显著(P<0.05)高于E-(图1),分别显著提高了38.1%和22.4%。SA处理下,胁迫第6天和第8天时,SA+、SA-的Chl含量均达到显著(P<0.05)水平(图1),SA+比SA-的Chl含量分别显著(P<0.05)提高了23.0%和19.7%。ABA处理下,除第0天外,胁迫第2,4,6和8天时,ABA+的Chl含量显著(P<0.05)高于ABA-(图1),分别显著提高了32.2%,31.1%,32.6%和15.2%。
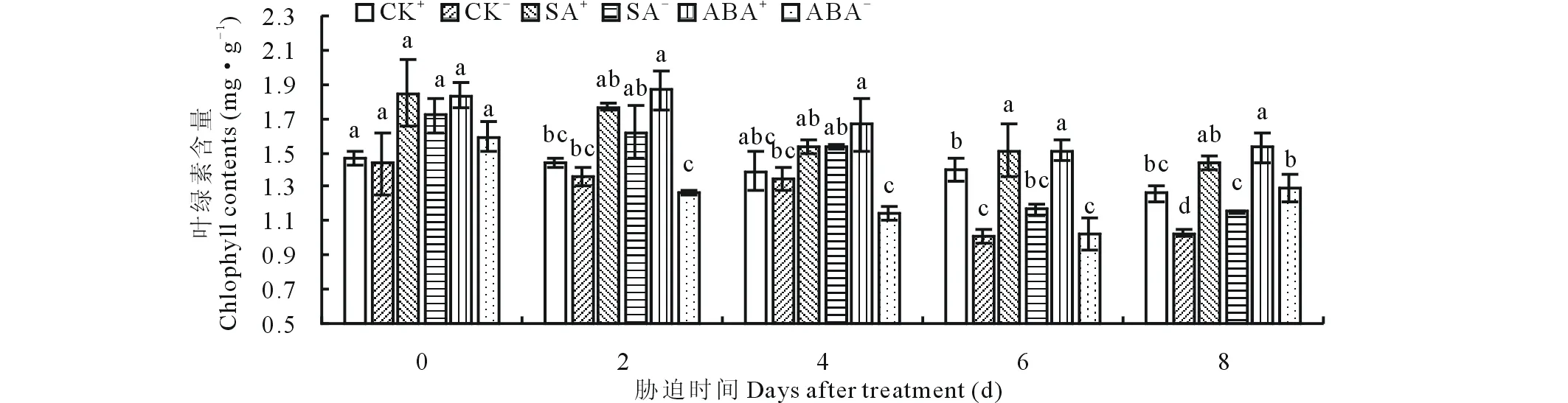
图1 不同激素处理下叶绿素含量随时间的变化Fig.1 The change of chlorophyll content when treated with different hormones 不同字母代表同一胁迫时间内不同处理间的差异显著性(P<0.05),CK+:带内生真菌幼苗的空白对照;CK-:不带内生真菌幼苗的空白对照;SA+:水杨酸处理下带内生真菌的幼苗;SA-:水杨酸处理下不带内生真菌的幼苗;ABA+:脱落酸处理下带内生真菌的幼苗;ABA-:脱落酸处理下不带内生真菌的幼苗,下同。Different letters mean significant difference (P<0.05) among different treatments, CK+: Blank control of endophyte-infected seedling;CK-: Blank control of endophyte-free seedling;SA+: Endophyte-infected seedling were treated by salicylic acid;SA-:Endophyte-free seedling were treated by salicylic acid;ABA+: Endophyte-infected seedling were treated by abscisic acid;ABA-: Endophyte-free seedling were treated by abscisic acid. The same below.
2.2 不同外源激素和内生真菌互作对醉马草幼苗MDA含量的影响
SA或ABA处理下,随胁迫时间的延长,醉马草幼苗MDA含量均呈上升趋势,且与CK比较,外源激素处理条件下幼苗MDA含量整体低于CK(图2)。CK处理下,胁迫第6天时,E+、E-的MDA含量均达到显著(P<0.05)水平(图2)。SA处理下,胁迫第6天和第8天时,SA+、SA-的MDA含量达到显著(P<0.05)水平(图2),SA+的MDA含量比SA-分别显著(P<0.05)降低了25.4%和19.4%。ABA处理下,胁迫第6天和第8天时,ABA-的MDA含量比ABA+分别显著(P<0.05)提高了21.1%和34.3%。
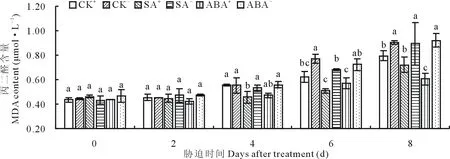
图2 不同激素处理下MDA含量随时间的变化Fig.2 The change of MDA content when treated with different hormones
2.3 不同外源激素和内生真菌互作对醉马草幼苗渗透调节物的影响
2.3.1不同外源激素和内生真菌互作对醉马草幼苗Pro含量的影响 随胁迫时间的延长,醉马草幼苗Pro含量均呈先升高后降低又升高的趋势,且与CK比较,外源激素处理下幼苗Pro含量高于CK。CK处理下,胁迫第2,6,8天时,E+、E-的Pro含量达到显著(P<0.05)水平(图3),E+的Pro含量显著(P<0.05)高于E-,分别提高32.8%,44.2%和23.6%。SA处理下,随胁迫时间延长,SA+、SA-的Pro含量均达到显著(P<0.05)水平(图3),SA与E+比SA 与E-相互作用效果明显,SA+的Pro含量比SA-分别显著(P<0.05)高20.3%,42.9%,51.8%,47.6%和18.2%。ABA处理下,除胁迫第2天外,各时间段ABA+的Pro含量均显著(P<0.05)高于ABA-(图3),且分别提高了37.9%,54.7%,28.1%和26.1%。
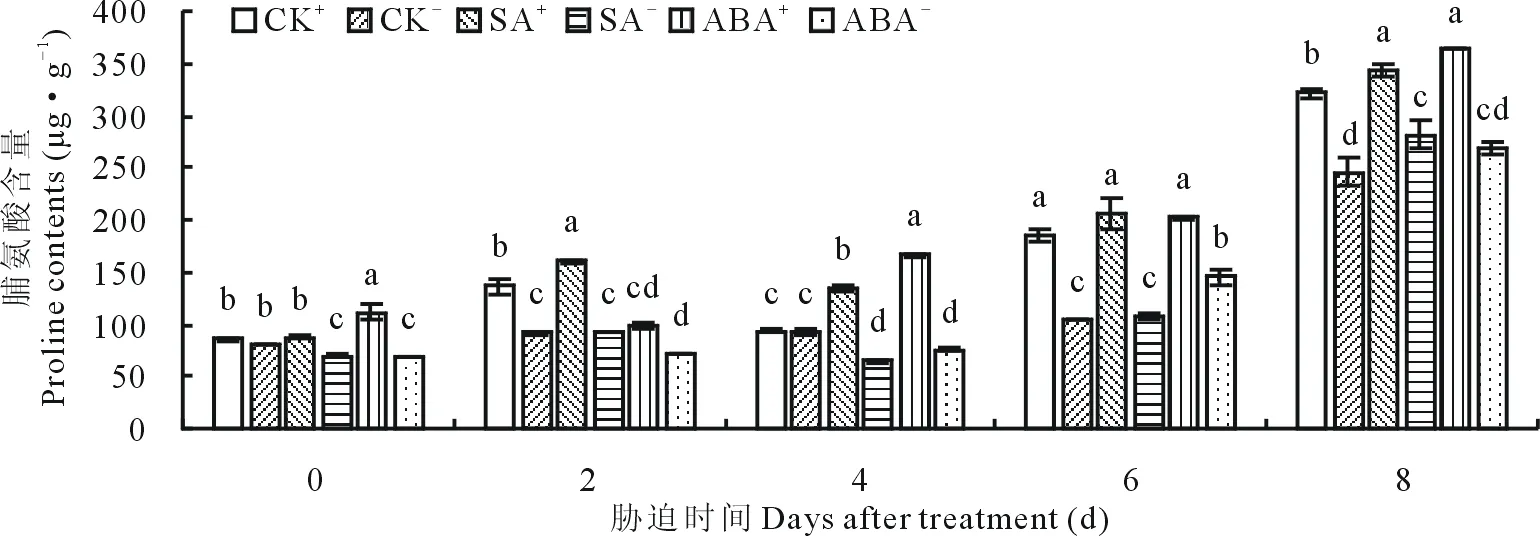
图3 不同激素处理下脯氨酸含量随时间的变化Fig.3 The change of proline contents when treated with different hormones
2.3.2不同外源激素和内生真菌互作对醉马草幼苗SS含量的影响 随胁迫时间的延长,醉马草幼苗SS含量总体呈先升高后降低的趋势,且与CK比较,一定胁迫时间内,外源激素处理下幼苗SS含量高于CK。CK处理下,胁迫第6天时,CK的SS含量达到显著(P<0.05)水平(图4),CK+的SS含量比CK-显著(P<0.05)高26.5%。SA处理下,胁迫第6天和第8天时,SA+的SS含量显著(P<0.05)高于SA-(图4),分别提高了23.7%和15.3%。ABA处理下,胁迫第6天和第8天时,ABA+、ABA-的SS含量达到显著(P<0.05)水平(图4),ABA+的SS含量比ABA-分别显著提高了9.1%和11.4%。

图4 不同激素处理下可溶性糖含量随时间的变化Fig.4 The change of soluble sugar contents when treated with different hormones
2.4 不同外源激素和内生真菌互作对醉马草幼苗保护性酶的影响
2.4.1不同外源激素和内生真菌互作对醉马草幼苗SOD活性的影响 随胁迫时间的延长,醉马草幼苗SOD活性总体呈先升高后降低的趋势,且与CK比较,一定胁迫时间内,外源激素处理下幼苗SOD含量高于CK。CK处理下,各胁迫时间段E+幼苗SOD活性均显著(P<0.05) 高于E-(图5)。SA处理下,除第0天外,随着胁迫时间的延长,各时间段SA+、SA-的SOD活性均达到显著(P<0.05)水平(图5),SA+的SOD活性比SA-分别显著(P<0.05)高出39.0%,62.1%,10.2%和44.8%。ABA处理下,除胁迫第4天外,其他各胁迫时间段ABA+的SOD活性均显著(P<0.05)高于ABA-(图5),ABA提高了E+幼苗的SOD活性,比E-分别显著(P<0.05)提高了17.6%,53.2%,10.1%和15.0%。
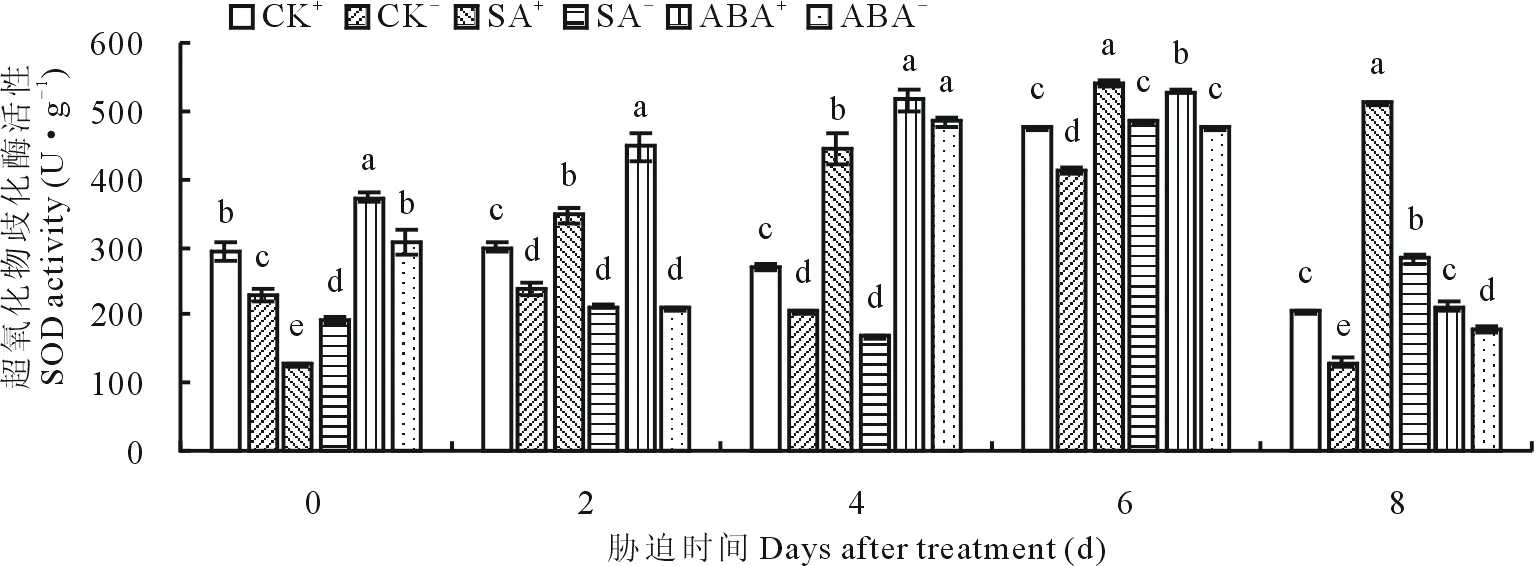
图5 不同激素处理下SOD活性随时间的变化Fig.5 The change of SOD activity when treated with different hormones
2.4.2不同外源激素和内生真菌互作对醉马草幼苗POD活性的影响 随胁迫时间的延长,醉马草幼苗POD活性总体呈先升后降的趋势,且与CK比较,一定胁迫时间内,外源激素处理条件下幼苗POD活性高于CK。CK处理下,胁迫第0天和第4天时E+、E-的POD活性达到显著(P<0.05)水平(图6), E+的POD活性比E-分别显著(P<0.05)提高了18.0%和16.2%。SA处理下,胁迫第4天和第6天时,SA+、SA-的POD活性达到显著(P<0.05)水平(图6),SA+的POD活性比SA-分别显著(P<0.05)高出14.6%和18.0%。ABA处理下,胁迫第0,4,6天时,ABA+的POD活性显著(P<0.05)高于ABA-(图6),分别高出24.8%,33.0%和17.9%。

图6 不同激素处理下POD活性随时间的变化Fig.6 The change of POD activity when treated with different hormones
内生真菌、激素、时间单独作用和激素与时间、激素与内生真菌、内生真菌与时间及三者互作均对Pro含量和SOD活性有显著性(P<0.05)的影响(表1)。
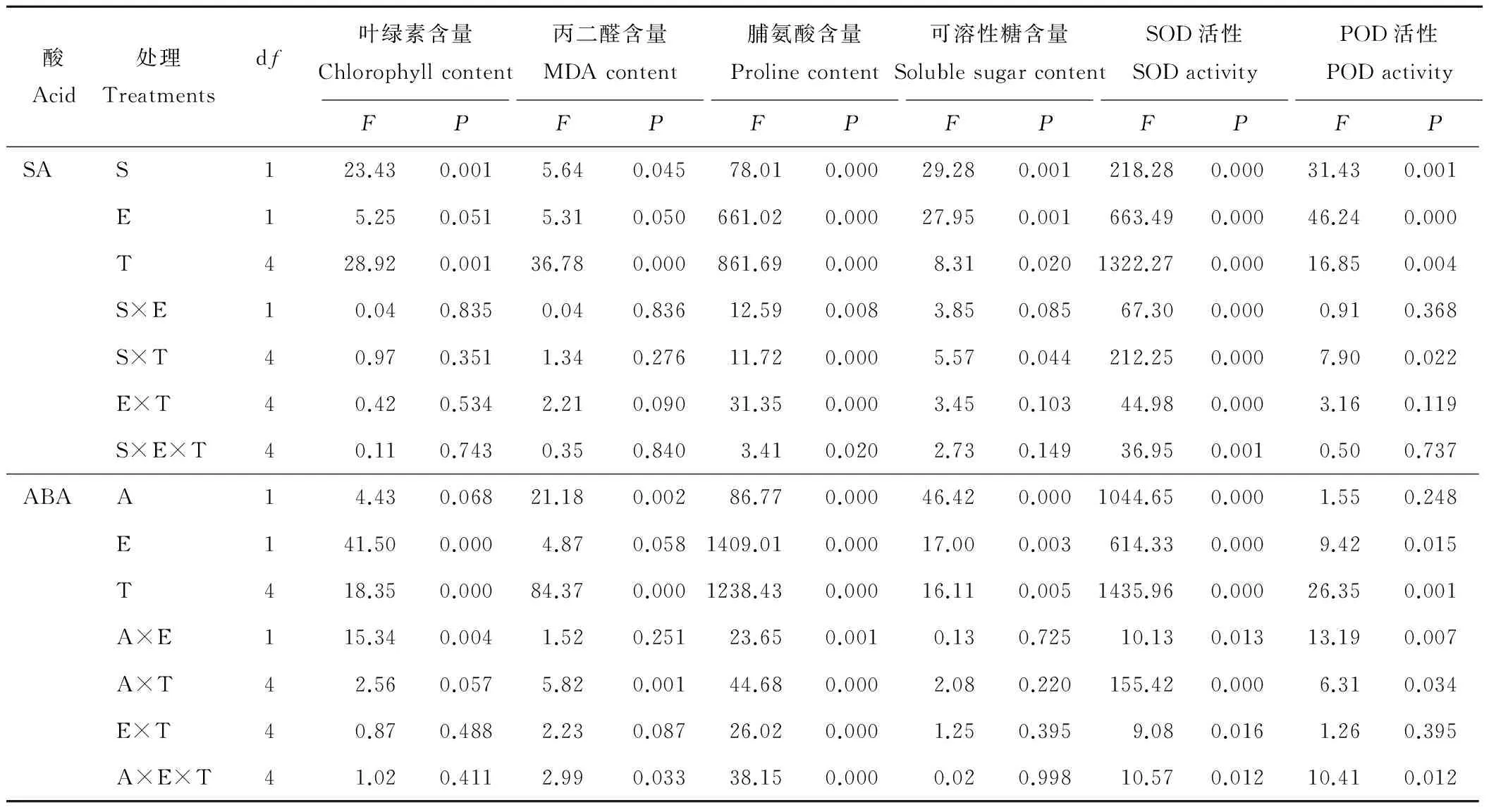
表1 SA(S)、ABA(A)和内生真菌(E)在不同的时间(T)对5 ℃低温胁迫下的醉马草CHL、MDA、Pro、SS、SOD和POD的重复度量方差分析结果Table 1 Results of repeated measures ANOVA for the effects of chlorophyll content, MDA content, proline content, soluble sugar content, SOD activity and POD activity of A. inebrians under 5 ℃
S: SA; E: Endophyte; T: Time; A: ABA.
3 讨论与结论
有研究表明Neotyphodium内生真菌侵染可以增加叶绿素和可溶性糖的含量,从而提高宿主中华羊茅(Festucasinensis)的抗寒性[36]。杨洋[37]研究发现,无论在实验室、盆栽与田间试验,带内生真菌的中华羊茅SOD、POD活性高于不带内生真菌的中华羊茅,且带有内生真菌的中华羊茅对可溶性糖和脯氨酸的积累能力显著强于不带菌的植株。本研究表明,在CK、SA和ABA处理下,E+幼苗Chl含量、保护性酶活性和渗透调节物质含量均高于E-幼苗,说明内生真菌对醉马草幼苗在低温胁迫下起到了一定的缓解作用。MDA含量是植物细胞膜脂过氧化程度的体现,它含量的高低可作为考察植物细胞受胁迫严重程度的指标之一,MDA含量越高说明植物体受损越严重,抗逆性越差,反之则抗逆性较强。在CK、SA和ABA处理下,随着低温胁迫时间的延长,E+、E-的MDA含量均呈升高趋势,且E+的增加程度显著(P<0.05)低于E-,这表明E+抵抗低温的能力高于E-。
冷害作为环境胁迫的一个重要因子,直接影响植物的生长和发育。植物幼苗长期受低温胁迫首先会表现出叶片失绿和萎蔫现象,导致其生长迟缓,光合作用减弱。用ABA处理小麦(Triticumaestivum)幼苗,无论短期还是长期处理,ABA都能不同程度提高小麦幼苗的光合能力、抗氧化酶活性等指标,从而增强对环境胁迫的抗性[38]。在低温胁迫时,喷施一定浓度的SA可以提高香蕉(Musanana)幼苗的光合能力,从而增强香蕉幼苗的抗寒性[39]。本研究中,与对照相比较,SA和ABA处理下,叶绿素含量均高于CK。SA处理下,在胁迫的第6天和第8天时E+的Chl含量显著高于(P<0.05)E-,而ABA处理下,从第2天开始就达到显著水平,说明SA和ABA均可以提高植物的抗寒性,只是各自表达的时间不一致。主要是因为ABA属于激素类物质,在寒冷胁迫下,脱落酸启动细胞抗冷基因的表达,诱导植物产生抗寒能力。SA是非激素类有机物质,在植物体内通过传导作用提高植物的抗逆能力[20]。植物处于冷害胁迫下,也会导致活性氧清除系统活性降低,细胞膜脂化程度降低、抗逆能力下降等一系列外部及内部变化[12,40],施用外源激素SA和ABA可以增强植物的抗寒性,在多种植物上都表现为保护性酶活性提高、渗透调节物质含量的增加和MDA含量的下降[41-45]等,本试验对低温下外源SA和ABA处理醉马草抗寒性的研究也得到了相同的实验结果。
MDA是膜脂氧化反应的最终产物之一,它的产生能加剧膜的损伤,植物受到逆境伤害时,植物体内MDA含量会升高,导致植物细胞膜系统受到伤害。本试验结果表明,在CK、SA和ABA处理下,随胁迫时间的延长,醉马草幼苗MDA含量均呈上升趋势,在胁迫的第6天和第8天时,E+与E-的MDA含量均达到显著水平,且SA和ABA处理下,MDA含量显著(P<0.05)低于CK。说明低温处理下,随胁迫时间的变化醉马草幼苗细胞膜膜脂过氧化的程度越大,对植株的伤害越严重,而SA和ABA处理减弱膜脂过氧化产物MDA的积累。此结果与魏安智等[28]和徐冬梅[31]的研究结果一致。游离脯氨酸和可溶性糖作为主要的渗透调节物质,具有清除活性氧和稳定细胞膜结构的作用,从而起到抗寒作用[46]。本研究表明,在SA和ABA处理下,醉马草幼苗Pro和SS含量在胁迫的不同时间段均高于CK。是因为植物体内脯氨酸含量在一定程度上反映了植物的抗逆性,植物在遭受逆境胁迫时体内脯氨酸含量会显著增加[47]。可溶性糖是植物抗逆作用中一个重要的渗透调节因子,也是植物体中重要的能源和碳源,植物遭受低温胁迫时常常会积累大量的可溶性糖,含量越多,表明植物抗寒的能力越强[48]。本研究结果表明,SA和ABA处理后进一步提高了低温胁迫下醉马草体内渗透调节物质Pro的含量和SS的积累。此结果与李冬花[44]研究外源ABA和SA对低温胁迫下圆柏属植物幼苗生理特性时也得到了类似的结果。活性氧积累是引起质膜受损的重要原因,而低温胁迫能造成细胞内活性氧积累。提高抗氧化酶活性与抗氧化剂含量,清除积累的活性氧是提高植物抗寒性的主要途径之一[25]。SOD是植物细胞内的一类重要保护酶,可以保护细胞免受氧化损伤,是活性氧清除剂之一,能将超氧化物阴离子自由基快速氧化为 H2O 和 O2,保护细胞膜的结构和功能不受到伤害; POD 合成以清除活性氧自由基的积累,这是细胞酶保护系统的一种应激机制[21]。在本研究结果中,除胁迫当天,与对照相比,喷施外源激素的醉马草幼苗无论是E+还是E-的SOD和POD活性均显著高于(P<0.05)对照处理,说明SA和ABA处理减弱了低温胁迫对SOD和POD活性的影响,降低了自由基的伤害,从而增加了抗寒能力。此结果与李兆亮等[45]和李雪梅等[38]的研究结果一致。但随着胁迫时间的持续,SOD和POD活性降低,幼苗受到的伤害越严重,是因为SOD和POD则是冷害敏感型,这种酶合成的启动时间较早,在受害初期就已经大量合成,这也是植物自身膜保护系统的调控机制。此结果与刘海卿等[41]的在对不同叶龄期叶面喷施ABA对北方白菜型冬油菜抗寒性的研究中已得到证实。
在SA和ABA处理下,SA+和ABA+的Chl和SS含量始终大于CK+,SA+和ABA+的MDA含量始终小于CK+,在胁迫的第4,6,8天时,SA+和ABA+的Pro含量、SOD和POD的活性大于CK+,而在不同胁迫时间段,均有SA-和ABA-的Chl含量、Pro含量、SOD和POD的活性小于CK-。说明ABA和SA与内生真菌互作对低温胁迫下醉马草幼苗的生长起到了保护作用。
References:
[1] Shi Z C. China Grass Important Poisonous Plant. Beijing: China Agriculture Press, 1997.
史志诚. 中国草地重要有毒植物. 北京: 中国农业出版社, 1997.
[2] Nan Z B, Li C J.Neotyphodiumin native grasses in China and observations on endophyte/host interaction//Roberts C A, West C P, Spiers D A.Neotyphodiumin Cool-season Grasses. Ames, Iowa: Blackwell Publishing, 2000: 163-185.
[3] Li C J, Nan Z B, Gao J H. Detection and distribution ofNeotyphium-Achnatheruminebriansassociation in China//Robert K, Charles R K, Ryan L. USA Proceedings of 5th International Neotyphodium/Grass Interactions Symposium, Arkansas, 2004, 210: 24-27.
[4] Schardl C L, Leuchtmann A, Spiering M J. Symbioses of grasses with seed borne fungal endophytes. Annual Review of Plant Biology, 2004, 55(55): 315-340.
[5] Song M, Chai Q, Li X,etal. An asexualEpichloё endophyte modifies the nutrient stoichiometry of wild barley (Hordeumbrevisubulatum) under salt stress. Plant and Soil, 2015, 387(1): 153-165.
[6] West C P, Gwinn K D. Role of Acremonium in drought, pest, and disease tolerances of grasses. Proceedings of the Second International Symposium on Acremonium/Grasses Interational: Plenary papers. Paimerston North, New Zealand, 1993: 11-30.
[7] Li C J, Zhang X X, Li F,etal. Disease and pests resistance of endophyte infected and non-infected drunken horse grass//Popay A, Thom E R. Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses. New Zealand Grassland Association, Dunedin, New Zealand, 2007: 111-114.
[8] Bacon C W, Richardson M D, White J F. Modification and uses of endophyte-enhanced turfgrasses: A role for molecular technology. Crop Science, 1997, 37(37): 1415-1425.
[9] Zhang Z Q, Shu C S, Dong X F,etal. Effect of low temperature on seed germination and seedling growth in wheat. Southwest China Journal of Agricultural Sciences, 2010, 23(1): 22-25.
张泽全, 舒长生, 董雪芳, 等. 低温对小麦种子萌发和幼苗生长的影响. 西南农业学报, 2010, 23(1): 22-25.
[10] Deng L Y, Wang H C. Cold hardiness of grape in relation to membrane lipid composition. Journal of Plant Physiology, 1982, 8(3): 273-283.
邓令毅, 王洪春. 葡萄的膜脂和脂肪酸组分与抗寒性关系的研究. 植物生理学报, 1982, 8(3): 273-283.
[11] Wang X X, Li S D, Dong H R,etal. The effect of low temperature on free fatty acid in tomato seedlings and florescence. Journal of Horticulture, 1997, 24(2): 161-164.
王孝宣, 李树德, 东惠茹, 等. 低温胁迫对番茄苗期和开花期脂肪酸的影响. 园艺学报, 1997, 24(2): 161-164.
[12] He J, Liu H X, Wang Y R,etal. Low temperature and photosynthesis of plants. Plant Physiology Communications, 1986, (2): 3-8.
何洁, 刘鸿先, 王以柔, 等. 低温与植物的光合作用. 植物生理学报, 1986, (2): 3-8.
[13] Dai Y H, Liu X Y, Meng Q W,etal. Effect of low temperature on lipid metabolism of thylakoid membrane. Chinese Bulletin of Botany, 2004, 21(4): 506-511.
代玉华, 刘训言, 孟庆伟, 等. 低温胁迫对类囊体膜脂代谢的影响. 植物学通报, 2004, 21(4): 506-511.
[14] Duan W, Li X G, Meng Q W,etal. Mechanism of plant light suppression at low temperature. Journal of Northwest Botanical Sciences, 2003, 23(6): 1017-1023.
段伟, 李新国, 孟庆伟, 等. 低温下的植物光抑制机理. 西北植物学报, 2003, 23(6): 1017-1023.
[15] Fryer M J, Andrews J R, Oxborough K,etal. Relationship between CO2assimilation, photosynthetic electron transport, and active O2metabolism in leaves of maize in the field during periods of low temperature. Plant Physiology, 1998, 116(2): 571-580.
[16] Prasad T K. Role of catalase in inducing chilling tolerance in pre-emergent maize seedlings. Plant Physiology, 1997, 114(4): 1369-1376.
[17] Van Breusegem F, Slooten L, Stassart J M,etal. Effects of overproduction of tobacco MnSOD in maize chloroplasts on foliar tolerance to cold and oxidative stress. Journal of Experimental Botany, 1999, 50(330): 71-78.
[18] Delauney A J, Verma D P S. Proline biosynthesis and osmoregulation in plants. The Plant Journal, 993, 4(2): 215-223.
[19] Wang M L. Plant osmotic stress and its relationship with proline. Beijing Garden, 2006, 22(2): 21-24.
王茂良. 植物抗渗透胁迫及其与脯氨酸的关系. 北京园林, 2006, 22(2): 21-24.
[20] Sun Y J, Wang G H. Effects of exogenous growth regulators on cold resistance of plant. Crop Research, 2011, 25(3): 287-291.
孙玉洁, 王国槐. 外源生长调节剂对植物抗寒性的影响. 作物研究, 2011, 25(3): 287-291.
[21] Wu J Y, Liu H Q, Sun W C,etal. Exogenous ABA effect on physiology of winter rapeseed (BrassicarapeL.) seedling in northern China. Chinese Journal of Oil Crop Sciences, 2015, 37(3): 310-315.
武军艳, 刘海卿, 孙万仓, 等. 苗期喷施外源脱落酸对北方白菜型冬油菜越冬生理的影响. 中国油料作物学报, 2015, 37(3): 310-315.
[22] Chen T H, Gusta L V. Abscisic acid-induced freezing resistance in cultured plant cells. Plant Physiology, 1983, 73(1): 71-75.
[23] Janda T, Szalai G, Tari I,etal. Exogenous salicylic acid has an effect on chilling symptoms in maize (ZeamaysL.) plants//Sowinski P, Zagdanska B, Aniol A,etal. Crop Development for the Cool and Wet Regions in Europe. EC, Luxembourg. ISBN 92-828-1810-1. 1997: 179-187.
[24] Wasilewska A, Vlad F, Sirichandra C,etal. An update on abscisic acid signaling in plants and more. Molecular Plant, 2008, 1(2): 198-217.
[25] Liu D B, Wei J Y, Cui B M,etal. Effects of ABA on cold resistance of banana seedlings. Chinese Journal of Tropical Crops, 2007, 28(2): 1-4.
刘德兵, 魏军亚, 崔百明, 等. 脱落酸对香蕉幼苗抗寒性的影响. 热带作物学报, 2007, 28(2): 1-4.
[26] Wei X, Ni H, Zhang H H,etal. Effects of exogenous abscisic acid and brassinolide on drought resistance of soybean seedlings. Chinese Journal of Oil Crop Sciences, 2016, 38(5): 605-610.
魏鑫, 倪虹, 张会慧, 等. 外源脱落酸和油菜素内酯对干旱胁迫下大豆幼苗抗旱性的影响. 中国油料作物学报, 2016, 38(5): 605-610.
[27] Zhang Y, Xu X, Zhu Y X,etal. Progress of mechanisms of ABA response to plant salt stress. Agricultural Science Bulletin, 2015, (24): 143-148.
张岩, 许兴, 朱永兴, 等. ABA响应植物盐胁迫的机制研究进展. 中国农学通报, 2015, (24): 143-148.
[28] Wei A Z, Yang T X, Zhang R,etal. Effect of exogenous ABA on cold-resistant ability and related physiological indicators of almond-apricot during blossom period. Journal of Northwest A & F University (Naturnal Science Edition), 2008, 36(5): 79-84.
魏安智, 杨途熙, 张睿, 等. 外源ABA对仁用杏花期抗寒力及相关生理指标的影响. 西北农林科技大学学报(自然科学版), 2008, 36(5): 79-84.
[29] Chen S N, Guo Z H, Shen Y G,etal. The effect of exogenous ABA on the scavenging systems of oxygen free radical of plateau rice seedlings in chilling stress. Journal of Yunnan University (Naturnal Science Edition), 1996, 18(2): 167-172.
陈善娜, 郭浙红, 沈云光, 等. 在低温胁迫下外源脱落酸对高原水稻自由基清除系统的影响. 云南大学学报(自然科学版), 1996, 18(2): 167-172.
[30] Yuan Y B, Cao Z Z. The role of salicylic acid in plants. Chinese Bulletin of Botany, 1994, 11(3): 1-9.
原永兵, 曹宗巽. 水杨酸在植物体内的作用. 植物学通报, 1994, 11(3): 1-9.
[31] Xu D M. Effect of AMF and Exogenous Salicylic Acid on Cold Resistance of theSedumaizoonL. Ya’an: Sichuan Agricultural University, 2015.
徐冬梅. AMF和外源SA对养心菜耐寒性的影响. 雅安: 四川农业大学, 2015.
[32] Wang Y, Li J L, Jiang T,etal. Effect of pre-treatments with SA, H2O2and 6-BA on chilling tolerance inZoysiamatrella. Acta Prataculturae Sinica, 2010, 19(2): 76-81.
王艳, 李建龙, 姜涛, 等. SA、H2O2和6-BA预处理对沟叶结缕草耐寒性的影响. 草业学报, 2010, 19(2): 76-81.
[33] Li C J, Nan Z B, Liu Y. Methodology of edophyte detection of durnken horse grass (Achnatheruminebrians). Edible Fungi of China, 2008, 27(Suppl 1): 16-19.
李春杰, 南志标, 刘勇. 醉马草内生真菌检测方法的研究. 中国食用菌, 2008, 27(增刊1): 16-19.
[34] Liu L, Li X Z, Chen Z J,etal. Effects of interaction between exogenous hormones andEpichloё on germination of drunken horse grass (Achnatheruminebrians) under low temperature stress. Journal of Southwest University for Nationalities (Natural Science Edition), 2016, 42(4): 373-382.
柳莉, 李秀璋, 陈振江, 等. 外源激素与内生真菌互作对醉马草低温胁迫下种子萌发的影响. 西南民族大学学报(自然科学版), 2016, 42(4): 373-382.
[35] Li H S. Principles and Techniques of Plant Physiology and Biochemistry Experiment (for the twenty-first century course). Beijing: Higher Education Press, 2000.
李合生. 植物生理生化实验原理与技术(面向21世纪课程教材). 北京: 高等教育出版社, 2000.
[36] Peng Q Q. Effect ofNeotyphodiumEndophyte on Chilling Tolerance toFestucasinensis. Lanzhou: Lanzhou University, 2012.
彭清青.Neotyphodium内生真菌对中华羊茅耐寒性的影响. 兰州: 兰州大学, 2012.
[37] Yang Y.NeotyphodiumEndophyte inFestucasinensisand Effect of Cold Tolerance to Host. Lanzhou: Lanzhou University, 2010.
杨洋. 中华羊茅内生真菌及其对宿主抗寒性的影响. 兰州: 兰州大学, 2010.
[38] Li X M, Chen Q, Wang L L,etal. Effects of abscisic acid on photosynthesis and antioxidant enzymes in wheat seedling. Journal of Shenyang Normal University(Natural Science Edition), 2006, 24(2): 221-223.
李雪梅, 陈强, 王兰兰, 等. 脱落酸对小麦幼苗光合及抗氧化酶的影响. 沈阳师范大学学报(自然科学版), 2006, 24(2): 221-223.
[39] Kang G Z, Wang G C. Participation of H2O2in enhancement of cold chilling by salicylic acid in banana seedlings. Botany of Journal, 2003, 45(5): 567-573.
[40] Yu F Y, Xu T Z.A review on plant stress physiology. World Forestry Research, 2003, 16(5): 6-11.
喻方圆, 徐锡增. 植物逆境生理研究进展. 世界林业研究, 2003, 16(5): 6-11.
[41] Liu H Q, Wu J Y, Sun W C,etal. Effect of exogenous abscisic acid on the cold resistance ofBrassicarapacv. Longyou (winter rape) 8 cultivated at different foliar in northern China. Acta Prataculturae Sinica, 2015, 24(9): 173-180.
刘海卿, 武军艳, 孙万仓, 等. 不同叶龄期叶面喷施ABA对北方白菜型冬油菜抗寒性的影响. 草业学报, 2015, 24(9): 173-180.
[42] Fang Y, Wu J Y, Sun W C,etal. Inducing effects of exogenous ABA on seed germination and cold tolerance of winter rape seedlings. Agricultural Research in the Arid Areas, 2014, 32(6): 70-74.
方彦, 武军艳, 孙万仓, 等. 外源 ABA 浸种对冬油菜种子萌发及幼苗抗寒性的诱导效应.干旱地区农业研究, 2014, 32(6):70-74.
[43] Bao Z J. Research on the cold resistance of soybean seedlings processed by abscisic acid and salicylic acid. Journal of Baicheng Normal University, 2015, 29(11): 37-42.
鲍智娟. 脱落酸和水杨酸处理大豆对其幼苗抗冷性影响的研究. 白城师范学院学报, 2015, 29(11): 37-42.
[44] Li D H. Effects of Exogenous Abscisic Acid (ABA) and Salicylic Acid (SA) on Physiological Characteristics ofSabinaSeedlings under Low Temperature Stress. Lanzhou: Lanzhou Jiaotong University, 2014.
李冬花. 外源ABA和SA对低温胁迫下圆柏属植物幼苗生理特性的影响. 兰州: 兰州交通大学, 2014.
[45] Li Z L, Yuan Y B, Liu C L,etal. Regulation of antioxidant enzymes by salicylic acid in cucumber leaves. Acta Botanica Sinica, 1998, 40(4): 356-361.
李兆亮, 原永兵, 刘成连, 等. 水杨酸对叶片抗氧化剂酶系的调节作用. 植物学报, 1998, 40(4): 356-361.
[46] Ji Y, Zhang X Q, Peng Y,etal. Effects of drought stress on lipid peroxidation, osmotic adjustment and activities of protective enzymes in the roots and leaves of orchardgrass. Acta Prataculturae Sinica, 2014, 23(3): 144-151.
季杨, 张新全, 彭燕, 等. 干旱胁迫对鸭茅根、叶保护酶活性、渗透物质含量及膜质过氧化作用的影响. 草业学报, 2014, 23(3): 144-151.
[47] Hare P, Cress W, Van Staden J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell & Environment, 1998, 21(6): 535-553.
[48] Yang D G, Wu G X, Tang X L,etal. Research progress on the plant molecular response mechanisms under the low-temperature stress. Journal of Maize Sciences, 2009, 17(2): 99-101.
