混播方式对豆禾混播草地植物根系构型特征的影响
2018-01-19朱亚琼郑伟王祥关正翾
朱亚琼,郑伟,2*,王祥,关正翾
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区草地资源与生态重点实验室,新疆 乌鲁木齐830052)
牧草根系是牧草吸收、运移土壤水分、养分的关键载体[1],让牧草地上部分生长与土壤紧密结合起来[2]。根系构型是指植物根系在生长介质(土壤)中的空间分布与组织结构,包括根系几何形态、拓扑结构[3]和分形特征[4]等。其中,根系几何形态参数包括根长、根系直径、根系生物量和根表面积等;拓扑结构参数主要通过根系的连接数、分叉数、根系分支率和拓扑指数来反映[5];分形可以对自相似和标度不变性结构特征的不规整形体进行定量分析,而重复性分支使得根系呈现一定的自相似性和分形特点[6],利用分形维数、分形丰度分析根系空间的分布形态和分支状况[7],有利于提高定量描述根系形态的可靠性。根系构型决定了根系在土壤空间中的位置、空间拓展能力和资源获取方式[8],是牧草在草地群落中适应环境异质性[9]和种间相互作用[10]的结果。牧草在群落中动态发展、变化过程中,往往需要适时调整其生态适应对策,以提高其适合度。其中根系形态构型特征会随着环境变化进行适应性调整[11-12],表现出可塑性,从而构建与群落中其他牧草和环境相适应的根系水分、养分吸收和传输体系[13]。
在存在种间竞争的豆禾牧草混播复合系统中,禾本科牧草的再生能力和生长速率往往高于豆科牧草,在地上种间竞争中处于强竞争者的地位[14]。而地下根系对养分和水分的竞争过程更为复杂,有的牧草根系空间扩展能力强,占据更大的土壤空间[15];有的牧草向根系分配更多的氮,使根系中细根的比例增加,增加根系与土壤的接触面积,以增加其竞争能力[16];有的则提高单位根系的吸收功能[17];有的则调整根系分支数(分叉数)和连接长度的权衡关系[5,13],实现根系功能性状间的资源权衡。因此,混播复合系统中豆禾牧草根系构型的变化是适应竞争环境,提高竞争效率的有效适应对策[18]。在豆科牧草(作物)与禾本科牧草(作物)间混作系统中,根系研究主要集中在根系几何形态变化对间混作系统的适应方式和对地上产量的贡献[19-23]、根系几何形态变化与氮、磷吸收利用效率的关系[21,24-29]、根系在土壤空间中垂直分布变化[30-31]等方面,对于自然草地生态系统中牧草根系构型对环境的适应对策[4-5,13,32]、退化草地中牧草构件生物量分配模式[33]、影响根系构型的主要功能性状[34]等也做了系统研究。而从根系构型,特别是全面的从根系几何形态、拓扑结构和分形特征的角度出发,对竞争生境中根系构型可塑性及适应对策的研究仍不明确。
与此同时,关于红豆草(Onobrychisviciaefolia)+无芒雀麦(Bromusinermis)型混播草地的研究主要集中在混播比例、施肥、刈割次数及留茬高度等对牧草产量和牧草品质的影响[35]。如郑伟等[36]研究结果表明,含红豆草的混播组合拥有较高的ADF(acid detergent fiber)产量,而不含红豆草的混播组合ADF产量则较低;另在混播种类较少的组合中,红豆草的存在会使该混播组合表现出较好的产量和较强的组分稳定性。也有学者认为,混播种类较少的红豆草与禾本科牧草混播比混播种类多的组合优越[37]。张强强等[38]指出,在新疆伊犁昭苏地区红豆草和无芒雀麦均表现出有很好的适应与竞争能力,二者混播是较为理想的草种组合。但从根系构型特征的角度出发,对红豆草+无芒雀麦型混播草地的地下竞争研究还较少,成为解释该型混播草地混播优势产生机制的重要制约因素。因此,本研究以红豆草与无芒雀麦型混播草地为对象,将种群空间距离(行距)、种群空间作用方式(同行/异行/异行阻隔)作为地下根系竞争环境的变化因素,从牧草根系的几何形态、拓扑结构和分形特征等角度出发,利用盆栽试验,分析和比较2种牧草在竞争生境中根系构型可塑性及适应对策的变化,为从地下根系方面阐明豆禾牧草混播复合系统高效生产机制做出贡献。
1 材料与方法
1.1 试验区概况
试验地点位于新疆伊犁哈萨克自治州昭苏盆地腹内的昭苏马场(81°03′-81°05′ E,42°38′-43°15′ N),该地区属温带山区半湿润半干旱冷凉气候类型[36]。根据国家气象数据网的数据(站点号51437),2013-2015年,研究区年均温4.40 ℃,年最高温均值为29.50 ℃,年最低温均值为-21.50 ℃,年均降水量468.03 mm,生长季(5-10月)降水量均值为352.20 mm,占年降水量的75.25%。
1.2 盆栽试验设计
本试验选用的材料为红豆草,无芒雀麦。试验前从昭苏马场山地草甸取回0~20 cm的土壤,去除根系、凋落物和大石块,混匀,作为盆栽试验基质。昭苏马场山地草甸土壤为黑钙土,土壤有机质含量为13.63%~14.89%,全氮、全磷、全钾量分别为9.53,2.16,12.20 g·kg-1,碱解氮、有效磷、有效钾含量分别为459.20,144.43,473.58 mg·kg-1[36]。
试验设4种播种模式处理,分别为同行混播+15 cm行距(T15),异行混播+15 cm行距(Y15),异行阻隔混播+15 cm行距(中间用木板阻隔YZ15)和异行混播+30 cm行距(Y30),红豆草和无芒雀麦混播比例均为5∶5。单播红豆草和无芒雀麦作为对照,行距均为15 cm。每个花盆均只播种2行,所有处理均为3次重复,共21个花盆;15 cm行距花盆大小为70 cm(长)×30 cm(宽)×30 cm(高),30 cm行距花盆大小为70 cm(长)×45 cm(宽)×30 cm(高),播种时距花盆边缘均为7.5 cm。无芒雀麦单播的播量为30 kg·hm-2,红豆草单播的播量为60 kg·hm-2,混播比例按种子占单播重量的实际用价来计算,混播与单播密度相同。具体设计及播量见表1。

表1 红豆草和无芒雀麦混播草地的播量、混播方式和混播比例Table 1 Mixed patterns, sowing ratios and sowing quantity of O. viciaefolia+B. inermis mixtures
1.3 测定项目与方法
1.3.1出苗及返青率测定 于2015年7月25日进行播种,播深为2~3 cm,播种后及时浇水,并每隔15 d观察出苗情况,直至当年全部停止生长。于翌年返青期进行返青率的测定。
出苗率=单位面积实际出苗数/单位面积实际播种种子数×100%
返青率=单位面积实际返青苗数/单位面积实际出苗数×100%
1.3.2根系形态参数的测定 在无芒雀麦和红豆草种子成熟后,将花盆中的植株和土壤全部取出,用筛孔直径为0.25 mm的网兜将根系部位包裹住,自来水冲洗干净,然后用吸水纸吸干水分,将混播处理的无芒雀麦和红豆草根系分开,用吸水纸平整包裹每种牧草根系,带回实验室进行扫描。在扫描前用细毛刷仔细梳理根系,使其分布均匀,必要时放入水中进行梳理;把每株牧草根系放入扫描根盘中,经甲基蓝染色,均匀平铺于透明胶片上;扫描结束后,根系形态以图形文件格式保存,再利用万深LA-S植物根系分析系统(杭州万深检测科技有限公司)对之分析,获得单株总根长(root length,RL)、根表面积(root surface area,RSA)、根体积(root volume, RV)、根系直径(root diameter, RD)等数据。其中根系直径按照≤0.16 mm,0.16~0.50 mm和≥0.50 mm划分为3个等级,并分别测定其根系长度[39]。
每株牧草根系扫描完成后,将地上部和地下部分开,80 ℃烘干至恒重称量,得到地上和根系生物量(root biomass,RB)。利用根系总根长与根系生物量计算比根长(ratio of root length to root biomass,RLB),即RLB=RL/RB。利用根表面积与根系生物量计算比根面积(ratio of root surface area to root biomass,RSAB),即RSAB=RSA/RB。
1.3.3根系拓扑结构的测定 拓扑指数:Bouma等[11]和Fitter[18]提出了根系拓扑结构的两种极端类型:鱼尾形分支(人字形分支)和叉状分支模式(图1),并且通过拓扑指数来反映不同植物根系的分支模式。其中,拓扑指数TI=lgA/lgM(式中:M为根系所有外部连接的总数,分支与分生组织之间为外部连接;A为最长根系通道内部连接的总数,两个分支点之间为内部连接)。典型的鱼尾形分支TI=1,叉状分支TI接近0.5;拓扑指数越接近1,牧草根系越接近鱼尾形分支;相反,拓扑指数越接近0.5,牧草根系越接近叉状分支[5]。有研究表明,植物根系的分支介于鱼尾形分枝与叉状分支之间,完全的鱼尾形分支和叉状分支在实际研究中很难发现[40]。
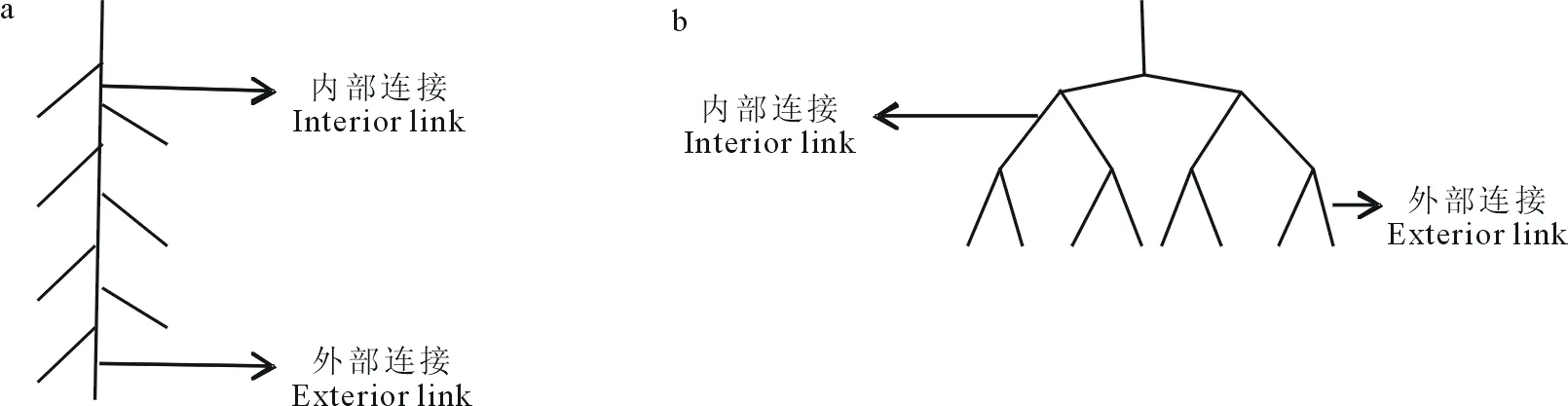
图1 根系拓扑结构示意Fig.1 Schematic diagram of topology of root system a:鱼尾形分支 Herringbone branching; b:叉状分支 Dichotomous branching.
针对Fitter[18]拓扑模型中出现的真空区域,Oppelt等[41]在Fitter拓扑模型基础上提出了新的修正拓扑参数,来说明根系分支状况的中间过渡形式:把从基部到根终端连接数量称为拓扑长度a,最大拓扑长度和Fitter模型的等级A相同,b为根系平均拓扑长度,并且b和根系基部到根终端通道的所有连接总数Pe相关联,可用下式表示:b=Pe/v0(式中:v0相当于Fitter拓扑模型的外部连接总和M),由于a、b随着根系的v0值的改变而变化,所以通过对a、b进行线性转换得到修正值qa、qb,其范围为0~1,方程如下:
式中:lbv0=lnv0/ln 2,其中,鱼尾形分支修正值qa=qb=1;叉状分支qa=qb=0。
1.3.4根系分形维数的测定 采用盒维数法计算根系分形维数[13]:在绘制的根系分布图上绘制单位边长r的正方形,计算根系所截的正方形数目Nr,随着正方形边长逐渐减小,根系所截Nr逐渐增大,得到不同边长水平上相应Nr值后,分别以lgr、lgNr为横坐标和纵坐标作图,得到回归直线方程:lgNr=-FDlgr+lgK。回归直线斜率的负数就是所求的分形维数FD(fractal dimension)。
1.4 数据处理
采用Excel 2010、SPSS 20.0对数据进行统计分析,用Origin 8.0制图。采用q检验法(SNK)和最小差异显著法(LSD)进行不同混播处理间地上生物量、根系生物量、根长、根表面积、根体积、比根长、比根面积和根冠比的差异显著性检验(P<0.05),采用最小二乘法进行地上、地下生物量与根长和根表面积之间的线性回归分析。
2 结果与分析
2.1 出苗及返青情况
由表2可知,同行混播+15 cm行距(T15)和异行混播+15 cm行距(Y15)无芒雀麦和红豆草具有较高的出苗率,显著高于其他混播处理(P<0.05)。从2016年3月20日开始,每隔15 d观察返青情况,直至2016年4月15日(返青情况见表2)。同行混播+15 cm行距+根系阻隔(YZ15)和异行混播+30 cm行距(Y30)的无芒雀麦具有较高的返青率,显著高于单播和T15;单播红豆草具有较高的返青率,显著高于T15和Y30。但各混播处理间基本苗数无显著差异(P>0.05)。

表2 红豆草和无芒雀麦混播草地出苗及返青情况(均值±标准差)Table 2 Seedling emergence and green emergence of O. viciaefolia+B. inermis mixtures (%)
注:同列不同小写字母表示差异显著(P<0.05),下同。
Note:Values designated by different lowercase letters were significantly different in the same column (P<0.05), the same below.
2.2 不同混播方式下地上生物量、根系生物量和根冠比的比较
种群空间结构对无芒雀麦根系生物量、地上生物量影响明显(图2):异行混播处理较单播和同行混播处理根系生物量、地上生物量均有显著增加(P<0.05);行距越大根系生物量、地上生物量值越高;异行+30 cm行距混播处理(Y30)根系生物量、地上生物量比同行+15 cm行距(T15)处理增加了35.15%和45.38%,比单播处理(CK2)增加了22.59%和35.29%,且差异显著(P<0.05);异行+15 cm行距(Y15)根系生物量、地上生物量也显著高于CK2(P<0.05);但T15、异行+15 cm行距+根系阻隔(YZ15)与CK2相比,根系生物量、地上生物量没有显著变化。T15、Y30根冠比显著高于CK2(P<0.05),且Y30根冠比显著高于其他处理(P<0.05);T15、Y15和YZ15之间根冠比无显著差异(P>0.05)。
种群空间结构对红豆草地上生物量影响明显,而对根系生物量和根冠比影响较小:异行混播处理较T15、CK1均有显著增加(P<0.05);行距越大地上生物量值越高;Y30地上生物量比T15增加了24.55%,比CK1增加了15.38%,且差异显著(P<0.05)。T15根系生物量显著高于其他处理(P<0.05),其他处理间无显著差异(P>0.05);Y15根冠比显著小于其他处理(P<0.05),其他处理间无显著差异(P>0.05)。
2.3 不同混播方式下不同直径范围内根系长度的分布
如表3所列,无芒雀麦和红豆草在不同混播处理下不同直径范围的根系长度分布发生了显著变化:与T15相比,Y15、Y30处理无芒雀麦≤0.16 mm的细根和≥0.50 mm的粗根显著增多(P<0.05),而Y15处理0.16~0.50 mm的根系明显减少(P<0.05);Y30处理无芒雀麦3个直径范围内的根系均较多。Y15处理红豆草≤0.16 mm的细根比CK1显著增多(P<0.05),≥0.50 mm的粗根则显著少于Y30;Y15和YZ15处理0.16~0.50 mm的根系显著多于其他处理(P<0.05)。这表明异行混播处理下无芒雀麦≤0.16 mm的细根显著增加,而红豆草0.16~0.50 mm的根系显著增加。

图2 不同混播方式对牧草地上生物量、根系生物量和根冠比的影响Fig.2 Effects of different mixed patterns on aboveground biomass, root biomass and root-shoot ratio of B. inermis and O. viciaefolia 不同小写字母(a, b, c, d)表示无芒雀麦不同处理间差异显著(P<0.05),不同小写字母(a′, b′, c′, d′)表示红豆草不同处理间差异显著(P<0.05),下同。Lowercase letters (a, b, c, d) indicate significant differences between different processing tags of B. inermis (P<0.05), other lowercase letters(a′, b′, c′, d′) indicate significant differences between different processing tags of O. viciaefolia (P<0.05), the same below.

牧草种类Foragespecies处理Treatment直径范围Diameterrange≤0.16mm0.16~0.50mm≥0.50mm无芒雀麦B.inermisT159.67±1.23b22.62±6.23a10.11±0.95bY1515.78±2.26a11.14±2.03b24.48±4.15aYZ1514.59±3.12ab23.82±5.69a21.53±5.36aY3019.22±2.85a25.23±4.16a20.01±1.12aCK215.24±1.56a26.53±2.20a15.37±1.04ab红豆草O.viciaefoliaT1518.04±4.89ab23.17±6.86c14.95±2.11abY1521.29±5.12a41.93±9.86a10.23±1.36bYZ1516.37±4.49ab52.35±9.89a14.59±2.03abY3017.48±4.15ab34.94±8.10b19.78±2.89aCK115.07±2.05b14.01±2.01d12.17±1.49b
2.4 不同混播方式下根系形态参数的比较
如图3所示,无芒雀麦在不同混播处理下根系长度(RL)、根表面积(RSA)、根体积(RV)、比根长(RLB)和比根面积(RSAB)发生了显著变化:Y30处理的RL、RLB和RSA较高,与T15均有显著差异(P<0.05),Y30处理的RL比YZ15和T15处理分别增加了30%和40%;YZ15处理的RSA、RV和RSAB较高,均显著高于T15处理(P<0.05),YZ15处理的RSA比Y15和T15处理分别增加了39.97%和67.74%;Y30处理具有较小的RV和RSAB,显著低于YZ15处理(P<0.05);由同行混播转变为异行混播,RL、RSA均呈增加趋势。红豆草在不同混播处理下RL、RV、RLB和RSAB发生了显著变化:由同行混播转变为异行混播RL呈增加趋势,YZ15处理显著高于T15和CK1处理(P<0.05);Y30、YZ15和T15处理的RV显著高于Y15和CK1处理(P<0.05);T15处理具有较高的RLB值,且显著高于Y15和YZ15处理(P<0.05);YZ15处理具有较高的RSAB值,且显著高于Y15和T15处理(P<0.05)。红豆草在不同混播处理下RSA未发生显著变化(P>0.05)。由此可见,异行混播+行距增大使得2种牧草的细根比例增加,增加了RL、RSA和RLB;异行混播+阻隔使得2种牧草的根系直径增加,从而使得RV和RSAB增加。

图3 不同混播方式对牧草根系形态参数的影响Fig.3 Effects of different mixed patterns on root morphological parameters of B. inermis and O. viciaefolia
2.5 种群空间距离(行距)与根系形态参数变化的关系
将同行混播(T15)设定为豆禾牧草种群空间距离为0 cm,Y15设定为豆禾牧草种群空间距离为15 cm,Y30设定为豆禾牧草种群空间距离为30 cm,YZ15由于根系阻隔,假定为豆禾牧草种群空间距离为45 cm。由图4可知(n=20),2种牧草的RL均呈多项式变化曲线,即随着种群空间距离增加,RL先增加,在Y30(行距30 cm)处到达高峰,然后下降。2种牧草的RSA也均呈多项式变化曲线,但无芒雀麦的RSA在Y15(行距15 cm)处到达最小值,以后随着种群空间距离增加而增加;红豆草的RSA随着种群空间距离增加而增加,在YZ15(行距45 cm)处到达最高值。另外,从图4还可知,无芒雀麦的RL和RSA均高于红豆草。上述结果表明,随着种群空间距离增加,2种牧草的根系形态发生显著变化,朝着根系更长、表面积更大的方向改变。
2.6 不同混播方式下根系拓扑指数与分形维数的比较
由表4可以看出,2种牧草不同混播方式下拓扑指数(TI)均接近于1,修正拓扑指数(qa)大于0.5,表明2种牧草根系分支结构均接近鱼尾状分支模式(图1a),根系主要通过快速生长延伸,扩大其有效营养空间,从而有效地从土壤中获取资源。独立样本t检验表明,两种牧草根系TI无显著差异(F=0.05,P=0.83>0.05)。无芒雀麦分形维数(FD)均值为1.5534,大于红豆草FD均值(1.4528);独立样本t检验表明,2种牧草分形维数无显著差异(F=1.18,P=0.34>0.05)。说明在相同生境条件下,无芒雀麦和红豆草根系分支模式表现出趋同性。
无芒雀麦所有混播处理TI显著大于CK无芒雀麦处理(P<0.05);YZ15处理具有较高的基部到终端连接总和(Pe)、外部连接总和(vo)和TI值,其中Pe和vo值显著高于其他处理(P<0.05),其根系构型具有典型的鱼尾状分支特点;Y30和T15处理具有较小的qa值,二者根系构型有向叉状分支过渡的趋势(图5);YZ15处理具有较高的FD值,显著大于CK2和T15处理(P<0.05)。红豆草YZ15和Y30处理具有较高的Pe和vo值,但qa值较低,TI值小于1,二者根系构型有向叉状分支过渡的趋势;T15处理和CK1具有较高的qa值,其中CK1TI值接近1,其根系构型具有典型的鱼尾状分支特点(图5);YZ15处理也具有较高的FD值,显著大于T15和Y15处理(P<0.05)。

图4 种群空间距离(行距)与根系形态参数变化的关系Fig.4 The relationship between the space length and root morphology YBL表示无芒雀麦根长随种群空间距离(行距)变化曲线方程,YOL表示红豆草根长随种群空间距离(行距)变化曲线方程,YBA表示无芒雀麦根表面积随种群空间距离(行距)变化曲线方程,YOA表示红豆草根表面积随种群空间距离(行距)变化曲线方程。YBL is stand for the curve equation of root length changes with population space distance about B. inermis; YOL is stand for the curve equation of root length changes with population space distance about O. viciaefolia; YBA is stand for the curve equation of surface area changes with population space distance about B. inermis; YOA is stand for the curve equation of root surface area changes with population space distance about O. viciaefolia.
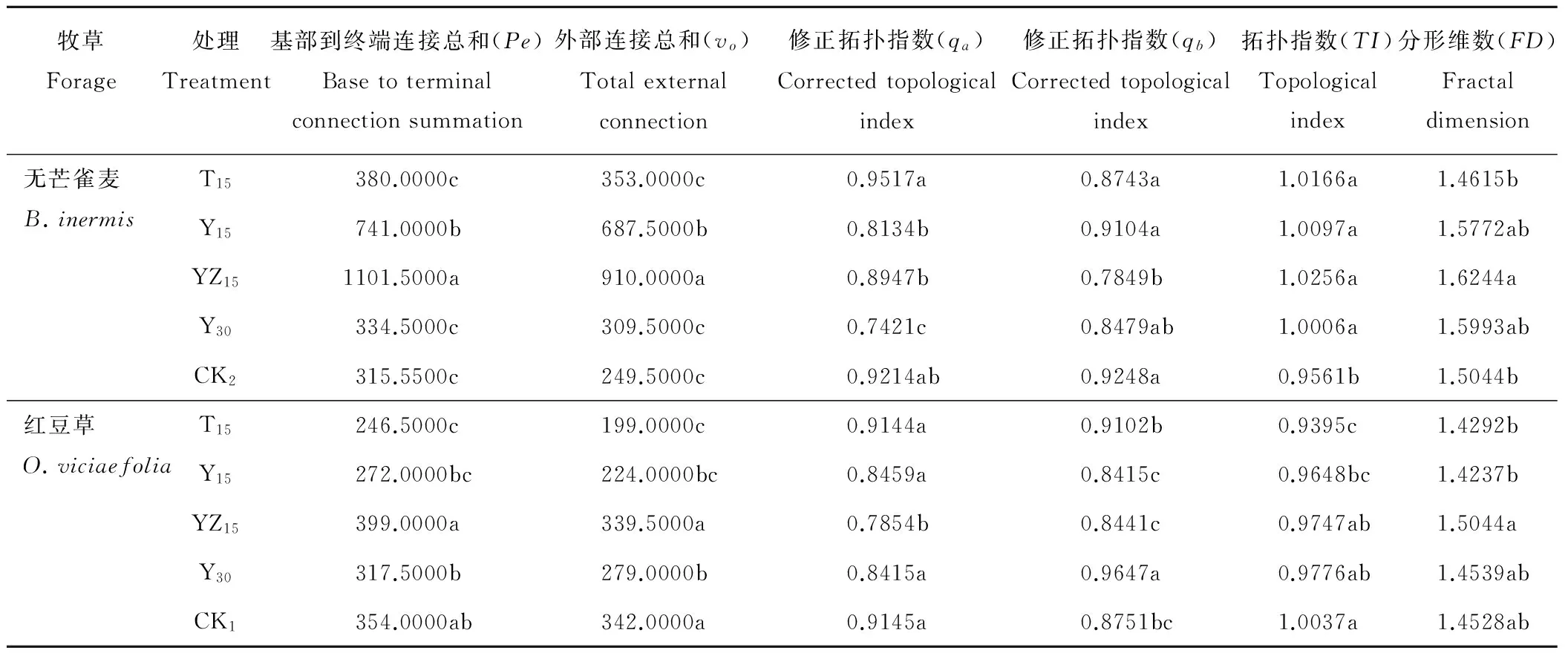
牧草Forage处理Treatment基部到终端连接总和(Pe)Basetoterminalconnectionsummation外部连接总和(vo)Totalexternalconnection修正拓扑指数(qa)Correctedtopologicalindex修正拓扑指数(qb)Correctedtopologicalindex拓扑指数(TI)Topologicalindex分形维数(FD)Fractaldimension无芒雀麦B.inermisT15380.0000c353.0000c0.9517a0.8743a1.0166a1.4615bY15741.0000b687.5000b0.8134b0.9104a1.0097a1.5772abYZ151101.5000a910.0000a0.8947b0.7849b1.0256a1.6244aY30334.5000c309.5000c0.7421c0.8479ab1.0006a1.5993abCK2315.5500c249.5000c0.9214ab0.9248a0.9561b1.5044b红豆草O.viciaefoliaT15246.5000c199.0000c0.9144a0.9102b0.9395c1.4292bY15272.0000bc224.0000bc0.8459a0.8415c0.9648bc1.4237bYZ15399.0000a339.5000a0.7854b0.8441c0.9747ab1.5044aY30317.5000b279.0000b0.8415a0.9647a0.9776ab1.4539abCK1354.0000ab342.0000a0.9145a0.8751bc1.0037a1.4528ab
2.7 生物产量、根冠比与根系形态参数之间的相互关系
对无芒雀麦根系生物量、地上生物量和根冠比与根系形态参数之间进行拟合发现(n=50,表5):RL、RSA与根系生物量、地上生物量呈线性的正相关关系(P<0.01),与根冠比相关性不显著(P>0.05);RV与根系生物量、地上生物量和根冠比相关性均不显著(P>0.05);根系平均直径(RD)与根系生物量呈线性的正相关关系(P<0.05),与地上生物量相关性不显著(P>0.05),与根冠比也呈线性的正相关关系(P<0.01);RLB和RSAB与根系生物量呈非线性相关(幂函数,P<0.05),与地上生物量呈线性的负相关关系(P<0.05),与根冠比相关性不显著(P>0.05)。对红豆草根系生物量、地上生物量和根冠比与根系形态参数之间进行拟合发现(表6):RL、RSA与根系生物量、地上生物量呈线性的正相关关系(P<0.01),与根冠比呈非线性相关(幂函数,P<0.05);RV、RD与根系生物量、地上生物量和根冠比相关性均不显著(P>0.05);RLB与根系生物量呈非线性相关(二次多项式函数,P<0.05),与地上生物量、根冠比相关性均不显著(P>0.05);RSAB与根系生物量呈非线性相关(对数函数,P<0.05),与地上生物量、根冠比相关性均不显著(P>0.05)。上述结果表明,根系形态参数与根系生物量关联性较强,而与地上生物量、根冠比关联性较差。
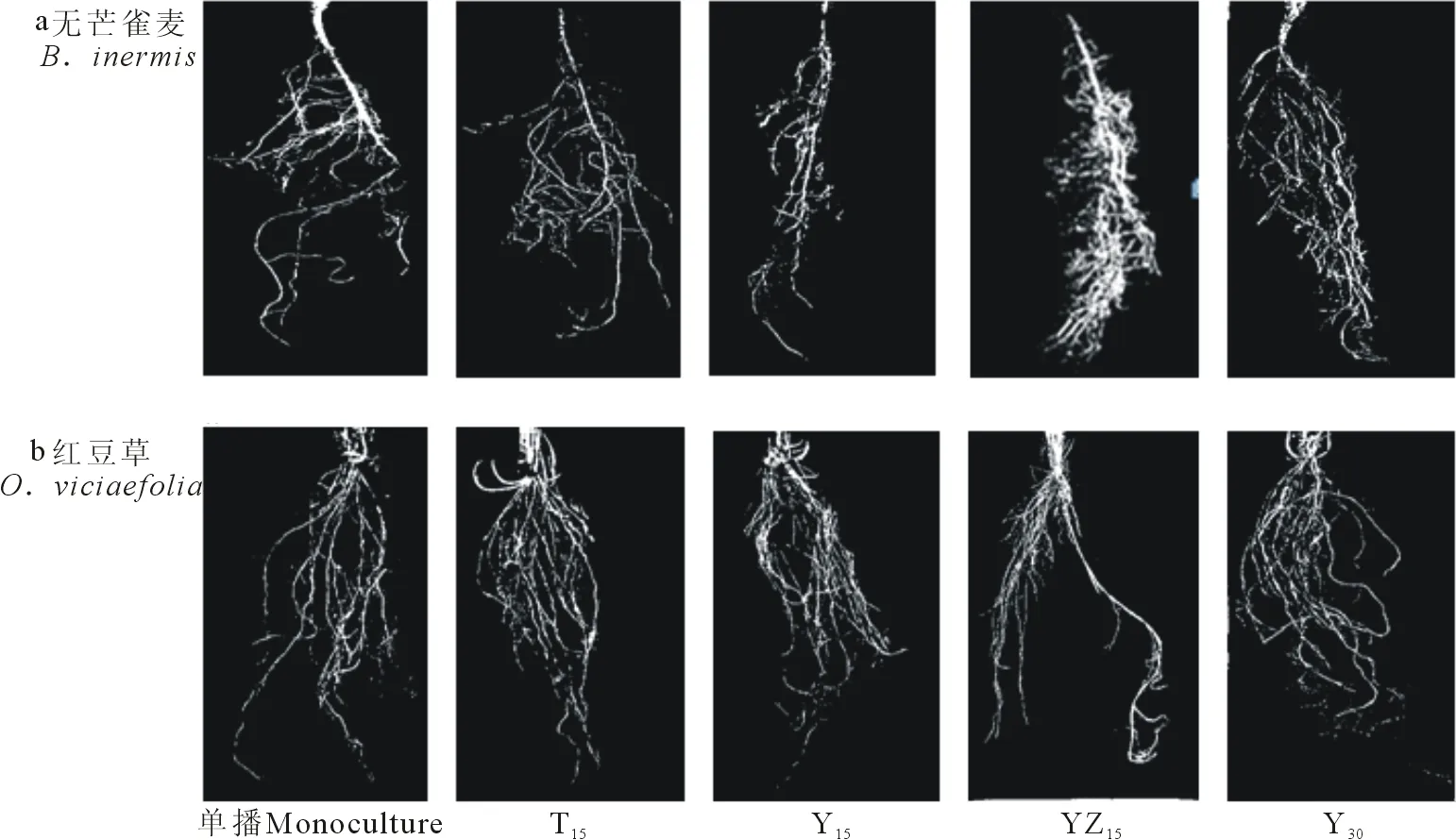
图5 不同混播方式下2种牧草根系拓扑结构Fig.5 Root topology structures of B. inermis and O. viciaefolia under different mixed patterns
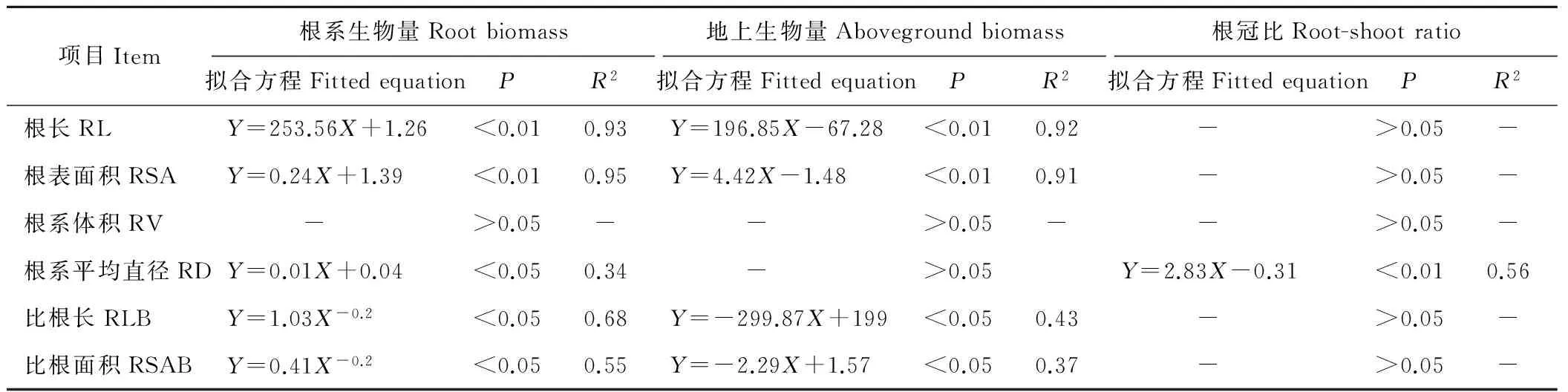
项目Item根系生物量Rootbiomass拟合方程FittedequationPR2地上生物量Abovegroundbiomass拟合方程FittedequationPR2根冠比Root-shootratio拟合方程FittedequationPR2根长RLY=253.56X+1.26<0.010.93Y=196.85X-67.28<0.010.92 ->0.05-根表面积RSAY=0.24X+1.39<0.010.95Y=4.42X-1.48<0.010.91 ->0.05-根系体积RV ->0.05- ->0.05- ->0.05-根系平均直径RDY=0.01X+0.04<0.050.34 ->0.05Y=2.83X-0.31<0.010.56比根长RLBY=1.03X-0.2<0.050.68Y=-299.87X+199<0.050.43 ->0.05-比根面积RSABY=0.41X-0.2<0.050.55Y=-2.29X+1.57<0.050.37 ->0.05-

表6 红豆草地上、地下生物量及根冠与根系形态参数之间的拟合方程Table 6 The fitted equations between aboveground biomass, root biomass, root-shoot ratio and root morphological parameters of O. viciaefolia
3 讨论
3.1 不同混播方式对根系形态的影响
群体空间配置的变化(间套作,同行/异行)直接改变了植物群体光能的截获、地上地下种间竞争过程以及产量潜力的发挥[43]。而根系形态决定了植株获得水分和养分的能力,影响地下种间竞争过程和产量的形成[44]。本研究表明,随着采用异行混播和行距的增大,混播牧草间相互竞争强度下降,无芒雀麦和红豆草地上生物量均增加,无芒雀麦根系生物量也增加,但红豆草根系生物量变化不明显(图2)。这可能是由于无芒雀麦在混播群体中属于高位植物,而红豆草属于低位植物[14];红豆草为了获取更多光资源,光合产物优先供应地上光合器官的生长,影响了地下根系的生长。与此同时,由于地上种间竞争强度的减弱,牧草地上获取光资源环境的改善,使得地下根系生长获得更多资源分配,无芒雀麦和红豆草的根冠比在异行混播+30 cm行距处理下均明显增加。这种地下根系生长获得更多资源分配在根系形态上的表现为:异行混播处理下无芒雀麦≤0.16 mm的细根和红豆草0.16~0.50 mm的根系增加(表3);异行混播+行距增大(Y30)使得2种牧草的RL、RSA和RSAB增加;异行混播+阻隔(YZ15)使得2种牧草的RV和RSAB增加(图3)。已有研究表明,植物根系形态对竞争/胁迫生境的适应往往是通过根系空间分布格局[13]、分支角度(基根角度)[29]、根长和根系表面积[45]等特征的改变表现出来,是植物根系构建的资源配置模式对异质性生境适应的结果[13]。本研究中,2种牧草的RL、RSA与根系生物量和地上生物量均呈线性的正相关关系(表5,6)。由此可见,混播群体中牧草根系形态对种间竞争的适应,是通过牧草根系可塑性变化导致的资源配置模式变化来实现的。
关于种群相互作用对根系生长和形态的影响有不同解释,有研究表明这种相互作用对根系生长的影响是养分竞争的结果[17],也有研究表明影响根系生长的是不同植物个体的生理和基因识别的结果[46]。本研究中,种群空间距离增加,2种牧草的RL、RSA均呈增加趋势,但异行混播+阻隔(YZ15)处理下红豆草RL、RSA具有较高值;大部分处理无芒雀麦的RL、RSA、RV、RLB和RSAB均高于红豆草(图2,4)。这表明牧草的根系形态是由遗传因素控制的,但种间相互作用通过权衡地上、地下生长,改变了其表型特征。
3.2 不同混播方式对牧草根系拓扑结构与分形特征的影响
分析根系拓扑结构主要是找出生境条件的改变是否会使根系构型构建策略(如生长、分叉、变细/变粗)发生适应性变化[5]。根系的拓扑结构直接影响到植物根系吸收效率和根系构建过程中所需的碳量[13],可以用根系生长资源投入(如碳)与土壤水分养分收益的权衡关系来诠释[41]。已有研究认为:鱼尾状分支结构根系中,次级根分支较少,重叠较少,内部竞争较小;叉状分支的根系由于次级分支较多,重叠增加,内部竞争较强,然而叉状分支可以加强根系在土层中的空间分布,有利于提高植物在资源贫瘠生境中竞争土壤资源的能力[11,18]。宋清华等[13]指出,在地上和地下生存空间拥挤、资源贫瘠的环境中,为了避免不必要的种内竞争,牧草往往限制了地下生物量的分配比例,增加根系连接长度、减少分叉数来权衡资源投入与土壤水分养分收益的关系。本研究表明,2种牧草根系拓扑指数(TI)接近于1,且修正拓扑指数qa≈修正拓扑指数qb≈1,根系分支模式均接近鱼尾形分支;但随着种群空间距离的增加(Y30、YZ15),2种牧草根系的拓扑结构有向叉状分支方向发展的趋势(表4,图5)。在豆禾牧草混播竞争与共存的生境中:种间竞争强的生境中(T15),无芒雀麦和红豆草通过减少根长、根系表面积和根系连接长度(Pe,v0),增加根系中较粗根系比例来权衡根系构型与土壤水分养分吸收的关系;随着种间竞争强度(Y30)的下降,无芒雀麦和红豆草不仅根长、根系表面积和根系连接长度增加,而且分支能力也在增加,增加了地下生物量的分配比例,使得混播豆禾牧草养分水分高效利用具有更加坚实的基础。
分形维数不仅可表征植物根系发育程度,而且还能精确反映在各种胁迫下根系结构的变化和适应策略。单位碳的投入下,根系分支越简单,空间占有能力越强;根系分支越复杂,根系重叠加剧,空间占有能力越弱[42]。因此,水分或养分贫瘠条件下根系分支简单,分形维数较小;反之,根系分支复杂,分形维数较大[32]。本研究中,相同生境条件下,无芒雀麦分形维数大于红豆草,且随着种群空间距离的增加(Y30、YZ15),2种牧草的分形维数也在增加。这表明强种间竞争使得根系分支简单,分形维数变小,而降低种间竞争强度有利于混播牧草根系发展更为复杂的根系结构,增加空间占有能力,进一步提高养分水分利用效率。
4 结论
种群的空间距离(行距)、种群的空间作用方式(同行/异行/异行阻隔)改变了无芒雀麦+红豆草混播草地地下根系的竞争环境,使得2种混播牧草的根系形态、拓扑结构和分形特征发生了改变。从同行混播到异行混播以及行距增加使得地上种间竞争强度的减弱,牧草地上获取光资源环境改善,2种牧草地上生物量增加;通过权衡资源分配,使得地下根系生长获得更多的资源分配,根系形态发生了改变:无芒雀麦的根系生物量和根冠比增加,2种牧草根系更长,细根比例更多,根系表面积也增大,空间拓展能力增强。2种牧草在不同混播方式下根系分支结构均为鱼尾状分支;随着种群空间距离的增加,2种牧草的根系拓扑结构发生了优化(由鱼尾状分支向叉状分支过渡)和增加分形维数,进一步增加了空间占有能力,使得混播牧草水分、养分利用效率提高。这种结果较好地反映了牧草根系通过形态、拓扑结构和分形的可塑性来响应种间竞争关系变化的协同适应,是混播群体高产和高效利用水分养分的重要基础。
References:
[1] Kong X P, Zhang M L, de Smet I,etal. Designer crops: Optimal root system architecture for nutrient acquisition. Trends in Biotechnology, 2014, 32: 597-598.
[2] Plaza-Bonilla D,lvaro-Fuentes J, Hansen N C,etal. Winter cereal root growth and aboveground-belowground biomass ratios as affected by site and tillage system in dryland Mediterraneanconditions. Plant and Soil, 2014, 374: 925-939.
[3] Wen W L, Guo X Y, Zhao C J,etal. Crop roots configuration and visualization: A review. Scientia Agricultura Sinica, 2015, 48(3): 436-448.
温维亮, 郭新宇, 赵春江, 等. 作物根系构型三维探测与重建方法研究进展. 中国农业科学, 2015, 48(3): 436-448.
[4] Song Q H, Zhao C Z, Shi Y C,etal. Fractal root system ofMelicaprzewalskyialong different aspect in degraded grassland. Chinese Journal of Plant Ecology, 2015, 39(8): 816-824.
宋清华, 赵成章, 史元春, 等. 高寒草地甘肃臭草根系分形结构的坡向差异性. 植物生态学报, 2015, 39(8): 816-824.
[5] Guo J H, Zeng F J, Li C J,etal. Root architecture and ecological adaptation strategies in three shelterbelt plant species in the southern Taklimakan Desert. Chinese Journal of Plant Ecology, 2014, 38(1): 36-44.
郭京衡, 曾凡江, 李尝君, 等. 塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略. 植物生态学报, 2014, 38(1): 36-44.
[6] Eshel A. On the fractal dimensions of a root system. Plant, Cell and Environment, 1998, 21: 247-251.
[7] Walk T C, van Erp E, Lynch J P. Modelling applicability of fractal analysis to efficiency of soil exploration by roots. Annals of Botany, 2004, 94: 119-128.
[8] Malamy J E. Intrinsic and environmental response pathways that regulate root system architecture. Plant, Cell & Environment, 2005, 28: 67-77.
[9] Guswa A J. Effect of plant uptake strategy on the water optimal root depth. Water Resources Research, 2010, 46: W09601, doi: 10.1029/2010WR009122.
[10] Chaila S, Diaz L, Sobrero M T,etal. Competencia deSicyospolyacanthusen caa de azúcar interference ofSicyospolyacanthusin sugarcane. Planta Daninha, 2004, 22(4): 545-551.
[11] Bouma T J, Nielsen K L, Vanhal J,etal. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Functional Ecology, 2001, 15: 360-369.
[12] Oppelt A L, Kurth W, Godbold D L. Topology, scaling relations and Leonardo’s rule in root systems from African tree species. Tree Physiology, 2001, 21: 117-128.
[13] Song Q H, Zhao C Z, Shi Y C,etal. Trade-off between root forks and link length ofMelicaprzewalskyion different aspects of slopes. Chinese Journal of Plant Ecology, 2015, 39(6): 577-585.
宋清华, 赵成章, 史元春, 等. 不同坡向甘肃臭草根系分叉数和连接长度的权衡关系. 植物生态学报, 2015, 39(6): 577-585.
[14] Zheng W, Zhu J Z, Kuerban,etal. Dynamics of interspecific competition of legume-grass mixture under different mixed sowing patterns. Acta Agresia Sinica, 2010, 18(4): 568-575.
郑伟, 朱进忠, 库尔班, 等. 不同混播方式下豆禾混播草地种间竞争动态研究. 草地学报, 2010, 18(4): 568-575.
[15] Schenk H J. Root competition: beyond resource depletion. Journal of Ecology, 2006, 94(4): 725-739.
[16] Craine J M. Competition for nutrients and optimal root allocation. Plant and Soil, 2006, 285(1/2): 171-185.
[17] Fransen B, Blijjenberg J, de Kroon H. Root morphological and physiological plasticity of perennial grass species and the exploitation of spatial and temporal heterogeneous nutrient patches. Plant and Soil, 1999, 211(2): 179-189.
[18] Fitter A H. Architecture and biomass allocation as components of the plastic response of root system to soil heterogeneity//Caldwell M M, Peary R W. Exploitation of Environmental Heterogeneity by Plants: Ecophysiological Process Above and Belowground. San Diego: Academic Press, 1994: 305-324.
[19] Hao Y R, Lao X R, Zhao B Q,etal. Effect of separating root method on wheat and corn intercropping system. Journal of Triticeae Crops, 2003, 23(1): 71-74.
郝艳如, 劳秀荣, 赵秉强, 等. 隔根对小麦/玉米间套种植生长特性的影响. 麦类作物学报, 2003, 23(1): 71-74.
[20] Song R, Mu Y, Wang Y L,etal. Effects of intercropping of maize and soybean on the morphological character of roots. Journal of Northeast Normal University (Natural Science Edition), 2002, 34(3): 83-86.
宋日, 牟瑛, 王玉兰, 等. 玉米、大豆间作对两种作物根系形态特征的影响.东北师范大学学报(自然科学版), 2002, 34(3): 83-86.
[21] Li S M. Mechanism of Interspecific Facilitation on Phosphorus Uptake by Crops in Intercropping System. Beijing: China Agriculture University, 2004.
李淑敏. 间作作物吸收磷的种间促进作用机制研究. 北京: 中国农业大学, 2004.
[22] Ma Z H, Che R J, Wang H Y,etal. Effect of different seeding rates and planting patterns on root morphological traits and root vigor of super-high-yield soybean cultivars. Scientia Agricultura Sinica, 2015, 48(6): 1084-1094.
马兆惠, 车仁君, 王海英, 等. 种植密度和种植方式对超高产大豆根系形态和活力的影响.中国农业科学, 2015, 48(6): 1084-1094.
[23] Eekerena N V, Bosa M, Wita Jan de,etal. Effect of individual grass species and grass species mixtures on soil quality as related to root biomass and grass yield. Applied Soil Ecology, 2010, 45(3): 275-283.
[24] Li L, Zhang F S, Li X L,etal. Interspecific facilitation of nutrient uptake by intercropped maize and faba bean. Nutrient Cycling in Agroecosystems, 2003, 65(1): 61-71.
[25] Li L, Li S M, Zhou L L,etal. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus deficient soils. PNAS, 2007, 104(27): 11192-11196.
[26] Li W X. Nitrate Accumulation in Soil and Nutrient Acquisition by Intercropped Wheat, Maize and Faba Bean. Beijing: China Agriculture University, 2001.
李文学. 小麦/玉米/蚕豆间作系统中氮、磷吸收利用及其环境效应. 北京: 中国农业大学, 2001.
[27] Zhang Y K, Chen F J, Li L,etal. The role of maize root size in phosphorus uptake and productivity of maize/faba bean and maize/wheat intercropping systems.Scientia Sinica (Life Science), 2012, 42(10): 841-849.
张义凯, 陈范骏, 李隆, 等. 不同类型的玉米根系对间作体系磷高效吸收以及生产力的影响. 中国科学: 生命科学, 2012, 42(10): 841-849.
[28] Li Y Y, Hu H S, Cheng X,etal. Effects of interspecific interactions and nitrogen fertilization rates on above-and below-growth in faba bean/mazie intercropping system. Acta Ecologica Sinica, 2011, 31(6): 1617-1630.
李玉英, 胡汉升, 程序, 等. 种间互作和施氮对蚕豆/玉米间作生态系统地上部和地下部生长的影响. 生态学报, 2011, 31(6): 1617-1630.
[29] Cui H, Li L Y, Xie X L,etal. Differences in root architecture of severalStylosanthesgenotypes and their phosphorus efficiency. Acta Prataculturae Sinica, 2013, 22(5): 265-271.
崔航, 李立颖, 谢小林, 等. 不同基因型柱花草的根系构型差异及其磷效率. 草业学报, 2013, 22(5): 265-271.
[30] Zhang E H, Huang G B. Temporal and spatial distribution characteristics of the crop root in intercropping system. Chinese Journal of Applied Ecology, 2003, 14(8): 1301-1304.
张恩和, 黄高宝. 间套种植复合群体根系时空分布特征. 应用生态学报, 2003, 14(8): 1301-1304.
[31] Gan Y W, Li L, Li L H,etal. Study of root distribution of walnut/wheat intercropping system in Southern Xinjiang. Acta Agriculturae Boreali-occidentalis Sinica, 2015, 24(3): 102-110.
甘雅文, 李隆, 李鲁华, 等. 南疆核桃间作冬麦复合系统根系分布特征研究. 西北农业学报, 2015, 24(3): 102-110.
[32] Yang X L, Zhang X M, Li Y L,etal. Analysis and estimation of root architecture in the hinterland of Taklimakan desert base on the fractal theory. Journal of Arid Land Resources and Environment, 2015, 29(8): 145-160.
杨小林, 张希明, 李义玲, 等. 基于分形理论的塔克拉玛干沙漠腹地自然植物根系构型特征分析. 干旱区资源与环境, 2015, 29(8): 145-160.
[33] Zhou B, Yan X H, Xiao Y A,etal. Module biomass ofAgeratumconyzoidespopulations in different habitats.Acta Ecologica Sinica, 2015, 35(8): 1-8.
周兵, 闫小红, 肖宜安, 等. 不同生境下入侵植物胜红蓟种群构件生物量分配特性研究. 生态学报, 2015, 35(8): 1-8.
[34] Zhou Y S, Wang L Q. Ecological adaptation of root architecture to grassland degradation inPotentillaacaulis. Chinese Journal of Plant Ecology, 2011, 35(5): 490-499.
周艳松, 王立群. 星毛委陵菜根系构型对草原退化的生态适应. 植物生态学报, 2011, 35(5): 490-499.
[35] Cop J, Korosec J. Composition and herbage yield of grass/while clover mixtures in relation to nitrogen fertilization under cutting//Mannetie L, Frame J. Grassland and Society Proceeding of the 15th General Meeting of the European Grassland Federation June 6-9. 1994: 77-80.
[36] Zheng W, Zhu J Z, Jianaerguli. A comprehensive evaluation of the productive performance of legume-grass mixture under different mixed sowing patterns. Acta Prataculturae Sinica, 2012, 21(6): 242-251.
郑伟, 朱进忠, 加娜尔古丽. 不同混播方式豆禾混播草地生产性能的综合评价. 草业学报, 2012, 21(6): 242-251.
[37] Zheng W, Zhu J Z, Jianaerguli,etal. Effects of different mixed sowing patterns on production performance of legume-grass mixture.Chinese Journal of Grassland, 2011, 33(5): 45-52.
郑伟, 朱进忠, 加娜尔古丽, 等. 不同混播方式对豆禾混播草地生产性能的影响. 中国草地学报, 2011, 33(5): 45-52.
[38] Zhang Q Q, Jin G L, Zhu J Z,etal. Analyzing spatial patterns of prime plant population in mix-sowed artificial grassland with different established years. Acta Agrestia Sinica, 2011, 19(5): 735-739.
张强强, 靳瑰丽, 朱进忠, 等. 不同建植年限混播人工草地主要植物种群空间分布格局分析. 草地学报, 2011, 19(5): 735-739.
[39] Wang J Y, Wang H Q, Liang X D,etal. Response of root morphology and N absorption to nitrate nitrogen supply in hydroponic oats. Journal of Plant Nutrition and Fertilizer, 2016, 22(4): 1049-1055.
王俊英, 王华青, 梁晓东, 等. 水培燕麦根系形态和氮吸收流量对硝态氮供应浓度的响应. 植物营养与肥料学报, 2016, 22(4): 1049-1055.
[40] Vercambre G, Pagès L, Doussan C,etal. Architectural analysis and synthesis of the plum tree root system in an orchard using a quantitative modelling approach. Plantand Soil, 2003, 251: 1-11.
[41] Oppelt A L, Kurth W, Jentschke G,etal. Contrasting rooting patterns of some arid-zone fruit tree species from Botswana-II. Coarse root distribution. Agroforestry Systems, 2005, 64: 1-11.
[42] Yang P L, Luo Y P. Fractal characteristics of winter wheat roots. Chinese Science Bulletin, 1994, 39(20): 1911-1933.
杨培岭, 罗远培. 冬小麦根系形态的分形特征. 科学通报, 1994, 39(20): 1911-1933.
[43] Yang F, Huang S, Gao R,etal. Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red: far-red ratio. Field Crops Research, 2014, 155: 245-253.
[44] Costa C, Dwyer L M, Hamilton R I. A sampling method for measurement of large root systems with scanner-based image analysis. Agronomy Journal, 2000, 92(4): 621-627.
[45] Yang F, Lou Y, Liao D P,etal. Effects of row spacing on crop biomass, root morphology and yield in maize-soybean relay strip intercropping system. Acta Agronomica Sinica, 2015, 41(4): 642-650.
杨峰, 娄莹, 廖敦平, 等. 玉米-大豆带状套作行距配置对作物生物量、根系形态及产量的影响. 作物学报, 2015, 41(4): 642-650.
[46] Ljung K, Hull A K, Yamada M,etal. Sites and regulation of auxin biosynthesis inArabidopsisroots. Plant Cell, 2005, 17(4): 1090-1104.
