紫玉兰和二乔玉兰花部形态的变异研究
2018-01-10唐浩君司马永康
唐浩君 司马永康
摘要:为紫玉兰(Magnolia liliflora(Desr)D.L.Fu)和二乔玉兰(Magnolia soul.-Bod)的分类学、形态学和园林植物开发等提供基础数据,特对其花部特征变异进行了研究,通过方差分析得出:两者花部都存在数量和质量上的变异,紫玉兰的变异主要发生在株间,而二乔玉兰的变异则发生在株间和株内。这两者的变异都主要发生在雄蕊和心皮特征上。
关键词:花部;形态特征;变异式样图
紫玉蘭(Magnolia liliiflora(Desr.)D.L.Fu)又名木笔、辛夷,是木兰科(Magnoliaeeae)玉兰属乔木;二乔玉兰(Magnolia soulangeana Soul.-Bod)为玉兰和紫玉兰的杂交种,性状介于二者之间。是木兰科木兰属,原产于我国中部。在现有研究报道中,已见的紫玉兰和二乔玉兰的报道多在其分类学㈣、繁殖与栽培技术、生理与遗传特性、植物化学、孢粉学和胚胎学等方面,尚未见花部形态变异研究。本文研究了紫玉兰和二乔玉兰的花部数量变异及其变异式样,并对比这2个近缘种的关系,旨在为紫玉兰和二乔玉兰的分类学、形态学和园林植物开发等提供基础数据。
1验材料与研究方法
1.1试验材料
2010年2月~2011年4月,在云南农业大学校园内和云南省林业科学研究院昆明树木园内的引种居群中,共采5株紫玉兰作为实验材料,代号Z,每株10朵,共50朵;共采6株二乔玉兰作为试验材料,代号R,每株5朵,共30朵。
1.2研究方法
每朵花观察了如下形状与性状:花的形状、颜色,梗毛被颜色,花被片的形状、颜色、毛被、顶端形状及顶端边缘,雌蕊的形状,花药的颜色,花隔的颜色,雌蕊群的形状和颜色,雌蕊的毛被,花托的颜色和毛被,心皮软硬、厚薄,柱头的颜色等。每朵花测量并记录如下数量性状:花序梗(数、长,径)、花梗(数、长、径)、花被片(总数,总轮数,第1轮数、长、宽,第2轮数、长、宽,第3轮数、长、宽)、雄蕊(总数,雄蕊长、宽,药长,隔长,雄蕊S、Z数)、雌蕊群(长、径)、心皮数、胚珠数、柱头长、雌蕊(总数,雌蕊S、Z数)等。其中雄蕊与心皮每朵花中随机抽取10个测定,然后计算平均值。对观察记录的数据整理后进行方差分析,计算出最小值、最大值、平均值、标准差、变异系数,株内均方和自由度、株间均方和自由度、F值。并针对性状的大小变化区间作出变异式样图。变化区间分为10个等级,(Xmax-Xmin)/10为每2个区间差,具体数据根据实际情况适当调节。同时,为方便分析形态性状的变异,将形态性状的不同状态进行数字化编码。
2结果与分析
2.1花部数量变异
通过对紫玉兰和二乔玉兰的花部性状观察记录得出,两者在数量上都没发生变异的性状有:花序梗数、雌蕊群数和花被片轮数,其数量分别为1个、1个和3轮。从花部数量的变异系数来看(表1),紫玉兰花部的花梗数没有发生变化,最为稳定,其数量为1;而二乔玉兰的花被片总数、第1轮花被片数、第2轮花被片数、第3轮花被片数和胚珠数是没有变化的,其数量分别为9片、3片、3片、3片和2粒。其它花部数量性状中,变异系数在25%以下的排序为:第1轮花被片数(Z)<第2轮花被片数(Z)<花被片总数(Z)<第3轮花被片数(Z)<心皮数(R)<雄蕊数(R)<心皮数(Z)<雌蕊Z数(R)<雄蕊数(Z)<雄蕊S数(Z)<雄蕊Z数(R)<雄蕊Z数(Z)<雌蕊S数(R)<雄蕊S数(R)<雌蕊S数(Z)<胚珠数(Z);变异系数在25%以上的排序为:雌蕊Z数(Z)<花梗数(R),其中二乔玉兰花梗数的变异系数最大,其值为33.9%。
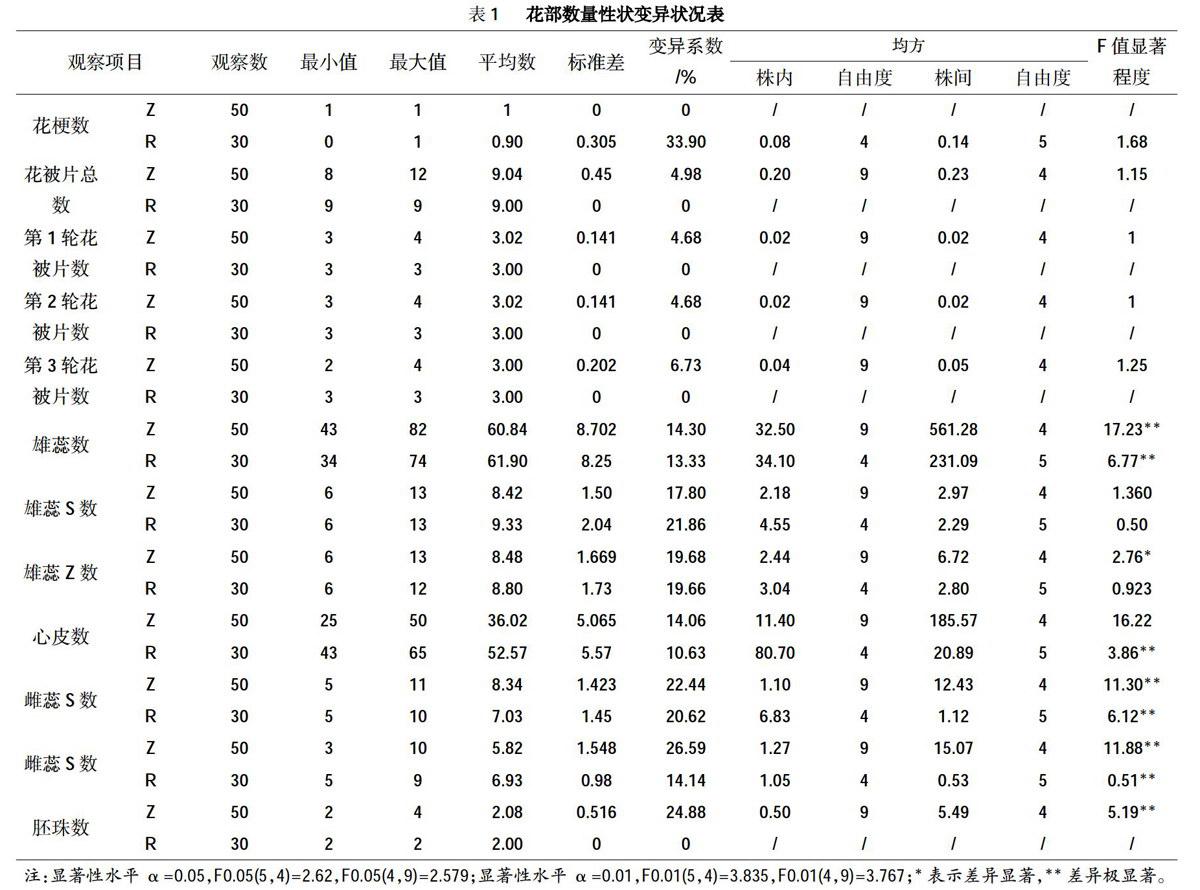
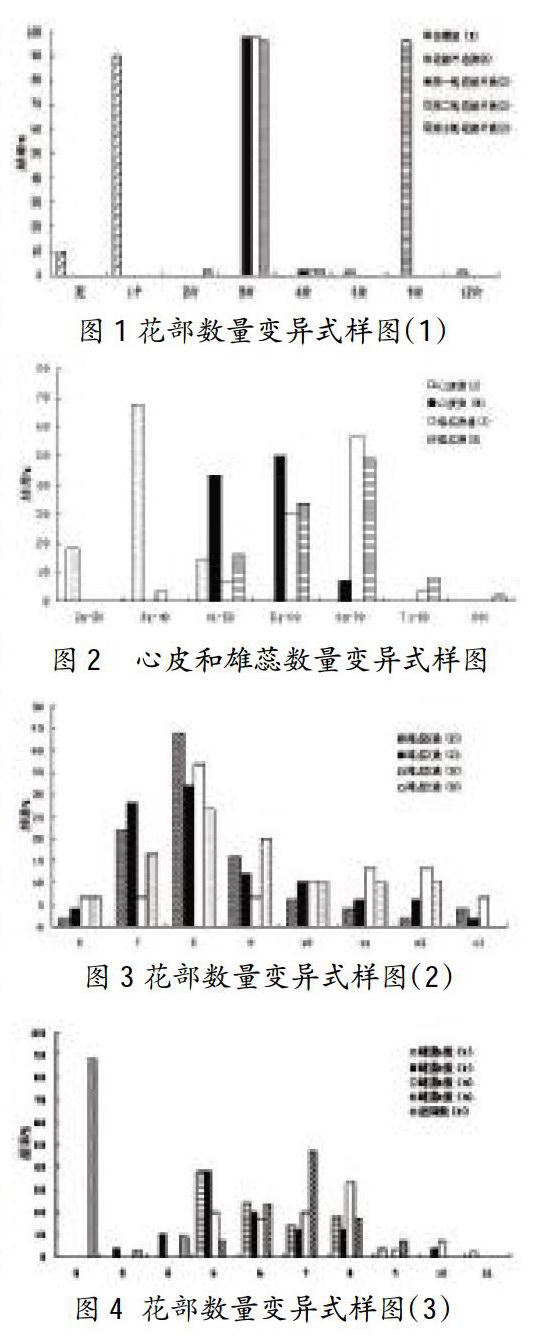
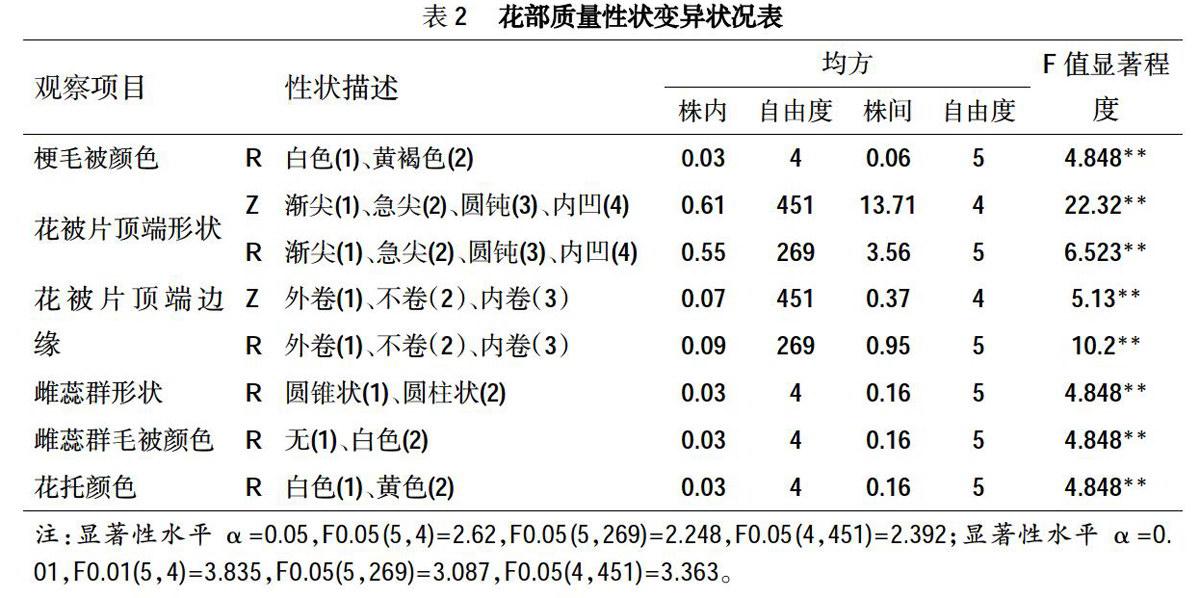
从花部数量性状的变异式样图(图1)可以看出,在花梗数量中,二乔玉兰出现花梗的频率为90%,而没有花梗的频率为10%。在花被片数量中,紫玉兰的总花被片8片的出现频率为2%,9片为96%,12片为2%;第1轮与第2轮花被片数出现频率相一致,3片的出现频率为98%,4片为2%;而第3轮花被片数中2片的出现频率为2%,3片为96%,4片为2%。在雄蕊数量中,紫玉兰雄蕊数的出现频率在30%以下的排序为:31~40枚和71~80枚<41-50枚,出现频率在30%以上的排序为:51~60枚<61~70枚;而二乔玉兰雄蕊数的出现频率在30%以下的排序为:大于80枚<71~80枚<41-50枚,出现频率在30%以上的排序为:51~60枚<61-70枚。在心皮数量中,紫玉兰心皮数的出现频率在30%以下排序为:41-50枚<21-30枚,而31~40枚的出现频率达到68%;二乔玉兰心皮数的出现频率在30%以下的只有61~70枚,而出现频率在30%以上的排序为41-50枚<51-60枚。关于雄蕊S、Z的数量,紫玉兰的雄蕊S数出现频率在20%以下的排序为:6列和12列<11列和13列<10列<9列,出现频率在20%以上的排序为:7列<8列;雄蕊z数的出现频率在20%以下的排序为:13列<6列<11列和12列<10列<9列,出现频率在20%以上的排序为:7列<8列;二乔玉兰的雄蕊S数出现频率在20%以下的排序为:6列和7列<9列和13列<10列<11列和12列,而8列的出现频率为36.6%;雄蕊z数出现频率在20%以下的排序为:6列<10列、11列和12列<7列,出现频率在20%的排序为:9列<8列。在雌蕊S、Z数量中,紫玉兰雌蕊s数的出现频率在20%以下的排序为:11列<9列<7列<8列,出现频率在20%以上的排序为:6列<5列;雌蕊z数出现频率在20%以下的排序为:10列和3列<4列<7列和8列,出现频率在20%以上的排序为:6列<5列;二乔玉兰的雌蕊s数的出现频率在20%以下的排序为:9列<10列<6列,出现频率在20%以上的排序为:5列和7列<8列;雌蕊Z数的出现频率在20%以下的排序为:5列和9列<8列,出现频率在20%以上的排序为:6列<7列。胚珠数量中,紫玉兰的胚珠数2颗的出现频率达到88%,3颗的为3%,4颗的为9%。
从花部数量性状的均方和F值来看(表1),株内变异的均方大于株间变异均方的数量性状有:二乔玉兰的雄蕊S数、雄蕊Z数、心皮数、雌蕊S数和雌蕊Z数,其中心皮数和雌蕊s数的变异极显著,其他3项数量性状的变异都不显著;紫玉兰的第1轮花被片和第2轮花被片数量的株内变异与株间变异均方相等,这2项数量性状的变异都不显著;株内变异的均方小于株间变异的均方的数量性状有:紫玉兰的花被片总数、第3轮花被片数、雄蕊数、雄蕊S数、雄蕊Z数、心皮数、雌蕊S数、雌蕊Z数和胚珠数,二乔玉兰的花梗数和雄蕊数,其中紫玉兰的花被片总数、第三轮花被片数和雌蕊S数以及二乔玉兰的花梗数和雄蕊数的变异都不显著,紫玉兰的雄蕊Z数的变异显著,而紫玉兰的雄蕊数、心皮数、雌蕊S数、雌蕊Z数和胚珠数的变异都极显著。
2.2花部质量性状变异
根据观察可得出紫玉兰花部质量性状如下:花形状为钟状,花颜色是紫红色或紫色的;花枝上密被白色短绒毛;花被片的形状为披针形或倒卵形,外轮花被片颜色为紫绿色,内轮花被片颜色为紫色或紫红色,腹面白色,花被片上無被毛,花被片顶端形状为渐尖或急尖或圆钝或内凹,顶端边缘为外卷或不卷或内卷;雄蕊形状为楔状,花药颜色为黄色,花隔颜色为紫红色;雌蕊群形状为圆柱状,颜色为绿色,并无被毛;花托颜色为淡黄色,并无被毛;心皮软而薄;柱头颜色为紫色或紫红色。二乔玉兰的花部质量性状为:花形状为钟状,花颜色为紫色;花枝上密被白色或黄褐色短绒毛;花被片的形状为倒宽卵圆形或倒卵形,外轮花被片颜色为紫色或深紫色,腹面白色,内轮花被片颜色为紫色或深紫色,腹面白色,花被片上无被毛,顶端形状为渐尖或急尖或圆钝或内凹,顶端边缘为外卷或不卷或内卷;雄蕊形状为楔状,花药颜色为黄色,花隔颜色为紫色;雌蕊群形状为圆锥状或圆柱状,颜色为紫色,被毛无或白色绒毛;花托颜色为白色或黄色,并无被毛;心皮软而薄;柱头颜色为紫色。
从形状性状的变异式样图(图5)可以看出,紫玉兰的花部质量性状的变异情况如下:花被片顶端形状中,形状为渐尖的出现频率为37.6%,急尖的为44%,圆钝的为12%,内凹的为6.4%;花被片的顶端边缘为外卷的出现频率为5.3%,不卷的为92.4%,内卷的为2.3%。二乔玉兰的花部质量性状的变异情况为:花被片的顶端形状中为渐尖的出现频率为28.5%,急尖的为46.7%,圆钝的为22.5%,内凹的为2.3%;花被片的顶端边缘为外卷的出现频率为8.1%,不卷的为88.9%,内卷的为3%。从颜色性状的变异式样图(图6)可得出,二乔玉兰的梗毛被的颜色中,颜色为黄色的出现频率为16.7%,白色的为83.3%;雌蕊群毛被中,颜色为白色的出现频率为16.7%,无毛被的出现频率为83.3%;花托颜色中,颜色为黄色的出现频率为16.7%,白色的为83.3%。
从花部质量性状的均方和F值来看(表2),本研究所列出的6个质量性状均表现出株内变异小于株间变异,并且其变异均表现出极显著。
2.3与近缘种的变异比较
如众所知,在花部结构中,花被片数及其基数是双向演化的,即花被片数量及其基数为中是较原始的,而花被片数量及其基数为少或多是较进化的;而雄蕊和心皮在数量上是单向演化的,即雄蕊和心皮数量为多是较原始的,而雄蕊和心皮数量为少是较进化的。
同已报道的同属木兰科的鹅掌楸(Liriodendron chinense Sargent)、焕镛木(Woonyoungia septentrionalis Law)和大果木莲(Manglietia grandis Hu et Cheng)相比较(如表3),就花部数量特征而言,紫玉兰的雄蕊数量特征较鹅掌楸的原始,但其心皮和花被片数量特征较鹅掌楸进化;而二乔玉兰除心皮数量特征较鹅掌楸进化外,其他均比其原始。而与焕镛木相比,紫玉兰的心皮数较原始外,雄蕊数量特征较进化,花被片数量特征相差不大;而二乔玉兰中出雄蕊数较进化,其他花部特征均表现较原始。与大果木莲相比,紫玉兰的花部特征都较进化;二乔玉兰中雄蕊和心皮数量特征较进化,而花被片数量特征较原始。紫玉兰与二乔玉兰相比,二乔玉兰的雄蕊数量特征比紫玉兰较进化外,其他花部特征均表现较原始。
3讨论
3.1花部数量特征
从上述研究表明,紫玉兰的花形状为钟状,花颜色为紫红色或紫色;花枝密被白色短绒毛;花被片形状为披针形或倒卵形,花被片8~12片,外轮花被片颜色为紫绿色,内轮花被片颜色为紫色或紫红色,腹面白色;花被片上无被毛,花被片顶端形状为渐尖或急尖或圆钝或内凹,顶端边缘为外卷或不卷或内卷;雄蕊形状为楔状,花药颜色为黄色,花隔颜色为紫红色;雌蕊群形状为圆柱状,颜色为绿色,无被毛;花托颜色为淡黄色,无被毛;心皮软而薄;柱头颜色为紫色或紫红色;胚珠2~4颗。二乔玉兰的花形状为钟状,花颜色为紫色;花枝上密被白色或黄褐色短绒毛;花被片的形状为倒宽卵圆形或倒卵形,花被片9片,外轮花被片颜色为紫色或深紫色,腹面白色;内轮花被片颜色为紫色或深紫色,腹面白色;花被片上无被毛,花被片顶端形状为渐尖或急尖或圆钝或内凹,花被片顶端边缘为外卷或不卷或内卷;雄蕊形状为楔状,花药颜色为黄色,花隔颜色为紫色;雌蕊群形状为圆锥状或圆柱状,颜色为紫色,被毛无或白色绒毛;花托颜色为白色或黄色,并无被毛;心皮软而薄;柱头颜色为紫色;胚珠2颗。
在刘玉壶(1996)中国植物志木兰科分类系统中,紫玉兰的描述与其基本相一致,而二乔玉兰中,花被片数量出入相对比较大,刘玉壶系统记载二乔玉兰的花被片6~9片,而本研究的结果都为9片,这有可能是他们观察的树种发生了比较大的变异,而本文所观察的树种性状比较稳定,而未出现花被片6~9片的情况;或者是当初并没观察到花期,而是通过花被片的瓣痕判断以致出现了误差。而司马永康(2011)最新研究报道紫玉兰花被片9~16片,也与本文的8~12片有所出入,可能是由于观察的年份不同,而近几年昆明出现了连续3年的干旱天气,这对植物的生长发育是有或大或小的影响,从而造成紫玉兰花被片变少的结果。
3.2花部变异特点
在不同性状间,存在数量和质量变异差异显著的花部结构。在紫玉兰中,变异系数最大的数量性状为雌蕊z数,其值为26.59%;二乔玉兰的变异系数最大的数量性状为花梗数,其值为33.9%;其他数量性状的变异系数都在25%以下。可以看出紫玉兰和二乔玉兰发生的变异都相对较小,性状相对比较稳定。对比紫玉兰和二乔玉兰发生的变异,也可以看出二乔玉兰的花被片数量特征要较紫玉兰稳定,而花梗、雄蕊和心皮的数量特征,紫玉兰要较二乔玉兰稳定。这有可能是因为二乔玉兰是由玉兰和紫玉兰杂交出来的缘故,二乔玉兰在一定程度上继承了紫玉兰相对稳定的性状,并转化成自己特有的性状。
从株间变异和株内变异分析,我们可以看出,二乔玉兰的大部分数量性状表现出株内变异大于株间变异,而紫玉兰的数量性状表现出株内变异等于或小于株间变异。在质量性状中,紫玉兰和二乔玉兰均表现出株内变异小于株间变异。这说明紫玉兰花部性状的变异主要发生在株间,而二乔玉兰花部数量性状主要发生在株内,花部质量性状的变异主要发生在株间。在同一居群中,不同单株表现出不同的变异类型或大小,这些变异有大有小,也就是同一居群中存在优良单株的可能性。我们从花部质量性状变异可以看出,紫玉兰和二乔玉兰的质量性状发生的变异都为极显著,也就是说在这两个品种中,存在许多变异因素,可以根据不同的选育目的,培育出不同性状的品种。比如,在颜色上的选择,可以针对有意向的颜色,对树种进行基因等方面的研究。同时这说明可以在紫玉兰中选育出优良单株,而二乔玉兰则要根据选育的目的与要求,制作不同的选育方案。这可为紫玉兰和二乔玉兰的选育工作提供一些基础数据。
3.3花部性状的演化
一个有机体的不同器官和不同部分的演化速度是不同的,因而造成同一分类群在具有许多原始特征的同时,又具有许多进化特征的现象。这种同一分类群具有不同演化级性状的现象称为特化交叠、镶嵌进化或异步演化。在绝大多数情况下,特别在被子植物的较原始类群中,异步演化表现得最为明显。木兰科一直被公认为一类相对而言较原始或最原始的类群。这在木兰科植物的花部结构中也可观察到异步演化。在所观察的花部性状项目中,通过数据分析,我们可以看出数量变异中单向演化中以少为原始,多为进化的有:紫玉兰的第1轮花被片数、第2轮花被片数和第3轮花被片数;而以多为原始,少为进化的有:紫玉兰的花被片总数、雄蕊数、雄蕊SZ数、心皮数和雌蕊SZ数,二乔玉兰的花梗数、雄蕊数、雄蕊SZ数、心皮数和雌蕊SZ数。性状变异中单向演化的有:梗毛被颜色、雌蕊群形状、毛被颜色和花托颜色;双向演化的有花被片顶端形状、花被片顶端边缘。对于单向演化的花部性状,可为我们选育工作提供更多的一个方向,或许能通过诱发培育更多样的品种。
在木兰科中花部雄蕊和心皮根据其螺旋排列的旋转方向和明显程度分为S、Z和SZ型3种排列方式,而通过数据分析可以看出,紫玉兰和二乔玉兰的S数和Z数都明显的存在。紫玉兰的雄蕊S数主要出现在7和8列(频率在20%以上),Z数也在7和8列(20%以上),心皮S数主要出现在6和5列(20%以上),Z数也在6和5列(20%以上);二乔玉兰的雄蕊S主要出现在8列(频率在20%以上),Z数在9和8列(20%以上),心皮S数在5、7和8列(20%以上),Z数在6和7列(20%以上)。由此可见,推测紫玉兰和二乔玉兰的雄蕊和心皮的主要排列方式是SZ型,其数值是以上数字的组合。其变异是双向演化,少或多为较进化,该性状是相对较稳定的。对比紫玉兰和二乔玉兰,可以看出这2个品种在SZ数和其他花部特征上有其相似性,表明这两个树种具有很强的亲缘性,这也正是二乔玉兰是玉兰和紫玉兰的杂交的结果。
紫玉兰的雄蕊数和二乔玉兰的心皮数保留了一定的原始性,这也许由木兰科植物授粉率低、败育率低和结籽率低造成的。而紫玉兰的花被片数和二乔玉兰的花梗数等是较进化的性状。由此可见,紫玉兰和二乔玉兰的原始性状和进化性状是交织出现,这可能是受地理分布的不同和环境条件的影响,使得他们在演化进程中既保留木兰科中一些较原始的性状,又由环境影响而产生了或多或少的进化性状。总体来说,紫玉兰在木兰科中是较进化的品种,二乔玉兰则保留更多的原始性状,是木兰科中比较原始的树种。
综上分析,可以看出,紫玉兰和二乔玉兰花部性状的变异主要发生在花被片、雄蕊和心皮3个方面。而花部作为一个适用于传粉的功能单位,这些性状变异极有可能跟花粉传递机制相关。而据已有研究可知,植物传粉主要是风媒和虫媒两种方式,而研究发现木兰科植物的雄蕊群包在花被片之内,周围杯状的花被片对花粉散布起阻挡和滞留作用,这限制了风媒传粉的作用。另外,据Collaway(1994)和姜景民等(1999)试验报道含笑属、玉兰属和木兰属,都属于雌蕊先成熟,且在花被片尚未张开之前,即已进入可授粉期,而在花被片张开后已过最佳授粉期,此时尚见有蜜蜂等昆虫钻入花中。可以看出,木兰科植物主要是通过虫媒的方式授粉的。传粉者与花是通过蜜腺、花粉、气味及花色等花部综合特征来维持传递关系。而花色、花形及大小等影响着昆虫的视觉敏感性,那么这些花部特征变异极有可能是为提高种内和种间生存竞争而引起的。而影响结籽率的因素是多方面的,有环境因子,有花的形态结构特征以及胚胎学等因子。通过对这2个品种花部形态变异的研究,更可以看出花被片和雄蕊等性状变异显著极有可能为提高其遗传多样性和进行自我保护机制等。这说明植物的性状变异都是为取得最佳适应性而发生的。
此外,表型性状变异在一定程度上反映了遗传丰富程度。紫玉兰和二乔玉兰的花部特征变异表明木兰科含笑属植物具有较丰富的遺传多样性。至于遗传多样性的分子检测有待进一步探索。