B细胞衔接蛋白BANK在小鼠实验性自身免疫性脑脊髓炎中的作用①
2017-12-20金桂花李红花藤本张庆镐
金桂花 李红花 李 雪 藤本·学 张庆镐
(延边大学医学院免疫学与病原生物学教研室,延吉 133002)
B细胞衔接蛋白BANK在小鼠实验性自身免疫性脑脊髓炎中的作用①
金桂花 李红花 李 雪 藤本·学②张庆镐③
(延边大学医学院免疫学与病原生物学教研室,延吉 133002)
目的探讨B细胞特异性衔接蛋白BANK(B cell adoptor protein with ankyrin repeats)在小鼠实验性自身免疫性脑脊髓炎(Experimental autoimmune encephalomyelitis,EAE)中的作用。方法MOG35-55多肽免疫C57BL/6鼠和BANK缺陷(BANK-deficient,BANK-/-)鼠制备EAE模型,观察实验动物临床症状及中枢神经系统的病理学变化;提取小鼠脑组织及脾脏,经流式细胞术检测中枢神经系统及外周免疫器官中的CD4+T细胞、CD8+T细胞及调节性T细胞的变化。结果BANK-/-鼠EAE临床症状评分明显高于C57BL/6鼠,且体重减轻明显(P<0.05);HE染色结果显示,BANK-/-鼠较C57BL/6鼠炎症感染灶明显增多。流式细胞术结果显示,相对于C57BL/6鼠BANK-/-鼠中枢神经系统中CD8+T细胞百分比明显增多,而脾脏中调节性T细胞百分比明显减少,CD4/CD8比值倒置(P<0.05)。结论B细胞衔接蛋白BANK的表达抑制EAE炎症反应。
B细胞;B细胞衔接蛋白;BANK;EAE
多发性硬化症(Multiple sclerosis,MS)是以中枢神经系统脱髓鞘为特征的自身免疫性疾病,实验性自身免疫性脑脊髓炎(EAE)被认为是研究MS的理想动物模型[1]。长期以来MS被认为是T细胞介导的自身免疫性疾病,然而近来的研究报道,B细胞在MS发病中起到潜在决定性作用,但具体机制尚不明确[2,3]。
细胞的发育、分化和激活是发挥免疫应答功能和免疫调节功能的前提。上述过程的有效实施,依赖于细胞表面的刺激信号,进而启动核转录因子及相关效应分子基因活化,还需要众多细胞内蛋白分子的联动和协同作用。衔接蛋白就是这类参与细胞信号转导调节中的重要分子。衔接蛋白无酶活性,通过参与信号转导中蛋白质与蛋白质间的反应,为大分子复合物的形成提供支架,从而介导胞内信号的整合与传递[4,5]。研究证明,衔接蛋白分子的异常表达会引起免疫功能的异常,甚至免疫疾病的发生。已发现的B细胞衔接蛋白有3种,包括BANK(B-cell adoptor protein with ankyrin repeats),BLNK(B-cell linker protein,BLNK)和BCAP(B-cell adaptor for PI3K)[6-9]。我们前期研究发现,BLNK缺陷导致B细胞功能缺失,导致BLNK-/-鼠EAE症状加重[10]。但是,BANK是否也参与EAE的发病和发展,相关报道罕见。本研究拟采用BANK缺陷鼠(BANK-/-鼠)通过建立EAE小鼠模型,明确B细胞衔接蛋白BANK和EAE发病和发展的相关性,本研究结果有助于揭示免疫调节分子与自身免疫性疾病的内在联系。
1 材料与方法
1.1材料
1.1.1实验动物 野生型C57BL/6鼠和BANK-/-鼠(C57BL/6)鼠购自美国Jackson实验室,采用6~8周龄雌性鼠,实验动物饲养于无特异性病原体的清洁环境中。
1.2方法
1.2.1EAE小鼠模型的制备 将100 μg MOG35-55多肽(MEVGWYRSPFSRVVHLYRNGK)加入100 μl完全弗氏佐剂(含结核杆菌终浓度为2 mg/ml)中乳化,将制成抗原配剂。实验小鼠经皮下注射200 μl抗原配剂,第0天和第2天腹腔注射500 μl百日咳毒素(含200 ng百日咳毒素)制备动物模型[11]。
1.2.2体重变化及临床症状评分 每天观察小鼠的体重变化及临床症状,连续观察28 d。临床症状评分标准如下:0分,不发病;1分,尾部张力低下;2分,步态笨拙或翻身反射消失;3分,双侧后肢部分麻痹;4分,后肢完全麻痹;5分,后肢麻痹伴前肢肌力下降;6分,前肢麻痹或濒死状态[11]。
1.2.3脑和脊髓组织的病理学改变 MOG35-55免疫后第16~18天处死小鼠,打开胸腔先经左心室灌注PBS缓冲液,然后缓慢灌注预冷4%多聚甲醛固定液后取脑和脊髓组织。组织经福尔马林固定,石蜡包埋用于苏木精伊红(HE)染色,观察病理学改变及炎症细胞浸润。HE染色标本在400倍光镜下计数炎症细胞,实质内浸润的炎性细胞数等于或超过20个的区域作为一个炎症感染灶。
1.2.4脑组织中淋巴细胞的分离 诱导EAE后的第18天处死小鼠,经左心室灌注50 ml PBS缓冲液后取小鼠脑组织。组织经胶原酶D(Sigma-Aldrich)和DNase Ⅰ(Sigma-Aldrich)消化45 min,采用Percoll不连续密度梯度沉淀法(70/37%)分离纯化脑组织中的淋巴细胞。分离纯化的淋巴细胞制备细胞悬液(1×106cell/ml)用于流式细胞术的检测。
1.2.5淋巴细胞的流式细胞术检测 诱导EAE后的第18天处死小鼠制备淋巴细胞悬液。染色用荧光抗体包括Thy1.2(53-2.1)、CD4(RM4-5)、CD8(53-6.7)、CD25(PC61)及Foxp3(FJK-16s)。调节性T细胞的细胞内染色采用Fixation/Permeabiliz-ation(eBioscience)进行破膜固定。应用同种型对照免疫球蛋白作为背景染色。数据分析使用FlowJo软件。

2 结果
2.1临床症状评分及体重的变化 如图1所示,BANK-/-鼠临床评分始终明显高于野生鼠(P<0.01),且临床症状减轻缓慢,小鼠自出现临床症状开始出现食欲下降,继而出现体重减轻(结果未显示)。在疾病的高峰期BANK-/-鼠临床评分为(3.5±0.2),与野生鼠相比具有显著统计学意义(2.8±0.3,P<0.01)。见图1、表1。
2.2动物发病情况 小鼠平均第9~11天开始出现临床症状,第15~19天症状达高峰,随着时间的推移,症状可自行缓解。大部分小鼠以平衡失调为首发症状,个别小鼠以尾部张力下降为首发症状,继而小鼠出现后肢无力拖地、后肢麻痹、前肢麻痹、抽搐等症状。对照组和实验组小鼠发病率均为80%以上,死亡率无统计学意义(P> 0.05)。见表1。

图1 BANK的缺失加重EAE反应Fig.1 Enhanced EAE response in BANK-/-miceNote: EAE were induced by MOG peptide immunization in wild-type and BANK-/-mice.Clinical score(±s)from ≥ 10 mice in each group.Significant differences between means of EAE clinical scores are indicated.*.P<0.05.
表1EAE临床症状评分
Tab.1EAEclinicalscoresfollowingMOGtreatmentA

GroupsIncidenceBMortalityMeandayofonsetMeanmaxiumscoreWild-type9/11(81.8%)1/11(9.1%)10.8±0.52.8±0.3BANK-/-12/12(100%)2/12(16.7%)10.5±0.33.5±0.21)


图2 野生鼠和BANK-/-鼠中枢神经系统的组织病理学变化Fig.2 Histopathology of CNS in wild-type and BANK-/-miceNote: A.Brain;B.Spinal cord.Representative brain and spinal cord sections harvested 17 days after MOG immunization by HE staining.Top and bottom panels are low and high magnifications,respectively.Arrows point to the foci of inflammatory area.Similar results were obtained in at least 2 independent experiments.
2.3脑和脊髓组织的病理学改变 HE染色结果显示,BANK-/-鼠中枢神经系统中炎症感染灶明显多于C57BL/6鼠,浸润的细胞以淋巴细胞为主。在炎性细胞浸润明显的区域可见脊髓有不同程度的丢失,见图2。
2.4脑组织中CD4+T细胞、CD8+T细胞与调节性T细胞的变化 流式细胞仪检测结果显示,相对于野生鼠BANK-/-鼠脑组织中CD8+T细胞百分比明显增多,两组小鼠之间具有统计学意义(P<0.05),而调节性T细胞(regulatory T cell,Treg)数量无明显改变(P> 0.05),见图3。
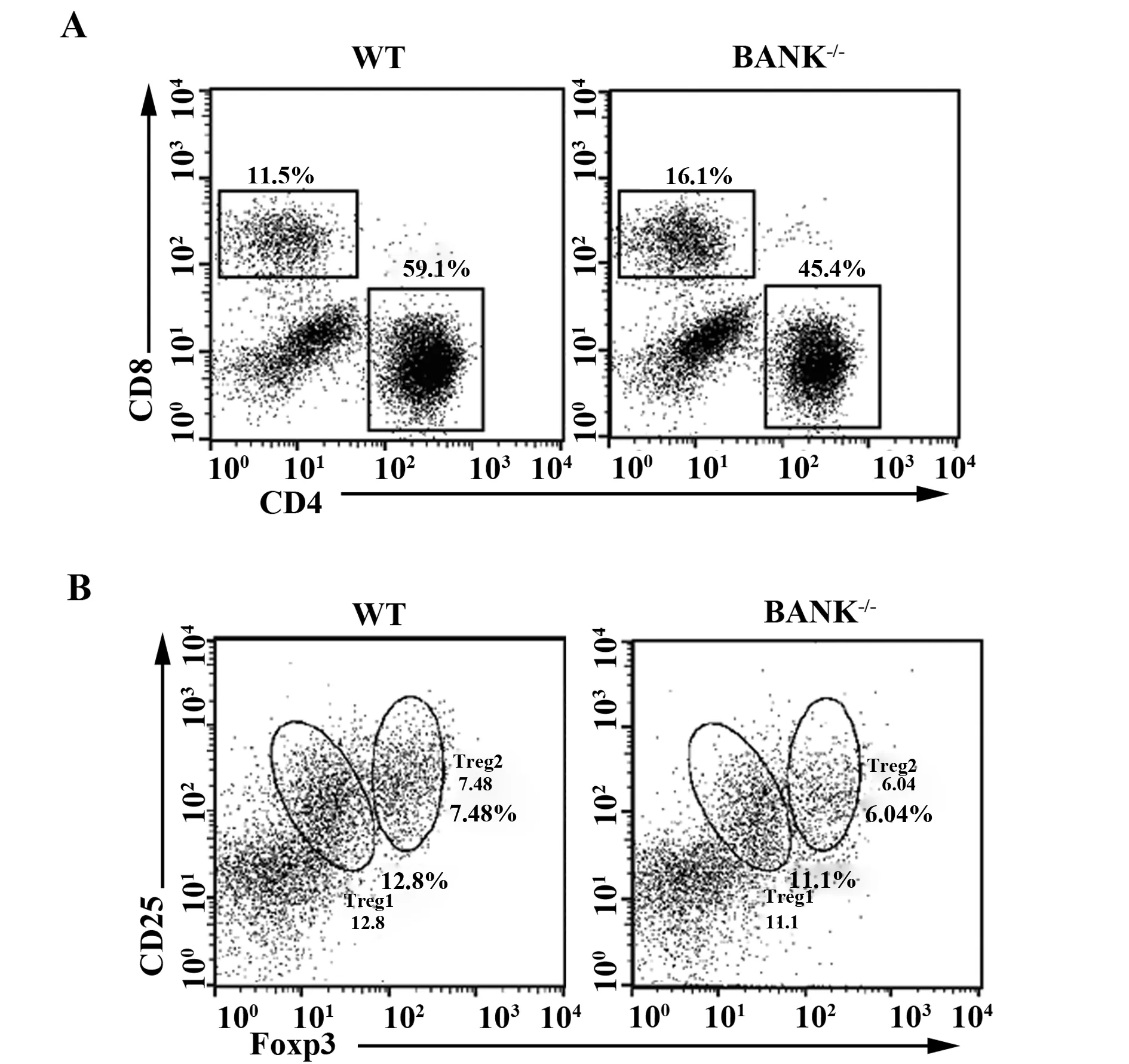
图3 野生鼠和BLNK-/-鼠脑组织中淋巴细胞的浸润Fig.3 Lymphocyte infiltration from spleen in wild-type and BANK-/-miceNote: CD4+ T cells and CD8+ T cells(A)and CD4+CD25+ FoxP3+ regulatory T(Treg)cells(B)from brain were analyzed by flow cytometry.

图4 野生鼠和BLNK-/-鼠脾脏中淋巴细胞的浸润Fig.4 Lymphocyte infiltration from spleen in wild-type and BANK-/-miceNote: CD8+ T cells,CD4/CD8 ratio and CD4+CD25+ FoxP3+ regulatory T(Treg)cells from spleen were analyzed by flow cytometry.Data were obtained from five mice in each group.
2.5脾脏中CD4+T细胞、CD8+T细胞与Treg的变化 流式细胞仪检测结果显示,较C57BL/6鼠BANK-/-鼠CD8+T细胞百分比明显增多(P<0.05)CD4/CD8比值倒置,而Treg百分比显著下降(P<0.05)。见图4。
3 讨论
MS好发于中青年,致残率高、易复发,病因及发病机制尚不明确。免疫学因素被认为是MS发病的关键因素,长期以来占主流地位的是T细胞介导的致病理论,这主要基于MS患者中枢神经系统脱髓鞘病灶周围有大量T细胞浸润的病理学证据[1]。然而近来研究显示,B细胞在MS发病中同样发挥着重要作用。B淋巴细胞去除药物利妥昔单抗治疗MS患者可以取得良好临床效果[12],是支持B细胞免疫参与MS发病的客观证据。对继发进展型MS患者脑组织的尸检研究发现,其脑膜上有异位的淋巴滤泡样结构形成,并与次级淋巴组织的生发中心(Germinal center,GC)有着相似的组织结构和B细胞克隆扩增,异位滤泡样结构与患者皮层病变的严重程度和临床病程的进展呈正相关[2,3]。
BANK(B-cell adoptor protein with ankyrin repeats)是蛋白酪氨酸激酶的底物,特异性表达于B细胞。BANK通过调节Akt的活化,从而抑制过剩的B细胞反应[13]。Durand等[14]证实,抗CD40的刺激信号促使B细胞高表达BANK1分子,同时上调Granzyme B和Irf4对免疫应答起到负向免疫调节作用。越来越多的研究证实,BANK与自身免疫性疾病易感性密切相关。对系统性红斑狼疮患者的研究中发现,BANK1基因(sr3733197、sr3733197和sr17266594位点)多态性与疾病易感性密切相关,提出BANK1可以作为疾病易感性的遗传标记[15,16]。Orozco等[17]证实类风湿性关节炎患者BANK1基因位点也与关节炎易感性相关。Rueda等[18]和Dieudé等[19]证实BANK1基因多态性与系统性硬化症的疾病易感性相关,并与患者血清中抗拓扑异构酶-Ⅰ抗体的增加成正相关。近来,Zouidi等[20]究提出BANK1和IL-15可以作为Ⅰ型糖尿病的新型易感基因。但是,目前为止,对于BANK在自身免疫性疾病中的作用多限于基因多态性的研究,在蛋白质水平上研究BANK通过何种机制参与自身免疫性疾病病程的相关报道非常罕见。因此,研究BANK在MS中的作用及机制,对于阐明MS的发病机理、寻求治疗MS的治疗靶点具有重要的指导意义。
本实验中我们探讨了B细胞特异性衔接蛋白BANK在EAE发生和发展中的作用。实验结果显示,随着疾病的进展BANK-/-鼠EAE临床评分急剧增高,临床评分始终明显高于野生鼠,且病程延长(图1、表1)。小鼠脑组织及脊髓组织病理变化也显示,BANK-/-鼠中枢神经系统中炎症细胞浸润及脱髓鞘等改变比野生鼠更为明显(图2)。以上结果提示BANK参与EAE的发病和发展。
在MS发病过程中,CD4+CD25+调节性T细胞数量及抑制T细胞的效应功能均有所下降[21]。我们的实验结果显示,相对于C57BL/6鼠,BANK-/-鼠脑组织中CD8+T细胞百分比显著增多(图3)。中枢神经炎症灶中并未发现B细胞及中性粒细胞的浸润(数据未显示),而在诱导EAE之前,两组小鼠的T细胞数量及百分比差异是无统计学意义(P> 0.05,数据未显示)。该研究结果表明,CD8+T细胞在EAE炎症灶中选择性的增多,提示BANK的表达参与效应性T细胞的功能。流式细胞术进一步显示,BANK-/-鼠表现为CD4/CD8 T细胞比值倒置,Treg百分比相对于野生鼠明显减少(图4)。因此,我们设想在EAE中B细胞可能通过促进Treg的增殖维持其数量,使得Treg更好地缓解EAE炎症反应。但是,BANK通过何种机制参与B细胞的功能以及BANK是否通过抑制T细胞的分化及增殖,并通过抑制促炎症因子的分泌参与EAE的病程,相关机制有待进一步阐明。
综上所述,我们采用BANK-/-鼠通过建立复发-缓解型EAE小鼠模型,明确了BANK的表达抑制EAE炎症反应。本研究结果为进一步阐明BANK在EAE中的作用机制奠定了理论基础,并为MS治疗提供了新的治疗靶点。
[1] Compston A,Coles A.Multiple sclerosis[J].Lancet,2008,372:1502-1517.
[2] Magliozzi R,Howell O,Vora A,etal.Meningeal B-cell follicles in secondary progressive multiple sclerosis associate with early onset of disease and severe cortical pathology[J].Brain,2007,130:1089-1104.
[3] Serafini B,Rosicarelli B,Magliozzi R,etal.Detection of ectopic B-cell follicles with germinal centers in the meninges of patients with secondary progressive multiple sclerosis[J].Brain Pathol,2004,14:164-174.
[4] Kurosaki T.Regulation of B-cell signal transduction by adaptor proteins[J].Nat Rev Immunol,2002,2:354-363.
[5] Kurosaki T,Hikida M.Tyrosine kinases and their substrates in B lymphocytes[J].Immunol Rev,2009,228:132-148.
[6] Aiba Y,Yamazaki T,Okada T,etal.BANK negatively regulates Akt activation and subsequent B cell responses[J].Immunity,2006,24:259-268.
[7] Nakayama J,Yamamoto M,Hayashi K,etal.BLNK suppresses pre-B-cell leukemogenesis through inhibition of JAK3[J].Blood,2009,113:1483-1492.
[8] Koretzky GA,Abtahian F,Silverman MA.SLP76 and SLP65:complex regulation of signalling in lymphocytes and beyond[J].Nat Rev Immunol,2006,6:67-78.
[9] Troutman TD,Hu W,Fulenchek S,etal.Role for B-cell adapter for PI3K(BCAP)as a signaling adapter linking Toll-like receptors(TLRs)to serine/threonine kinases PI3K/Akt[J].Proc Natl Acad Sci U S A,2012,109:273-278.
[10] Jin G,Hamaguchi Y,Matsushita T,etal.B-cell linker protein expression contributes to controlling allergic and autoimmune diseases by mediating IL-10 production in regulatory B cells[J].J Allergy Clin Immunol,2013,131:1674-1682.
[11] 金桂花,李雪,藤本·学,等.B细胞连接蛋白的表达抑制小鼠实验性自身免疫性脑脊髓炎[J].免疫学杂志,2016,32(10):852-863.
[12] Hauser SL,Waubant E,Arnold DL,etal.B-cell depletion with rituximab in relapsing-remitting multiple sclerosis[J].N Engl J Med,2008,358:676-688.
[13] Aiba Y,Yamazaki T,Okada T,etal.BANK negatively regulates Akt activation and subsequent B cell responses[J].Immunity,2006,24:259-268.
[14] Durand J,Huchet V,Merieau E,etal.Regulatory B cells with a partial defect in CD40 signaling and overexpressing Granzyme B transfer allograft tolerance in rodents[J].J Immunol,2015,195:5035-5044.
[15] Castillejo-López C,Delgado-Vega AM,Wojcik J,etal.Genetic and physical interaction of the B-cell systemic lupus erythematosusassociated genes BANK1 and BLK[J].Ann Rheum,2012,71:136-142.
[16] Kozyrev SV,Abelson AK,Wojcik J,etal.Functional variants in the B-cell gene BANK1 are associated with systemic lupus erythematosus[J].Nat Genet,2008,40:484.
[17] Orozco G,Abelson AK,González-Gay MA,etal.Study of functional variants of the BANK1 gene in rheumatoid arthritis[J].Arthritis Rheum,2009,60:372-379.
[18] Rueda B,Gourh P,Broen J,etal.BANK1 functional variants are associated with susceptibility to diffuse systemic sclerosis in Caucasians[J].Ann Rheum Dis,2010,69:700-705.
[19] Dieudé P,Wipff J,Guedj M,etal.BANK1 is a genetic risk factor for diffuse cutaneous systemic sclerosis and has additive effects with IRF5 and STAT4[J].Arthritis Rheum,2009,60:3447-3454.
[20] Zouidi F,Stayoussef M,Bouzid D,etal.Association of BANK1 and cytokine gene polymorphisms with type 1 diabetes in Tunisia[J].Gene,2014,536:296-301.
[21] Severin ME,Lee PW,Liu Yue,etal.MicroRNAs targeting TGFβ signalling underlie the regulatory T cell defect in multiple sclerosis[J].Brain,2016,139(6):1747-1761.
InhibitoryroleofBANKinprogressionofexperimentalautoimmuneencephalomyelitis
JINGui-Hua,LIHong-Hua,LIXue,FUJIMOTOManabu,ZHANGQing-Gao.
DepartmentofImmunologyandPathogenicBiology,CollegeofBasicMedicine,YanbianUniversity,Yanji133002,China
Objective:To investigate the role of B cell adoptor protein with ankyrin repeats(BANK)in experimental autoimmune encephalomyelitis(EAE).MethodsC57BL/6 mice and BANK-deficient(BANK-/-)mice were immunized with MOG peptide in CFA,and then observed the clinical symptoms and pathological severity.ResultsThe percentages of CD4+T cells,CD8+T cells and regulatory T cells in brain and spleen were analyzed by flow cytometry.BANK-/-mice showed significantly higher score at the peak and the plateau phase compared with wild-type mice(P<0.05).HE staining showed more widespread areas of inflammation and demyelination in BANK-/-mice when compared to wild-type mice on day 16.In addition,the frequency of CNS-infiltrating CD8+T cells was markedly higher in BANK-/-mice than in wild-type mice.In addition,the percentage of CD8+T cells from spleen in BANK-/-mice was also increased compared with wild-type mice(P<0.05).By contrast,the percentage of regulatory T cells and the ratio of CD4/CD8 T cells from spleen in BANK-/-mice were significantly lower than in wild-type mice(P<0.05).ConclusionThus,BANK expression in B cells can inhibit the development of EAE.
B cell;B cell adoptor protein;BANK;EAE
10.3969/j.issn.1000-484X.2017.12.020
R392.12
A
1000-484X(2017)12-1854-05
①本文为国家自然科学基金项目(81460255,61671098)。
②日本筑波大学医学医疗系皮肤科,筑波県 つくば市 3058575。
③大连大学医学院,大连 116622。
金桂花(1975年-),女,博士,副教授,硕士生导师,主要从事蛋白表达调控与自身免疫性疾病、B细胞方面的研究,E-mail:ghjin@ybu.edu.cn。
及指导教师:张庆镐(1964年-),男,博士,教授,博士生导师,主要从事蛋白表达调控与自身免疫性疾病、B细胞方面的研究,E-mail:zqg0621@ybu.edu.cn。
[收稿2017-09-27]
(编辑 许四平 刘格格)