葡萄膜黑色素瘤差异表达基因的富集分析
2017-11-22孙晓雨杨春花
孙晓雨 杨春花
葡萄膜黑色素瘤差异表达基因的富集分析
孙晓雨 杨春花
目的从基因转录组水平揭示葡萄膜黑色素瘤的发病机制,为临床诊疗提供新工具。方法在公共基因芯片数据库GEO中找到葡萄膜黑色素瘤的相关基因芯片数据,数据经以2为底的对数转换及标准化处理后,找到黑色素瘤中的差异表达基因。通过DAVID基因功能注释和功能分类对这些基因进行生物学和统计学分析。结果在葡萄膜黑色素瘤组织中共找到265个差异表达基因,包括上调基因95个,下调基因170个。基因富集分析结果显示,这些基因的功能大致分为细胞运输、细胞骨架结构、细胞周期、信号识别及转导、细胞分化调控等。结论利用生物信息学的方法能有效分析基因芯片数据并获取生物内在信息,黑色素瘤组织中多种基因的表达发生了改变,为确定黑色素瘤的早期诊断标志与新治疗靶位提供了新的思路。
葡萄膜黑色素瘤;差异表达;基因芯片;GEO;生物信息学
作者单位:250022 济南,山东济南循证医学科学研究中心基础医学部
视网膜母细胞瘤(retinoblastoma,Rb)是婴幼儿最常见的眼内恶性肿瘤[1],对视力和生命均构成了严重的威胁和危害。葡萄膜黑色素瘤(uveal melanoma),是成年人眼部发病率最高的恶性肿瘤,主要来源于葡萄膜黑色素细胞,具有增殖活性高和易于转移等特点,容易与许多眼底疾病混淆。葡萄膜黑色素瘤的死亡率很高,主要原因是通过血液循环系统发生转移的几率很高,主要致死原因是肝脏转移[2]。一旦确诊肿瘤转移,预后极差,数月内可致患者死亡[3]。鉴于其恶性程度高,是否能做到早期诊断对挽救患者的生命显得尤为重要。
近年的研究证明,一些分子途径与葡萄膜黑色素瘤的进展有关,其中GNAQ基因、GNA11基因、干细胞因子受体(c-Kit)、肝细胞生长因子(c-Met)以及微小RNA(miR34a)都通过调节特定的分子途径影响肿瘤的进展[4-6]。张旭等[7]在高侵袭转移性黑色素瘤细胞中发现骨桥蛋白的表达水平明显升高,可作为预测葡萄膜黑色素瘤侵袭能力、转移潜能以及患者预后的指标。但是并未进行转录组水平的高通量检测,其应用仍有较长的距离。
因此,为了给黑色素瘤的防治提供筛查标志及新的治疗靶点,从基因转录组水平揭示其发病机制至关重要。基因芯片作为一种高效、大规模获取生物信息的技术,能检测和分析肿瘤组织与正常组织的差异表达基因。本研究利用公共基因芯片数据库GEO中的芯片数据,对黑色素瘤的相关基因挖掘并进行生物信息学分析。
1 材料和方法
1.1 基因表达数据 在GEO(http://www.ncbi.nlm.nih.gov/geo/)数据库中选择GSE24673芯片数据进行数据分析。GSE24673由Krishnakumar等人提供,芯片平台为GPL6244(Human Gene 1.0 ST Arrays)。本研究对该平台的所有样本进行分析,分别是GSM607938、GSM607939、GSM607940、GSM607941、GSM607942、GSM607943、GSM607944、GSM607945、GSM607946、GSM607947和GSM607948。
1.2 实验过程 实验类型为芯片表达分析。实验中对原发性视网膜肿瘤中退化的和分裂的RNA样品进行了转录组分析。全局基因表达谱分析在对视网膜细胞瘤的早期诊断标记的确定方面具有巨大的应用前景。实验中分析了3个原发性视网膜肿瘤组织(一式三份)和2个正常健康成年人的视网膜组织中的基因表达水平。视网膜细胞瘤RNA的用量使用纳克级别,在Human Gene 1.0 ST Arrays进行处理。该芯片数据含实验组9个,GSM607947和GSM607948作为对照组,参照人类健康视网膜组织中全转录本。
1.3 数据预处理及差异表达基因的筛选 使用GEO2R读取芯片数据[8],得到28,869条基因数据,对表达数据进行log2转换,使基因的表达值转化为正态分布。之后对数据进行过滤及分位数法标准化。
limma包用于R分析差异表达基因,该模型将标准及信号强度的关系使用线性模型进一步强化,基于贝叶斯方法来确定差异表达基因,准确率较高,是目前使用最广泛的方法[9]。利用limma包提取线性模型中的差异表达基因,运用t检验(公式1)判断对照组和实验组基因的表达差异是否具有显著性。

其中均值

方差

式中ni为某一条件下的实验重复次数,Xij为某基因在第i个条件下第j次重复实验的表达水平测量值。根据统计量t值,得到P值。若P值小于0.05则认为某基因在两个不同条件下表达差异具有统计学意义。
运用Benjamini&Hochberg算法[10]对样本P值进行多重检验FDR校正(阈值为0.05)(公式2)。筛选|logFC|>2且P<0.05的基因视为差异表达基因。

公式中V表示不可观察的随机变量,R表示可观察的随机变量。
1.4 差异表达基因的富集分析 将得到的差异表达基因通过Fisher精确检验(FisherExact Test)进行富集分析,找到这些基因的功能特点及可能参与的生物学过程,富集分析对应的公式为:

上式中,N表示芯片上所有基因总数,n表示N中差异表达基因的总数,M表示N中属于某个GO term的基因个数,k表示n中属于某个GO term的基因个数。P值表示差异表达基因富集到这个GO term上的可信程度,本实验设定P小于阈值0.05时,认为差异表达基因显著性的富集到这个GO term上。
本实验对差异表达基因从功能分类(functional categories)和蛋白质结构域(protein domains)两个方面进行生物学分析。
1.5 差异表达基因的聚类分析 应用DAVID(The Database for Annotation,Visualization and Integrated Discovery)[11]中Functional Annotation Clustering,利用EASE检验对差异表达基因进行聚类分析,确定具有相同表达模式的基因,从而预测这些差异表达基因与黑色素瘤发生的关系。
2 结果
2.1 差异表达基因的筛选结果 使用GEO2R在线软件对芯片中的数据进行差异表达分析,共筛选得到265个满足条件的差异表达基因,包括上调基因95个,下调基因170个。图1列举了前20个差异表达较显著的基因,主要有:DCT(P=2.4E-08)、GUCY2F(P=9.9E-08)、TOP2A(P=3.3E-06)、PRAME(P=4.3E-04)、PVALB(P=6.9E-11)、RHO(P=2.4E-04)等。以基因DCT、DTL、GUCY2F和SLC4A10为例,其在实验组与对照组中的表达值的变化见图2。
2.2 差异表达基因的富集分析结果 通过Fisher检验富集分析,结合Functional categories信息(表1),发现这些基因的功能分类主要包含细胞分裂(P=4.57E-07,Count=16,7%)、细胞周期(P=9.7E-08,Count=22,9.7%)、细胞骨架(P=4.4E-04,Count=19,8.4%)、离子运输(P=4.09E-04,Count=18,7.9%)、信号识别及传导(P=4.09E-04,Count=53,23.3%)等。
根据Protein domains的数据库信息(表2),发现这些基因的蛋白结构域主要包括驱动蛋白保守位点(P=1.7E-03,Count=5,2.2%)、神经递质离子通道跨膜区域(P=2.2E-03,Count=5,2.2%)、配体结合区域(P=2.2E-03,Count=5,2.2%)、离子通道保守位点(P=2.3E-03,Count=5,2.2%)、r-氨基丁酸A受体(P=2.6E-03,Count=4,1.8%)、丝氨酸/苏氨酸蛋白激酶活性中心(P=0.01,Count=11,4.8%)等。
2.3 差异表达基因的聚类分析结果 通过Functional Annotation Clustering对差异表达基因进行聚类分析,设置Kappa Similarity Threshold为0.5且EASE为0.001时,差异表达基因的功能共分为8类。第一类为信号识别及传导(Enrichment Score=7.3),第二类为细胞周期(Enrichment Score=6.1),第三类为细胞组成(Enrichment Score=5),第四类为细胞骨架(Enrichment Score=4.6),第五类为信号转导(Enrichment Score=4.4),第六类为光传导(Enrichment Score=4),第七类为离子运输(Enrichment Score=3.6),第八类为细胞分化负调控(Enrichment Score=3.3)。
3 讨论
通过对芯片数据进行log2转换及标准化处理并运用Benjamini&Hochberg算法对样本P值进行多重检验FDR校正后,共找到265个差异表达基因。通过DAVID基因功能注释和功能分类对这些基因进行生物学和统计学分析,发现这些基因可能涉及到的生物学过程有:物质转运、细胞骨架结构、细胞周期、信号识别及传导、细胞分化调控等,与癌症相关的已知通路大体吻合。下面就这5方面对黑色素瘤的差异表达基因做一详细分析。

图1 黑色素瘤中差异表达较显著的前20个基因
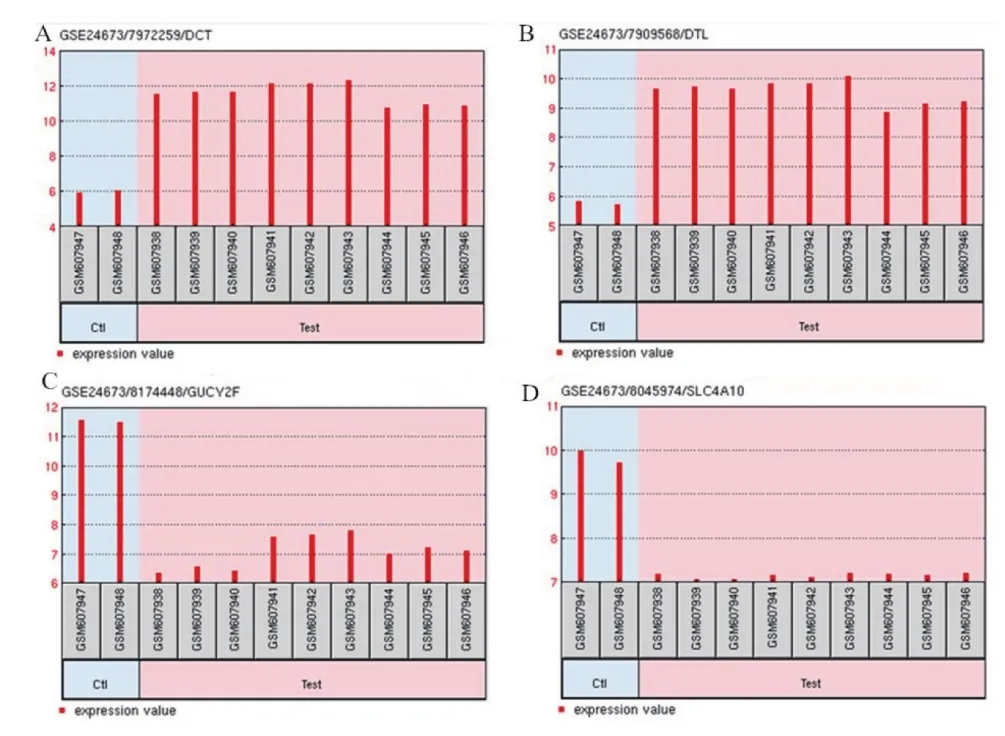
图2 实验组与对照组中基因DCT(A)、DTL(B)、GUCY2F(C)和SLC4A10(D)的表达值的变化
在细胞物质运输过程中,编码离子通道亚单位的基因发生突变或表达异常或体内出现针对通道的病理性内源性物质时,通道的功能将出现不同程度的削弱或增强,从而导致机体整体生理功能的紊乱[12,13]。SLC38A5、SLC24A1、SLC4A10、GABRG1、KCNT2、KCNJ13、HCN1、TRPM1、CNGB1、CACNG4、SCN7A等基因在HCO3–、Cl–、K+、Ca2+、Na+等离子的跨膜运输中起着关键的转运调控作用。其中,是机体最重要的pH缓冲体系的组成部分,它可以加速CO2的清除,调节细胞和整个机体的pH,调控液体的流动以及酸碱分泌等[14]。SLC4家族(solute carrier 4 family)的钠离子偶联碳酸氢根转运蛋白NCBT(Nacoupled HCO3–transporter)在细胞pH调控以及的跨上皮细胞转运中具有重要作用。Parker[15]曾研究报道,由SLC4A10基因编码的NBCn2是一种电中性的钠离子偶联的碳酸氢根转运体,在中枢神经系统中表达极为丰富,对于神经元的pH调控和脑脊液的产生等都具有重要的生理学作用。而NBCn2功能失常与癫痫、智障等疾病的发生密切相关[16]。本研究中,SLC4A10在黑色素肿瘤细胞中低表达,说明HCO3–在黑色素细胞的转运过程受到了阻碍,造成细胞生长环境失调,对细胞的肿瘤化有一定影响。

表1 差异表达基因的功能分类
微管是细胞骨架结构的主要组成部分,由α、β-微管蛋白异二聚体组成。微管在细胞分裂中具有极其重要的作用,现已成为抗肿瘤药物研究的重要靶点之一。此外,α、β-微管蛋白表达异常与细胞的癌变密切相关。Giarnieri等[17]研究发现,α、β-微管蛋白的表达与直肠癌的发生发展有关。应荣彪等[18]在乳腺非典型增生及导管内癌中发现α、β-微管蛋白均出现过度表达。本研究中,与此相关的基因KIFC1、KIF4A、PRC1、NEK2、TTK等均呈上调表达,推测这些基因的异常表达促进了细胞恶变和癌变的进展。
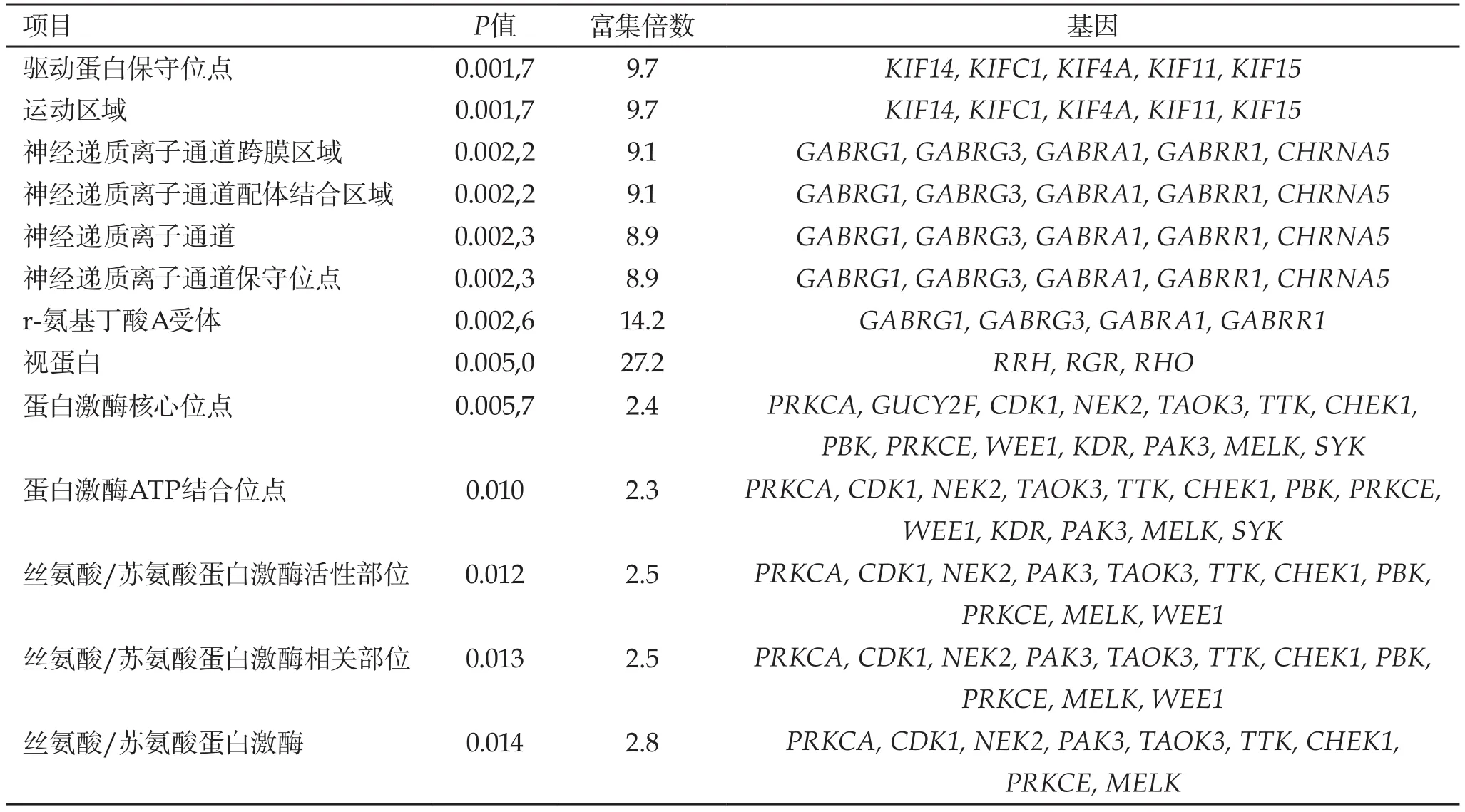
表2 差异表达基因的蛋白质结构域
细胞周期调控异常是肿瘤发生的关键。TOP2A、KIF家族(KIF14、KIF17等)、WEE1、MCM6等基因在细胞有丝分裂、减数分裂和胞质分裂过程中起重要作用。TOP2A编码的拓扑异构酶II能够在细胞的有丝分裂和减数分裂期间正确分离子染色体[19],若表达异常,可致细胞分裂紊乱。也有报道称有丝分裂驱动蛋白KIF14过表达与一系列人类癌症发生、发展密切相关。杨涛等[20]在研究肝细胞肝癌组织中KIF14表达与临床变量的关系时发现,KIF14过表达不利于手术预后,因此KIF14可作为手术预后评估的重要指标及潜在治疗靶点。WEE1基因编码产物是一种酪氨酸激酶的核蛋白,属于蛋白激酶中丝氨酸/苏氨酸家族的一员,是调控细胞周期G2期阻滞的关键元件,参与细胞有丝分裂前期的DNA复制及损伤修复过程[21]。MCM6(minichromosome maintenance protein 6)是微小染色体维持蛋白MCM家族的一员,存在于所有的真核细胞中,是DNA复制和延伸的关键蛋白,被认为是特异性细胞增殖相关因子[22,23]。本研究中,WEE1基因在黑色素肿瘤细胞中呈上调表达,说明其参与的DNA的修复异常与肿瘤的发生密切相关,可作为诊断早期黑色素瘤的候选基因,并为黑色素瘤的靶向分子治疗提供生物学依据。MCM6在黑色素瘤组织中高表达,说明其参与的DNA的复制异常与肿瘤的发生相关,可作为预防、诊断黑色素瘤的一个标记物。
信号转导异常在肿瘤形成及发展的各个阶段都有十分重要的意义。参与肿瘤生长调控的信号转导的基因有GUCY2F、GNAT1、ABLIM1、ABCA8等,在黑色素瘤组织中的表达均出现了不同程度的下调。Wood等[24]发现基因GUCY2F、EPHA3和NTRK3的低表达与乳腺癌、肺癌和胰腺癌组织细胞的恶性增殖有关。GUCY2F编码的鸟苷酸环化酶 (soluble guanylate cyclase,sGC)作为NO受体催化细胞内的GTP转化为cGMP,从而为蛋白激酶、磷酸二酯酶以及离子通道的调节提供第二信使。一旦NO信号转导通路异常,将会导致多种疾病的发生,如多种心血管疾病(如肺动脉高血压、心力衰竭、动脉粥样硬化和再狭窄等)及神经退行性疾病等。表明GUCY2F可作为黑色素瘤的诊断标志和治疗干预的靶位。
黑色素瘤特异性抗原基因(preferentially expressed antigen of melanoma,PRAME)的表达主要局限于肿瘤细胞,抑制视黄酸信号受体的活性。视黄酸与受体结合之后,起始与细胞增殖抑制、细胞分化、细胞凋亡有关的基因的转录。据Gudas等[25]报道,PRAME可调控干细胞的分化。此外,Epping等[26,27]研究发现,通过抑制PRAME对视黄酸信号受体的活性,将导致细胞的癌变及肿瘤的发展。本研究中,PRAME过表达异常显著(logFC=4.19),鉴于PRAME在肿瘤中的高水平表达,分析这种蛋白质在这些病变中的预后作用将是一项有意义的工作。
综上所述,本研究表明利用生物信息学的方法能有效地分析基因芯片数据,从而高效、大规模地获取生物内在信息;揭示黑色素瘤的发生是由于多种基因表达异常所致,为确定黑色素瘤的早期诊断标志与新治疗靶位提供了新的思路。
1 李凤鸣.眼科全书.北京: 人民卫生出版社,1996: 2381-2390.
2 吴中耀.现代眼肿瘤眼眶病学.北京: 人民军医出版社,2002:251-272.
3 Egan KM,Seddon JM,Glynn RJ,et al.Epidemlologic aspects of uvel melanoma.Surv Ophthalmol,1988,32(4):239-251.
4 Van Raamsdonk CD,Griewank KG,Crosby MB,et al.Mutations in GNA11 in uveal melanoma.N Engl J Med,2010,363(23): 2191-2199.
5 All-Ericsson C,Girnita L,Muller-Brunotte A,et al.C-Kitdependent growth of uveal melanoma cells: a potential therapeutic target? Invest Ophthalmol Vis Sci,2004,45(7):2075-2082.
6 Ye M,Hu D,Tu L,et al.Involvement of P13K/Akt signaling pathway in hepatocyte growth factor-induced migration of uveal melanoma cells.Invest Ophthalmol Vis Sci,2008,49(2): 497-504.
7 张旭,顼晓琳,李彬,等.骨桥蛋白在不同侵袭转移潜能葡萄膜黑色素瘤中的表达及意义.中华实验眼科杂志,2012,30(3):199-203.
8 Tanya B,Stephen EW,Pierre L,et al.NCBI GEO: archive for functional genomics data sets-update.Nucleic Acids Res,2013,41(D1): D991-D995.
9 Smyth G.Linear models and empirical Bayes methods for assessing differential expression in microarray experiments.Stat Appl Genet Mol Biol,2004,3: Article3.
10 Benjamini Y,Hochberg Y.Controlling the false discovery rate - a practical and powerful approach to multiple testing.J R Stat Soc B Met,1995,57(1): 289-300.
11 Dennis G Jr,Sherman BT,Hosack DA,et al.DAVID:Database for annotation,visualization,and integrated discovery.Genome Biol,2003,4(5): P3.
12 Chahine M,Chatelier A,Babich O,et al.Voltage-gated sodium channels in neurological disorders.CNS Neurol Disord Drug Targets,2008,7(2): 144-158.
13 Lossin C.A catalog of SCN1A variants.Brain Dev,2009,31(2): 114-130.
14 Romero MF,Fulton CM,Boron WF.The SLC4 family oftransporters.Pflugers Arch,2004,447(5): 495-509.
15 Parker MD,Musa-Aziz R,Rojas JD,et al.Characterization of human SLC4A10 as an electroneutral Na/HCO3cotransporter (NBCn2) with Cl-self-exchange activity.J Biol Chem,2008,283(19): 12777-12788.
16 Jacobs S,Ruusuvuori E,Sipila ST,et al.Mice with targeted Slc4a10 gene disruption have small brain ventricles and show reduced neuronal excitability.Proc Natl Acad Sci U S A,2008,105(1): 311-316.
17 Giarnieri E,De Francesco G,Carico E,et al.Alpha- and beta-tubulin expression in rectal cancer development.Anticancer Res,2005,25(5): 3237-3241.
18 应荣彪,冯俊,李建军,等.α、β-微管蛋白在乳腺癌变不同阶段的表达及意义.中国癌症杂志,2011,21(8): 595-598.
19 Sakaguchi A,Kikuchi A.Functional compatibility between isoform alpha and beta of type II DNA topoisomerase.J Cell Sci,2004,117(Pt 7): 1047-1054.
20 杨涛,孙轶飞,王立伟,等.KIF14过表达与肝细胞肝癌手术预后密切相关.河北医科大学学报,2014,35(3): 263-265.
21 Cozzi M,Giorgi F,Marcelli E,et al.Antitumor activity of new pyrazolo[3,4-d] pyrimidine SRC kinase inhibitors in Burkitt lymphoma cell lines and its enhancement by WEE1 inhibition.Cell Cycle,2012,11(5): 1029-1039.
22 Chong JP,Mahbubani HM,Khoo CY,et al.Purification of an MCM-containing complex as a component of the DNA replication licensing system.Nature,1995,375(6530):418-421.
23 Labib K,Tercero JA,Diffley JF,et al.Uninterrupted MCM2-7 function required for DNA replication fork progression.Science,2000,288(5471): 1643-1647.
24 Wood LD,Calhoun ES,Silliman N,et al.Somatic mutations of GUCY2F,EPHA3,and NTRK3 in human cancers.Hum Mutat,2006,27(10): 1060-1061.
25 Gudas LJ,Wagner JA.Retinoids regulate stem cell differentiation.J Cell Physiol,2011,226(6): 322-330.
26 Epping MT,Wang L,Edel MJ,et al.The human tumor antigen PRAME is a dominant repressor of retinoic acid receptor signaling.Cell,2005,122(6): 835-847.
27 Epping MT,Bernards R.A causal role for the human tumor antigen preferentially expressed antigen of melanoma in cancer.Cancer Res,2006,66(22):10639-10642.
Enrichment analysis of uveal melanoma-related differential expression genes
Xiaoyu SUN,Chunhua YANG Department of Basic Medicine,Ji'nan Evidence Based Medical Science Research Center of Shandong,Ji'nan 250022,China
ObjectiveTo better understand the molecular pathogenesis of uveal melanoma,and provide novel means for clinical diagnosis and treatment of this malignancy.MethodsThe gene chip data of uveal melanoma were obtained from GEO database and statistically analyzed after log2transformation and normalization to identify the differential expression genes related to uveal melanoma,follewed by bioinformatics analysis through functional annotation and functional clustering of DAVID.ResultsTwo hundred and sixty-five differentially expressed genes were identified in uveal melanoma samples,including 95 up-regulated and 170 downregulated genes.These genes were associated with the cellular transportation,cell skeletons,cell cycles,signal recognition and transduction and nervous system regulation.ConclusionBioinformatics analysis can contribute to analyzing the gene chip data effectively.The pathogenesis of uveal melanoma involves abnormal expression of multiple genes,and these data may benefit further investigations of the early diagnosis and treatment of the malignancy.
Uveal melanoma; Differential expression; Gene chip; GEO; Bioinformatics
