香果树(Emmenopterys henryi)种群种子雨、种子库及实生苗数量的海拔梯度变化
2017-11-10郭连金曹昊玮徐卫红田玉清肖志鹏
郭连金 曹昊玮 徐卫红 田玉清,2 肖志鹏
(1.上饶师范学院,上饶 334000; 2.中国科学院昆明植物研究所,昆明 650204)
香果树(Emmenopteryshenryi)种群种子雨、种子库及实生苗数量的海拔梯度变化
郭连金1曹昊玮1徐卫红1田玉清1,2肖志鹏1
(1.上饶师范学院,上饶 334000;2.中国科学院昆明植物研究所,昆明 650204)
香果树为我国Ⅱ级野生保护植物,由于原生境中香果树种群的数量迅速减少,目前已濒临灭绝,种群亟待恢复。研究了武夷山不同海拔的香果树种群种子雨、种子库及其幼苗存活现状,目的是阐明不同海拔地区的香果树种群自然更新的动态过程,寻找其更新脆弱的环节及其影响因素,为香果树自然种群的恢复提供理论依据。以分布于不同海拔高度(819、980、1 140及1 301 m)的香果树母树为中心,在其周围(东、南、西、北四个方位)布设种子雨收集框和土壤种子库样方,连续观测种子雨和种子库的动态,并对不同地表覆盖物中种子萌发及幼苗数量进行观测和记录。结果表明,武夷山香果树种群种子雨持续时间近2个月,尤其是11月底至12月中旬为种子雨高峰期;随着海拔的上升,香果树种子总密度以及虫蛀种子密度均显著降低,但千粒重以及饱满种子密度则有所增加;香果树土壤种子库为瞬时种子库,其水平分布以母树南侧和西侧种子密度较大,母树北侧种子密度最小。香果树种子的垂直分布主要集中于枯落物和苔藓层;3月时香果树土壤种子库中存留的饱满种子仅占种子散布后1.80%,而1年实生苗存活率为6.18%,土壤种子库的损耗是其种群自然更新困难的主要原因;不同地表覆盖物对香果树幼苗的存活产生显著影响,枯落物及苔藓层的幼苗死亡率显著高于土壤表面。建议清理林内枯落物及苔藓,增加林内裸露土壤的面积,减小地上草本的盖度,以减少种子霉烂、虫蛀的几率,增加香果树实生苗胚根与土壤接触的几率以及改善光照条件,促进香果树自然更新。
种子雨;土壤种子库;种子特征;种子萌发;幼苗更新
种子是植物重要的繁殖器官之一,是植物种群生活史的起点和终点,它靠自身或果实的重力以及外界力量从母体上散布到地表的过程,被称为种子雨[1]。种子雨散布的时空格局对其萌发、幼苗的存活以及种群的更新具有决定性的影响,对种群的适应性、生活史特征的演化产生重要作用[2]。植物的种子落地后,存在于土壤上层的地被物或土壤中,其总和为土壤种子库[3],Harper[4]将土壤种子库称为潜种群阶段,土壤种子库的研究对于种群生态对策、物种进化等问题的解决具有重要的学术价值,对其组成、数量以及分布的研究有助于对植物更新的了解[5~6]。近年来,已有不少国内外学者对不同植物物种的种子雨、土壤种子库以及成苗定居等方面进行了大量的研究工作[7~10]。岳红娟[11]研究了南方红豆杉(Taxuschinensisvar.mairei)的种子雨和土壤种子库,发现其种子雨持续3个多月,种子主要落在树冠内,土壤中大量种子被捕食,导致其更新困难。花楸(Sorbuspohuashanensis)种子雨则以果实散落的形式到达地表,96.1%果实分布于母树2 m范围内,土壤含水量增加可降低其幼苗的死亡率[12]。意大利槭亚种(Aceropalussubsp.granatense)土壤种子库中种子大部分已腐烂而丧失生活力,影响其自然更新[13]。尽管国内外学者对植物的种子雨、土壤种子库和幼苗进行了不少研究,但对于珍稀濒危植物相关方面的研究较少。珍稀濒危植物的种子雨、种子库、幼苗的定居是其生活史中的关键阶段,对其种群的更新尤为重要,研究珍稀濒危植物的种子雨、种子库以及幼苗的形成,对揭示其的致濒机制和促进其恢复有重要意义。
香果树(Emmenopteryshenryi)属茜草科(Rubiaceae),是第四纪冰川幸存孑遗植物之一,为中国特有单种属植物,是研究茜草科系统发育、形态演化及中国植物地理区系的重要材料,为国家Ⅱ级重点保护植物和林业部公布的国家珍贵树种[14]。近年来,由于香果树种群数量的急剧减少和我国对生物多样性保护的重视,从而引起了我国一些学者对香果树的关注。香果树主要分布于中国长江以南中海拔山区的常绿落叶林中,武夷山是其主要分布之一。武夷山香果树种群结构不完整,幼苗严重不足,种群呈衰退性趋势[15~16]。香果树为圆锥花序,无限花序之一,开花时间不整齐,具有花多果少的特征[17~18]。由于香果树每个果实具有200~800粒种子,故其种子数量相对较多。香果树种子微小,千粒重仅0.32~0.60 g[19],且种子寿命较短,室温下仅能维持10个月[20]。香果树种子萌发需光,属于光敏感种子,20℃为种子萌发的最佳温度[21~22],室内试验研究发现其萌发率在13%~20%[23]。截止目前,对香果树的研究主要集中于种群结构、种子特征及室内单一因素对其种子萌发的影响等方面,而原生境中环境复杂,对于其原生境中香果树的种子雨、土壤种子库分布格局、种子萌发以及幼苗数量变化等未见报道,而这正是该种群自然更新的关键。本文以武夷山香果树为研究对象,主要探讨(1)海拔对香果树种子雨的时空格局及其种子特征的影响;(2)不同海拔的香果树土壤种子库分布格局及种子质量的变化;(3)林下香果树实生苗的数量变化动态,旨在阐明香果树种群自然更新的动态过程、寻找其更新脆弱环节及影响因素,为该种群的恢复提供理论依据。
1 研究地自然概况
武夷山国家级自然保护区(27°33′~27°54′N,117°27′~117°51′E),是世界同纬度现存面积最大,保存最完整的中亚热带森林生态系统之一。区内自然资源丰富,保存有完好的地带性常绿阔叶林群落,是东南亚大陆生物多样性最丰富的地区,也是具有全球生物多样性保护意义的地区,本区气候属于典型的亚热带季风气候,平均气温在12~13℃,年降水量1 480~2 150 mm,年蒸发量1 492 mm左右,相对湿度78~84%,无霜期253~272 d。土壤类型主要有红壤、黄红壤、黄壤和山地草甸土[18]。香果树在研究地主要星散分布于沟谷水溪旁的阔叶林中,其伴生种主要有:银鹊树(Tapisciasinensis)、杉木(Cunninghamialanceolata)、红脉钓樟(Linderarubronervia)、盐肤木(Rhuschinensis)、野鸭椿(Euscaphisjaponica)、毛竹(Phyllostachysheterocyclacv.pubescens)、亮叶红淡(Adinandranitida)、伞形绣球(Hydrangeaangustipetala)、一年蓬(Erigeronannuus),兔儿伞(Syneilesisaconitifolia)、黄堇(Corydalispallida)、菝葜(Smilaxchina)、糯米团(Gonostegiahirta)等。
2 研究方法
2.1 样地设置及生殖构件特征统计
在武夷山国家级自然保护区选择典型的4个海拔梯度的香果树种群作为研究对象,分别设置4个面积为100 m×1 000 m的样地进行群落生态学调查(表1)。调查内容包括:①生境:海拔,坡向,坡位等,其中土壤含水量采用便携式土壤水分速测仪(TRIME-TD RZ)进行测定,用照度计(ZDH-10)测定林内地面光照,干湿球温度计(DMH2)通风干湿度温度计记录林内的温湿度(选择前两天无雨天气,林内光照、温湿度等的测定于每日10:00、13:00、15:00时进行);②群落学特征:树种组成、高度、盖度等;③对样地内香果树母株进行调查,记录其数量、胸径、树高等;④母树选择:2014年10月2日(种子雨前),每样地选择并标记5株有代表性的母树(表2),为避免香果树母树间种子雨的影响,本研究所标记的母树生存环境相对开阔,其周围200 m内没有其他母树开花、结果。

表1 香果树种群四生境概况
注:林冠层光照、温湿度、土壤含水量等的测定于2014年11月进行,每样地连续观测3日,于每日10:00、13:00、15:00时进行测定;草本层盖度为2015年5~11月每月一次调查的平均值。
Note:Light intensity on the ground in forest,temperature,humidity and soil water content were determined on June 20-22 2010,at 10:00,13:00 and 15:00 each day.Average values of herbaceous layer coverage were investigated monthly May-Nov,2015.

表2 香果树形态指标
2.2 种子雨收集
2014年10月底,在标记母树的周围按东、西、南、北4个方位,选取距离母树树干5、10、20、40和80 m均匀布设20个种子收集框,框口面积为100 cm×100 cm,框深20 cm[24]。由于香果树种子微小,本研究收集框采用网眼0.5 mm×0.5 mm的尼龙网进行收集种子,为避免小动物的破坏,收集框四周用4根1.3 m高的木棍撑起,使框底距地面1 m左右,每隔3 d观察并记录收集框内种子的数量,直至连续7 d种子数量不变结束观察。待香果树母树种子雨结束后,将收集框内种子按饱满、干瘪、空粒以及虫蛀分别进行记录。
2.3 种子库收集
于2015年1月20~25日,在与收集框相邻区域设置9个50 cm×50 cm的土壤种子库样方,分别用于1、3和5月3次对香果树种子进行观测。在每个土壤种子库样方中分别以枯落物和苔藓层、0~1和1~5 cm土壤层进行取样,每个样地每次共观测900份样品。采用网筛法分离土壤种子库中的香果树种子,并记录不同层筛选出的饱满、干瘪、虫蛀(种子的胚被啃食作为标准)以及霉烂种子数。
2.4 香果树实生苗的数量调查
由于单株香果树土壤种子库所形成的幼苗数量过少,无法进行统计,本研究以所标记的香果树母树为对象,5月确定萌发种子所处的地被物及土壤层次,自5月起,每隔2个月对所有实生苗观测一次,记录幼苗的数量、所处的方位及距母树的距离,并按东、南、西、北、距母树不同距离以及地被物和土壤层进行分类,以对香果树幼苗的数量动态进行分析。
2.5 数据处理
利用EXCEL2013对数据进行整理,计算平均数及标准差,采用SPSS19.0软件对香果树种群的种子雨、土壤种子库中种子的密度以及不同条件下幼苗的数量进行单因素方差分析,并对各因素水平间的数据进行LSD多重比较,显著水平选择α=0.05。文中图均由软件Origin 9.0制作。
3 结果与分析
3.1 香果树种子雨特征
3.1.1 香果树种子特征及组成
武夷山香果树种子的千粒重约0.57 g,种子长、宽和厚度范围分别为0.55~12.3、0.1~4.0和0.07~0.48 mm,种子饱满率仅为35%,不饱满的种子达到了50%,其余种子被虫蛀。不同海拔的香果树种群种子特征及组成存在一定差异,其中除A种群中种子平均长度显著小于其他种群外,海拔对其他3个种群种子形态指标影响不显著。不同种群的香果树种子密度随着海拔的升高呈显著降低的趋势,但千粒重和饱满种子密度与之相反,随着海拔的升高呈显著增加趋势。随着海拔的升高,香果树种群的干瘪、空粒、虫蛀以及总种子密度均呈下降趋势,其中不同种群香果树的虫蛀种子和总种子密度达到显著水平。
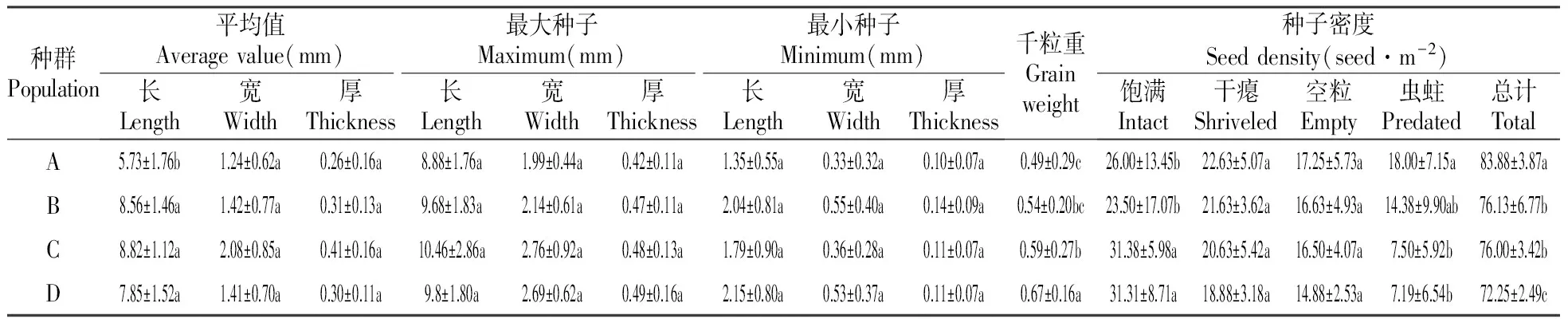
表3 不同海拔的香果树种群种子质量
注:同列数据标注不同小写字母代表数据之间存在显著差异;相同字母表示差异不显著(α=0.05)
Note:Values with different letters in the same column are significantly different and the same letters indicate no significant difference according to the LSD multiple comparison test(α=0.05).
3.1.2 种子雨散布的时间动态
武夷山香果树种群多沿沟谷、溪流分布,且主要存在于阳坡和半阳坡(表1),结实母树多集中于光照充足的林缘,种子雨持续时间近2个月。种子10月底开始成熟,11月初蒴果开裂种子散落至11月底为种子雨起始期,11月底至12月中旬种子雨强度最大,为种子雨高峰期,12月下旬为其末期。不同种群香果树种子雨高峰期其种子下落量均达到种子雨总量的70%以上,种子雨起始期和末期下落量较少(图1)。海拔对香果树种群种子雨的高峰期存在影响,随着海拔的上升,香果树种子雨高峰期逐渐推迟,低海拔高峰期前上升慢,但下降较快,由最大下落量当天仅需5 d左右就可进入种子雨末期,而高海拔由最大下落量日至末期需要10 d左右,种子雨末期相应向后推迟。
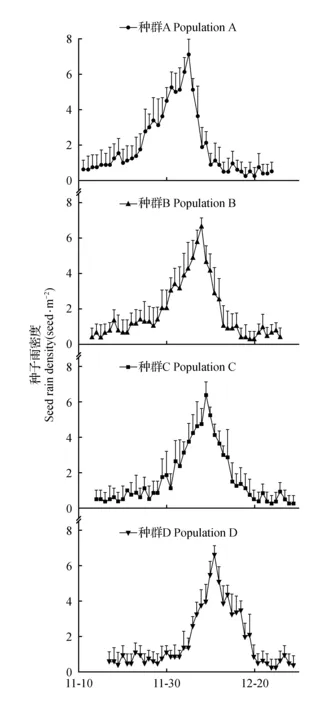
图1 不同海拔香果树种群种子雨动态变化Fig.1 Changes of the seed rain of E.henryi in different populations
3.1.3 种子雨的强度分布
由图2知,武夷山香果树种子可飘至距母树80 m远的地方,其种子雨大部分落在母树40 m范围内,40 m外仅有少量分布。不同种群的香果树种子雨强度分布格局存在显著差异(P<0.05),其中位于中海拔的种群B和种群C差异不显著。随着海拔的上升,距母树相同距离处的种子雨强度基本呈下降趋势。种群A中,随着距母树的距离的增加,相邻距离的香果树种子雨强度均呈显著下降趋势,而其余海拔在距母树树干5和10 m处种子雨强度差异不显著。
3.2 香果树土壤种子库时空分布格局
3.2.1 土壤种子库的水平分布特性
经单因素方差分析知,不同种群的香果树土壤种子库中种子密度之间存在显著差异(P<0.05),且不同类型的种子密度在母树周围的分布也存在显著差异(图3),其中母树南侧最大,西侧次之,北侧最小;同一方位种子密度以饱满种子最小,虫蛀、霉烂种子较大。1月下旬香果树土壤种子库饱满种子的平均密度已由12月底的27.05降至4.74 粒·m-2,且均显著小于同期其它各类种子密度,其最大饱满种子的密度主要分布于母树南方和西方;而虫蛀种子平均密度也由12月底11.77上升至24.65 粒·m-2,霉烂种子由0增至23.90 粒·m-2。虫蛀和霉烂种子在母树的南方密度较大、北方最小,两者占总种子数量的78.25%。
图2 香果树母树周围种子雨强度格局 不同字母间表示差异显著(P<0.05)Fig.2 Seed rain intensity around the mother tree in different population Data marked with the different small letters indicate a significant difference at 0.05 levels, respectively.

图3 不同海拔的香果树种群土壤种子库水平分布 同一类型的香果树的种子密度值上标不同字母代表数据之间存在显著差异,否则无差异。Fig.3 Horizontal distribution of soil seed bank of E.henryi in different population Density values of the same type of E.henryi seeds with different letters are significantly different,and values with the same letters.

图4 不同种群的香果树土壤种子库垂直分布 土壤种子库同一类型的香果树种子密度上标不同字母表示数据间显著差异。Fig.4 Horizontal distribution and vertical distribution of soil seed bank of E.henryi in different populations Density values with different letters in the same type of E.henryi seeds are significantly different(P<0.05).
3.2.2 土壤种子库的垂直分布特征
由图4可知,香果树土壤种子库中种子主要存留于枯落物和苔藓层中,其种子密度最大,平均为37.10粒·m-2,约占总种子数量的70%,其次为0~1 cm土壤表层,密度为14.44粒·m-2,约为总种子数量的20%,其余位于土壤1~5 cm,大于5 cm土层没有发现香果树种子。对不同种群的香果树土壤种子库中种子密度进行单因素方差分析,表明四者存在显著差异(P<0.05),随着海拔的上升,香果树土壤种子库中不同层中种子总密度呈显著下降趋势,但其饱满种子密度则出现种群B和C较小,种群A和D较大的现象。
3.2.3 土壤种子库时间变化动态
对香果树土壤种子库进行3次调查,结果显示随着海拔的上升,1月香果树总种子密度呈下降趋势,而3和5月呈现出上升趋势(图5);香果树饱满和干瘪种子比率在1和3月随着海拔的升高有小幅波动,但5月基本呈上升趋势;不同种群的香果树土壤种子库中虫蛀种子比率差异不显著,总体来看,3月调查的虫蛀种子比率高于1月,与5月差异不显著(P>0.05);不同种群的香果树种子库中发霉种子百分比在1和3月差异不显著,而5月D种群土壤种子库中霉烂种子的百分比显著低于A种群。
3.3 香果树实生苗分布特征
香果树原生境中种子萌发时间较林内其他物种晚,每年4月种子开始出现萌动,5月幼苗开始大量出现,6月及以后野外未发现种子萌发现象。香果树母树周围的1年实生苗数量变化见表4,由表4可知,随着海拔的升高,11月当年生实生苗的成活数量呈增加趋势。对不同种群的香果树实生苗的数量进行LSD多重比较发现,不同月份间存在显著差异,实生苗在7月以前死亡率极高,可达70%,而后逐渐下降,到11月降至30%以下。其中枯落物和苔藓层中的实生苗死亡率最高,可达80%以上,土壤0~1 cm层中实生苗最低,为48%,而土壤1~5 cm层中未发现有实生苗出现。母树不同方位的实生苗死亡率差异较小,其中母树南侧的实生苗死亡率高于其他。香果树幼苗随着距母树的距离增大,其总死亡率呈上升趋势。
4 讨论
4.1 香果树种子特征及种子雨散布
种子是植物生长周期中的特殊或重要阶段,它对于种群个体的繁殖、种群的扩展、种群遭破坏后的恢复和物种抵抗不良环境有着重要意义[25]。武夷山香果树种子具翅,千粒重在0.49~0.67 g,种子长、宽和厚度平均值依次约为7.74、1.54和0.32 mm。随着海拔的上升,其千粒重、种子饱满密度呈显著增加趋势,干瘪、空粒以及虫蛀种子密度呈减小趋势,这表明海拔对该物种的种子质量存在显著影响。

图5 香果树土壤种子库种子质量的变化 不同月份香果树种子比率或密度值上标不同字母表示数据间差异显著。Fig.5 Seed quality changes of soil seed bank in E.henryi forests Seed density or ratio values with different letters in the same E.henryi population are significantly different(P<0.05).

种群Population月份Month枯落物/苔藓Litterandmoss土壤Soil(0~1cm)东East南South西West北North距母树树干距离Distancefrommothertreetrunk(m)0~55~1010~2020~4040~80A5月May209.0±37.1a16.0±12.0a59.3±16.3a77.5±15.9a54.5±18.6a34±7.3a66.3±12.6a70.5±15.2a41.5±14.6a28.0±7.4a9.0±5.0a7月Jul.11.8±1.3b8.0±3.4b6.0±1.8b6.3±1.3b4.8±2.2b3.0±0.8b5.8±0.8b6.5±2.4b4.0±0.8b3.8±1.0b0.0±0.0b9月Sep.3.0±1.4c5.0±2.9c2.8±1.0c3.3±1.0c2.5±1.0bc1.0±0.0b1.8±0.5c2.8±1.5c2.3±0.5bc2.0±0.8bc0.0±0.0b11月Nov.1.3±1.3c4.8±2.5c1.5±06c1.8±0.5c1.3±0.5c0.8±0.5b1.5±0.6c2.8±1.5c1.0±0.0c1.0±0.0c0.0±0.0bB5月May169.5±32.9a28.3±10.0a51±19.7a59.3±17.9a58.5±24.4a29.5±5.6a55.3±6.3a62.5±16.1a36.8±10.4a25.5±9.3a7.8±6.2a7月Jul.17.8±2.2b9±4.1b6.5±2.4b8.3±2.5b8.3±3.3b4.0±0.8b6.5±0.6b9.0±2.2b4.0±1.3b3.8±1.3b2.0±0.8b9月Sep.7.3±1.0c2.8±1.5c2.5±1.0c3.0±0.8c3.0±1.2c2.3±0.5bc2.0±0.0c3.5±1.0c2.3±0.5bc2.0±0.8bc0.0±0.0b11月Nov.5.8±1.0c1.3±0.5c2.3±0.5c2.0±0.8c2.3±1.5c0.8±0.5c2.0±0.0c2.8±1.5c1.8±0.5c1.0±0.0c0.0±0.0bC5月May133.5±27.9a34.5±8.2a42±16.6a55.8±16.9a48±17.9a22.3±2.2a51.3±16.0a56.0±15.6a28.3±4.4a18.8±3.0a6±1.4a7月Jul.18.0±2.6b11±4.1b7.8±3.6b9.8±2.6b8.0±2.9b3.8±0.5b7.8±1.0b9.8±2.2b5.0±1.6b4.8±2.1b1.8±0.5b9月Sep.10.0±2.2c5.3±1.3c3.8±2.2c4.3±1.7c4.3±1.5c2.0±0.0b5.3±1.3bc5.3±1.3c3.0±0.8b1.8±1.0c0.3±0.5b11月Nov.9.0±2.2c3.0±0.8c3.5±1.7c4.0±1.4c4.0±1.4c2.0±0.0b3.3±0.5c4.8±1.5c2.5±1.0b1.5±0.6c0.0±0.0bD5月May96.5±17.1a46.5±14.2a47.0±22.0a39±15.7a36.3±15.7a20.8±3.9a40.8±9.2a46.8±11.7a25.3±7.9a19±5.4a4.8±1.0a7月Jul.24.8±4.6b16.5±4.5b13.0±5.9b11.3±4.6b10.8±4.6b6.0±1.4b11.8±3.3b13.5±2.4b7.0±1.4b6.0±1.4b3.0±0.8b9月Sep.15.0±3.7c9.5±1.3c7.8±2.8c7.3±2.9c7.3±2.9c3.5±1.3b5.5±1.3c8.5±1.3c5.0±0.8bc3.8±0.5b1.5±0.6b11月Nov.13.5±3.4c8.5±1.3c7.0±2.2c7.0±2.4c6.8±2.4c3.3±1.5b5.5±1.5c7.8±1.7c4.0±0.8c3.5±0.6b1.3±0.5b
注:同列中相同种群不同月份的香果树幼苗数量值上标不同字母表示数据间差异显著,相同字母表示数据间差异不显著。
Note: Seed density values with different letters in the sameE.henryipopulation are significantly different,and values with the same letters are nosignificantly different at α=0.05.
武夷山香果树种子雨始于11月10日左右,终于12月底,不同种群间因海拔的升高而出现种子雨高峰期推迟现象,但基本位于11月底至12月中旬。这与南方红豆杉(Taxuschinensisvar.mairei)[11]类似,与东北的长白落叶松(Larixolgensis)(10月达种子雨高峰期)以及东北的胡桃楸(Juglansmandshurica)(9月达高峰期)有差异[26~27],这主要是由于植物物种的种子雨动态变化与外界环境有关,同时也与树种自身属性有关。de Heredia[2]研究认为西班牙松(Pinuscanariensis)种子的扩散能力远高于其他松科植物。香果树果实成熟开裂后,种子在重力和风的作用下散落。母树的大小、果实的多少以及在树冠上的分布格局对种子近距离其主导作用,而远距离主要由于风的作用导致。香果树母树树高在15 m以上,冠幅约10 m左右,在离其主干5和10 m处的种子雨强度差异较小,但较远的地方,如40 m以及以外的地方种子雨强度较小,但这种散布能力也远高于南方红豆杉(树冠以外2 m)、垂序商陆(Phytolaccaamericana)(母株2 m内)[28],与水曲柳(Fraxinusmandshurica)[29]相当,这有利于香果树更新个体的发生及种群的稳定。
4.2 香果树土壤种子库分布及其种子变化
种子脱落后的其命运对种群更新具有很大作用。香果树种子成熟后,果皮开裂,种子于当年11月开始散落,土壤种子库开始形成,此时动物的搬运、取食、真菌感染、发芽(与温度、湿度、光、化感作用等因素有关)等都是影响种子库动态的因素,而所有这些因素因种子本身状况和生态环境的异质性影响程度又具差异性。本研究中香果树土壤种子库水平分布种子密度以南方最高,北方最低,垂直分布主要存在于枯落物层和苔藓层(70%),土层5 cm以下没有发现其种子存在。土壤种子库中饱满种子的密度均显著小于其它各类种子密度,1月饱满种子比率由种子雨中的36.68%降至7.81%,而虫蛀种子和霉烂种子由11.77%上升至78.25%。
孙书存[30]等研究表明一些水青冈、栎树、槭树等热带、温带植物种子肉质,易被取食和搬运,取食压力始终是土壤种子库种子损耗的首要因子。本研究中三次种子库调查发现,同一种群中香果树土壤种子库中的饱满、扁平的种子比率随着时间的推移,其数量显著下降,霉烂种子显著增加,而虫蛀种子则呈先增加后减少的趋势,5月土壤种子库中霉烂种子和虫蛀种子百分比均值依次为49.50%和38.89%,这表明香果树种子的损耗主要有霉烂和虫蛀所致。随着海拔的升高,5月霉烂种子数量呈减小趋势,这表明海拔越高,香果树种子库中种子受霉菌侵染的数量逐渐减小。这可能由于高海拔地区,其土壤温度较低、湿度较小,不利于土壤微生物的活动。
4.3 香果树林下实生苗的数量动态
Bonfil[31]研究认为种子大,淀粉、蛋白质等营养物质含量高,有利于种子发芽,且萌发幼苗抵抗不良环境的能力强,而云杉(Piceaasperata)种子较小,种子萌发生根的延伸能力有限[32]。本研究发现枯落物层和苔藓层上的香果树种子萌发后,大多胚根裸露在外面,多为耗尽营养死亡,这是由于其种子小,没有足够的营养物质使其胚根穿过地被物到达土壤层,极少数幼苗穿过地被物薄的地方,延伸到达土壤表面得以汲取营养,从而导致7月枯落物和苔藓层的香果树实生苗成活率(11.87%)低于土壤表层(35.53%),当少数实生苗胚根扎入土壤以后,由于地被物可维持局部环境相对稳定,导致其环境中的实生苗成活率升高,甚至高于土壤表层产生的实生苗(77.78%)。
香果树实生苗对光照要求严格,因光照不足,其长时间持续子叶期后而导致不能产生真叶而死亡[23]。香果树实生苗主要集中于其母树南方、距母树10 m以内的区域,距离越大,实生苗的数量越少,而7月香果树冠下实生苗存活率较低,特别是0~5 m范围内实生苗死亡率较高,其原因如前述,可能是母树下积累了大量落叶,阻碍了幼苗的进一步生长,其次为由于母树树冠的遮盖,光照条件较差,使其得不到足够的光照导致死亡。随着海拔的升高,相同月份当年实生苗的数量呈增加趋势,这可能是由于低海拔植物物种丰富,草本盖度较高(表1),香果树实生苗与其他物种竞争处于劣势,而高海拔地区则随着草本植物密度的减少,盖度降低,竞争压力随之减小,有利于其幼苗的存活。
4.4 香果树自然更新的障碍
植物能否正常生长、繁殖,关键是由其内在属性决定,稀有、濒危植物的生殖力、存活力和适应力低下是其走向衰退甚至灭绝的根本原因。张文辉[6]对10种濒危植物进行了分析,认为多数植物在由种子向幼苗转化阶段出现死亡率高或种子不能成苗的现象,成为种群衰退的关键阶段。香果树母树在现存生境中生长较好,能正常开花结果,种子散布较远,能在适宜的环境中萌发,但其土壤种子库中的种子大部分发霉或虫蛀,3月调查结果显示散布的种子中仅有1.80%的饱满种子保存下来,而实生苗萌发后仅有6.18%存活到当年的11月,由此可见,香果树种子至幼苗阶段是其自然更新的限制环节,其严重制约了香果树的自然更新。为降低种子霉烂和被捕食的危险,增加种子萌发后的成活率,需清理林下枯落物、苔藓层以及草本层,减少病原菌以及害虫的滋生环境,增加土壤表面的光照强度,另可通过收集种子,低温冷藏[20],待翌年3月播种,以增加饱满种子的数量。由于种子雨后,翌年6月野外生境中没有出现香果树种子萌发的现象,这可能预示着其寿命终结,这表明香果树土壤种子库为瞬时种子库。关于其野外条件下的香果树种子寿命的问题有待进一步研究。
致谢感谢武夷山国家自然保护区管理局科技科工程师徐自坤、谢鹏、金昌善等三位同志以及护林员詹建魁在香果树种子雨和种子库野外调查中的指导与帮助。
1.Janzen D H.Seed predation by animals[J].Annual Review of Ecology and Systematics,1971,2(1):465-492.
2.Heredia U L D,Nanos N,Garcíadelrey E,et al.High seed dispersal ability ofPinuscanariensisin stands of contrasting density inferred from genotypic data[J].Forest Systems,2015,24(1):e015.
3.Simpson R L.Ecology of soil seed bank[M].San Diego:Academic Press,1989:149-209.
4.Harper J L.Population biology of plants[M].London:Academic Press,1977:256-263.
5.Yu S L,Jiang G M.The research development of soil seed bank and several hot topics[J].Acta Phytoecologica Sinica,2003,27(4):552-560.
6.Zepeda C,Lot A,Nemiga X A,et al.Seed bank and established vegetation in the last remnants of the Mexican Central Plateau wetlands:the Lerma marshes[J].Revista De Biología Tropical,2014,62(2):455-472.
7.Bertiller M B,Carrera A L.Aboveground vegetation and perennial grass seed bank in arid rangelands disturbed by grazing[J].Rangeland Ecology & Management,2015,68(1):71-78.
8.Li W Q,Khan M A,Yamaguchi S,et al.Hormonal and environmental regulation of seed germination in salt cress(Thellungiellahalophila)[J].Plant Growth Regulation,2015,76(1):41-49.
9.Li X,Jiang D,Zhou Q,et al.Soil seed bank characteristics beneath an age sequence ofCaraganamicrophyllaShrubs in the Horqin sandy land region of northeastern China[J].Land Degradation & Development,2014,25(3):236-243.
10.Bebawi F F,Campbell S D,Mayer R J.Seed bank longevity and age to reproductive maturity ofCalotropisproceraAit. W.T.Aiton in the dry tropics of northern Queensland[J].The Rangeland Journal,2015,37(3):239-247.
11.Yue H J,Tong C,Zhu J M,et al.Seed rain and soil seed bank of endangeredTaxuschinensisvar.maireiin Fujian,China[J].Acta Ecologica Sinica,2010,30(16):4389-4400.
12.Xu J W,Shen H L,Zhang X L,et al.Sorbus pohuashanensis seed dispersal and germination and their relationships with population natural regeneration[J].Chinese Journal of Applied Ecology,2010,21(10):2536-2544.
13.Gómez-Aparicio L,Gómez J M,Zamora R.Spatiotemporal patterns of seed dispersal in a wind-dispersed Mediterranean tree(Aceropalussubsp.granatense):implications for regeneration[J].Ecography,2007,30(1):13-22.
14.Fu L G.Chinese red book of plants:rare and endangered plants[M].Beijing:Science Press,1991.
15.Guo L J.Population structure and dynamics of the endangered plantEmmenopteryshenryi[J].Journal of Wuhan Botanical Research,2009,27(5):509-514.
16.Guo L J,Li M,Lin S.Flowering phenology,reproductive module characteristics and their influencing factors of endangered plant SpeciesEmmenopteryshenryi[J].Forest Research,2015,28(6):788-796.
17.Guo L J.Spatial distribution pattern and number dynamics ofEmmenopteryshenryiseedlings endangered in China[J].Acta Bot.Boreal.-Occident.Sin.,2014,34(9):1887-1893.
18.Guo L J,Xue P P,Shao X H,et al.Growth characteristics and influencing factors ofEmmenopteryshenryiroot sprouts[J].Plant Science Journal,2015,33(2):165-175.
19.Li Z Y,Ban Q.Biological characteristics and reproductive methods ofEmmenopteryshenryi[J].Forest science and Technology,1995,(4):37-38.
20.Chen L,Zhou K.Study on the storage time ofEmmenopteryshenryiOliv.seeds[J].Journal of Huangshan University,2007,9(5):64-66.
21.Li T H,Zhou Y X,Duan X P,et al.Physiological characteristics of the dormancy and light-sensitive germination ofEmmenopteryshenryiseeds[J].Journal of Central South Forestry University,2004,24(2):82-84.
22.Kang H J,Chen Z L,Zhou Y H,et al.Seed germ ination and dynamics of seedling growth ofEmmenopteryshenryi[J].Journal of Central South University of Forestry & Technology,2011,31(1):32-37.
23.Yang K J.Primary study on conservation biology of the rare speciesEmmenopteryshenryiOliv.[D].Wuhu:Anhui Normal University,2007:27.
24.Huang H L,Zhang L,Liao C K.Seed rain,soil seed bank,and natural regeneration of naturalToonaciliatavar.pubescensforest[J].Chinese Journal of Applied Ecology,2012,23(4):972-978.
25.Yin H J,Liu Q.Seed rain and soil seed banks of Picea asperata in subalpine spruce forests,western Sichuan,China[J].Acta Phytoecologica Sinica,2005,29(1):108-115.
26.Liu Z G,Zhu J J,Yuan X L,et al.On seed rain and soil seed bank ofLarixolgensisin montane regions of eastern Liaoning Province,China[J].Acta Ecologica Sinica,2007,27(2):579-587.
27.Ma W L,Jing T,Kujansuu J,et al.The dynamics of seed rain and seed bank ofJuglansmandshuricapopulation in the Changbai Mountain.[J].Journal of Beijing Forestry University,2001,23(3):70-72.
28.Zhai S Q,Li C R,Xu J W,et al.Spatial and temporal dynamics ofPhytolaccaamericanaseed rain underRobiniapseudoacaciaforest in Lingshan Bay National Forest Park,Shandong,China[J].Acta Phytoecologica Sinica,2010,34(10):1236-1242.
29.Han Y Z,Wang Z Q.Spatial pattern of Manchurian ash seed dispersal in secondary hardwood forests[J].Acta Phytoecologica Sinica,2002,26(1):51-57.
30.Sun S C,Chen L Z.Seed demography ofQuercusliaotungensisin Dongling mountain region[J].Acta Phytoecologica Sinica,2000,24(2):215-221.
31.Bonfil C.The effects of seed size,cotyledon reserves,and herbivory on seedling survival and growth inQuercusrugosaandQ.laurina(Fagaceae)[J].American Journal of Botany,1998,85(1):79-87.
32.Aizn M A,Woodcock H.Effects of acorn size on seedling survival and growth inQuercusrubrafollowing simulated spring freeze[J].Canadian Journal of Botany,1996,74(2):308-314.
The National Natural Science Foundation of China(31360145)
introduction:GUO Lian-Jin(1976—),male,Associate Professor,mainly engaged in the research work of plant ecology and biodiversity.
date:2016-11-15
SeedRain,SoilSeedBankandQuantitativeDynamicsofSeedlingsofEmmenopteryshenryiPopulationsinDifferentAltitudeRegions
GUO Lian-Jin1CAO Hao-Wei1XU Wei-Hong1TIAN Yu-Qing1,2XIAO Zhi-Peng1
(1. Shangrao Normal University,Shanrao 334000;2.Kunming Institute of Botany,Chinese Academy of Sciences,Kunming 650204)
EmmenopteryshenryiOliv., the only species of the genus Emmenopterys belonging to the family Rubiaceae, is one of the most rare trees endemic to China. The size of the wild population ofE.henryihas been noted to be very small and to be endangered now. In Wuyishan Natural Reserve, the seed rain, seed banks and the dynamics of seedlings ofE.henryiwere observed continuously. We demonstrated the dynamic processes of natural regeneration ofE.henryipopulations to find out the most weak stages as well as their impacting factors, and ultimately to provide scientific basis for the recovery ofE.henryipopulations. FiveE.henryitrees were selected and labeled as the research objects in each plot with different altitudes. In the directions of east, west, south and north of everyE.henryitree labeled, five sampling points was arranged, and on each point one 0.5 m×0.5 m×0.2 m collection box was set to estimate the seed rain, nine 0.5 m×0.5 m mini-plots were set to study the dynamic of the soil seed bank, and the seed germination and growth dynamic of seedlings were observed in each plot. The seed rain lasted for about two months, and the maximum intensity of seed rain emerged from the late November to mid of December, which was the peak of seed rain. With the altitude increasing, both of the total seed density and predated seed density were decreased significantly, by contrast, the thousand seed weight and intact seed density were both increased. The seed bank ofE.henryipopulations was transient soil seed bank. The seed densities on the southern and eastern sides of the mother tree were more than those of other directions, and most of the seeds were distributed in litter and moss layers. In March, the proportion of intact seeds in soil seed bank was only 1.80% of all dispersed seeds and the survival rate of one-yearE.henryiseedlings was 6.18%. The loss of seeds in soil seed bank is the main reason resulting in the difficulty of the natural regeneration ofE.henryipopulation. Different ground surface types had a significant effect on the survival of the seedlings, since the mortality rate of seedlings growing in litter and moss layers was 88%, which was significantly higher than that in soil(64%). The litter and moss layers in the forest should be cleared up to release more space for soil and also reduce the coverage of the herbaceous, thus the probability of the seeds to go bad or be destroyed decreased and therefore the radicle of the seedlings could be better reach the soil. The light condition should also be improved, and therefore the natural regeneration ofE.henryipopulation could be improved.
seed rain;soil seed bank;seed characteristics;seed germination;seedling regeneration
国家自然基金(31360145)资助
郭连金(1976—),男,副教授,主要从事植物生态学和生物多样性研究。
2016-11-15
Q949.781.1
A
10.7525/j.issn.1673-5102.2017.03.008
