小兴安岭北部原始阔叶红松林红松结实气候敏感性及其种子年机制探讨
2017-11-10程春香毛子军靳世波宋国华孙鹏飞
程春香 毛子军 靳世波 宋国华 孙鹏飞 孙 涛
(1.东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040; 2.五营气象局/五营森林生态与农业气象试验研究站,伊春 153033; 3.丰林国家级自然保护区管理局,伊春 153033; 4.中国科学院沈阳应用生态研究所,沈阳 110016)
小兴安岭北部原始阔叶红松林红松结实气候敏感性及其种子年机制探讨
程春香1,2毛子军1*靳世波2宋国华3孙鹏飞2孙 涛1,4
(1.东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040;2.五营气象局/五营森林生态与农业气象试验研究站,伊春 153033;3.丰林国家级自然保护区管理局,伊春 153033;4.中国科学院沈阳应用生态研究所,沈阳 110016)
选取林分较为稳定的原始阔叶红松林作为研究地点,分析近16年的红松结实动态,研究红松(Pinuskoraiensis)结实的气候敏感性及其种子年机制。首先按资源限制理论分析了红松结实的资源限制性,再根据资源匹配(气候假说)理论和Nicholls关于产量研究的建议将红松种子产量分解为趋势产量和气候产量,利用红松气候产量与前3年的气象数据进行气候影响分析,并且结合前期的物候观测,找出了红松结实的气候敏感阶段及其敏感的气象指标。结果显示:(1)红松当年种子产量与去年和前年产量和呈显著负相关,二者呈指数关系,表明了红松结实中资源限制机制的存在。并且资源限制可能是球果成熟对同年其它生殖发育的单向自我营养限制关系。(2)去年与前年6月平均气温的差值(ΔT6)对当年红松种子产量具有很好的预测性。(3)花原基形成期是红松结实最为关键的气候敏感期,红松气候产量与此阶段气温和日照呈极显著负相关,与降水呈极显著正相关。(4)部分年份异常高温已经超出了花原基形成的最适温度范围,导致了红松产量的下降。(5)红松结实在生殖发育期内不同阶段都对气候变化表现敏感,红松种子年现象受到了资源限制机制和资源匹配机制的共同作用。本研究结果预示了温度升高,尤其是在气温升高过快的地区,红松结实对气候变暖具有一定的脆弱性并可能成为未来红松天然更新的重要瓶颈。
红松;小兴安岭;种子年;气候产量;气候敏感性
红松(Pinuskoraiensis)是东北亚地区的代表植物种和温带针阔混交林的建群种,为国家二级重点保护野生植物[1],主要分布于中国东北的小兴安岭和长白山区,对环境变化具有较高的敏感性[2],其种子具有极高的营养和医药价值[3]。红松种子产量年际波动较大,种子产量的多寡将直接影响种子经济效益和经营策略,甚至影响红松的天然更新。植物大量生产种子的年份常被称为植物的大年结实,即种子年现象[4]。通常,影响植物开花结实的原因分为内因与外因,内因是植物本身的资源限制,外因则是外界环境条件的影响[5]。对于这两种情况,学者们提出了不同的假说,其中较为普遍接受的有资源限制假说(resource switching,RS)和资源匹配假说(resource matching,RM)。资源限制假说认为[6~8]种子年现象是大年结实树种的内部自我资源限制。种子年的出现,限制了之后年份的产量,原因可能是种子年消耗了大量资源(主要为碳水化合物、N和P等矿质养分),林木内资源存储迅速减少,影响了之后年份的结实。资源匹配假说则认为[9~10],种子生产的年际变化是由重要的环境变量所决定的,主要包括气候因子,因此也称气候假说。红松的生殖周期为3年,其结实也必然受到内外因素的共同影响[8,11~12],一方面是受生殖过程中资源可利用性及其分配的调节,另一方面是气象条件对红松从花原基形成到球果成熟等生殖发育阶段有着重要影响,可使立地条件不同及异龄性很强的红松林形成一致的丰歉周期[2,8]。然而迄今有关影响红松结实的机理研究尚未见详细报道。
随着气候变暖问题受到国际关注,气候变化的敏感性和脆弱性逐渐被人们所认识和重视。敏感性是指系统受到与气候有关的刺激因素影响的程度,包括有利和不利影响。脆弱性则是指气候变化对系统造成的不利影响的程度[13]。气候变暖不同程度地影响了各种针叶林生态系统和裸子植物的生殖生长与球果和种子产量[11,14],也必将影响红松种子产量。因此进行红松结实的气候敏感性和脆弱性研究,准确预测种子产量,加强红松种子资源管理,调控松果采摘,对红松林群落的发展、稳定和更新以及可持续管理具有重要意义。
综上我们提出假设:(1)红松结实大小年的规律符合资源限制假说,种子产量存在自我限制性。(2)红松结实受资源限制与资源匹配机制的共同影响。(3)红松种子产量与其生殖发育各阶段的气象因子均密切相关[2],而花原基形成和幼果越冬时期的气候条件可能是影响红松结实的关键因子。鉴于此,(1)根据资源限制假说,我们进行红松种子产量年际间相关分析,以探讨红松资源限制性,并找出红松结实的自我限制规律。(2)进行红松气候产量分解,消除产量中资源限制因素的影响,结合红松生殖发育阶段开展红松气候敏感性和脆弱性研究,分析红松结实的气候敏感阶段及其敏感的气象指标。(3)找出影响红松种子产量的关键因子。本文研究结果可为揭示红松种子年可能的机理、探明红松结实对气候变化的敏感性和脆弱性、准确预测未来种子产量、加强红松种子资源管理及红松坚果林定向培育经营技术的合理制定提供科学依据。
1 研究地概况与研究方法
1.1 研究地概况及数据资料来源
1.1.1 研究地概况
本文研究地点为丰林国家级自然保护区,地理坐标为128°58′~129°15′E,48°02′~48°12′N。保护区位于黑龙江省小兴安岭南坡北段五营林区境内,总面积18 165.4 hm2。植物区系属于长白山植物区系小兴安岭亚区动植物区系,属古北界—东北区—长白山亚区,是我国温带阔叶红松林分布中心区域北缘。保护区主要以保护红松为主,保存着目前世界上最为典型而完整的以红松为主的北温带原始针阔叶混交林生态系统,红松的平均年龄约220 a。原始阔叶红松林是其最典型和最稳定的植被类型,作为小兴安岭地带性植被的顶极群落,这种林分对整个生态系统的周期性波动变化和稳定结构的维持十分重要,生态价值极为珍贵[15]。
该地区属于北温带大陆性季风气候,受森林和局部地形影响,四季气候变化很大。冬季气候严寒、干燥而漫长;夏季雨水充沛,气候湿热,时有洪涝和低温冷害发生;春季和秋季天气多变,春季降水少、多大风,易发生干旱;秋季常有冻害和早霜发生。独特的地理环境形成了该区特有的森林环境类型,其气象资料较能代表小兴安岭林区的背景值。
1.1.2 数据资料
红松种子产量数据来源于丰林国家级自然保护区,数据时间段为1999~2014年,方法为野外调查法。气候数据来源于黑龙江省伊春市五营区气象局,资料包括气温(T)、降水量(P)和日照时数(S),气温和降水时间段为1958~2014年,仪器自动观测,每分钟1个数据,数据最小单位为0.1℃和0.1 mm。日照时数时间段为1960~2014年,自记日照,每日人工进行日照纸更换,数据最小单位为0.1 h。数据按月、季、年进行统计,温度取平均值,降水和日照求和。季节根据当地气候进行划分,冬季(上1年12月~2月)、春季(3~5月)、夏季(6~8月)、秋季(9~11月)。
1.2 研究方法
1.2.1 红松种子产量野外调查法
保护区内的红松大部分都生活在云杉冷杉—红松林(Piceasp.-Abiesnephlolepis-Pinuskoraiensis)、椴树—红松林(Tiliaamurensis-Pinuskoraiensis)、枫桦—红松林(Betulacostata-Pinuskoraiensis)和蒙古栎—红松林(Quercusmongolica-Pinuskoraiensis)4个具有代表性的原始阔叶红松林群落中,但不同的群落类型间红松的生境具有很大的差异[16]。为了消除群落生境差异对红松种子产量带来的影响,调查样地按4个群落类型进行系统抽样随机选取。每个群落类型分别随机选取5块样地,共20块样地进行调查,每块样地面积50 m×50 m,样地内红松株数不少于30株。在每年9月份红松种子成熟时,对标准木球果进行逐株采摘,去果皮称重计算,求出单株平均种子产量,乘以丰林保护区的红松总株数,得出红松种子年产量。
1.2.2 资源限制和资源匹配分析
农业上,在较长时间序列的植物产量与气候因子关系的观测统计研究中,Nicholls[17]于1997年提出了对产量进行分解的建议,并被众多学者应用于农作物产量的气候影响评估[18~19],研究一般把产量分解为趋势产量(Yt)、气候产量(Yw)和随机误差(ΔY)3部分,Y=Yt+Yw+ΔY。本文在研究红松种子产量与气象因子关系时,结合资源限制和资源匹配假说,也将红松种子产量分解为趋势产量(Yt)和气候产量(Yw)。
根据资源限制假说[6~8],林木结实的自我资源限制,在外表现为产量本身对之后年份产量的限制性。因此我们选取红松种子产量作为研究对象,进行红松种子产量年际间的相关分析,并对红松产量与其之前年份的种子产量之和进行相关分析,来验证红松资源限制机制的存在,找出影响红松结实资源限制性的关键指标,计算红松结实的趋势产量(Yt)。研究地点原始阔叶红松林林分较为稳定,相对其它林分,气候影响较小[20],资源限制在正常年份规律性较强,有利于资源限制影响的分析及剥离。
红松从花原基形成到种子成熟过程历时近3年,当年成熟的球果,其花原基在2年前(前年)的夏季便开始形成[21],这时的气象条件对红松花原基形成量有着重要影响,将直接影响雌球花的数量,进而影响球果形成和种子产量;花原基形成后红松在第2年(去年)夏初时开花、授粉、幼果形成,然后经历短暂的发育和越冬,最终在当年受精发育球果成熟。因此去年和当年的气候条件对红松结实也有着重要影响。我们将红松当年的气候产量(Yw)与前年、去年和当年不同时段(月、季、年)的气温(热量)、降水(水分)和日照时数(光照)分别进行相关分析,并结合红松生殖发育阶段进行讨论,找出红松结实的气候敏感阶段及其敏感的气象指标。
1.2.3 相关分析数据处理
为了便于不同要素值间的比较,本文在相关分析前对红松种子产量和气象数据均进行了Z-score标准化处理,将各个序列值转为无量纲值。Z-score标准化[22]又称标准差标准化,经过处理的数据符合标准正态分布,即均值为0,标准差为1,其转化函数为式(1):
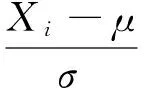
(1)
式中:μ为所有样本数据的均值;σ为所有样本标准差;Xi为标准化处理值,Yi为标准化处理过后的数值。
本文数据处理软件采用spss 19.0和excel 2003数学软件,绘图使用origin 8.0完成。
2 研究结果
2.1 五营气候变化特征
五营年平均气温呈增加趋势,年平均降水则呈下降趋势(图1),趋势倾向率分别为0.03℃·a-1和-1.79 mm·a-1,气候趋于高温干旱化,1999年之后趋势有所缓解,但2007和2008年出现了极端异常的高温干旱气候。2007和2008年平均气温分别为2.1和1.9℃,比平均值高出2.0和1.8℃。降水量分别为437.9和422.3 mm,比平均值少了194.8和210.4 mm。五营年日照时数趋势变化不明显,略呈增加趋势,趋势倾向率为0.40 h·a-1,1999年之后日照有所缩短,趋势倾向率为-6.35 h·a-1。

图1 五营林区气温、降水和日照时数的年际变化Fig.1 Yearly change of temperature,precipitation and sunshine hours in Wuying
2.2 红松结实规律及资源限制
总体上自1999年以来丰林自然保护区红松种子产量年际动态变化并不是通常认为的三年一小收,到五年一大收(图2)。当年(c)红松种子产量与去年(b)和前年(a)的种子产量均呈负相关(rb=-0.33,ra=-0.44),并且受气候异常的影响,2007和2008年种子产量出现异常波动,结实规律随之调整,产量值的标准偏差开始增大。分析当年红松种子产量与其之前年份的种子产量和相关结果显示,去年和前年的红松种子产量和(d)负相关达到最大(rd=-0.67,P<0.01)。分析发现去年和前年产量和增多,对当年结实具有限制性,导致当年红松种子产量减少,当去年和前年产量和大于200万千克时,红松种子产量均较少(除2008年,极端高温干旱),为歉收或绝收年;然而去年和前年产量和减少,限制性降低,红松种子产量一般维持在高位,但与去年和前年产量和趋势并不完全一致。可见红松结实存在资源限制机制,去年和前年产量和是影响红松结实资源限制性的关键指标,而且资源限制机制并不是影响红松种子产量的唯一因素。
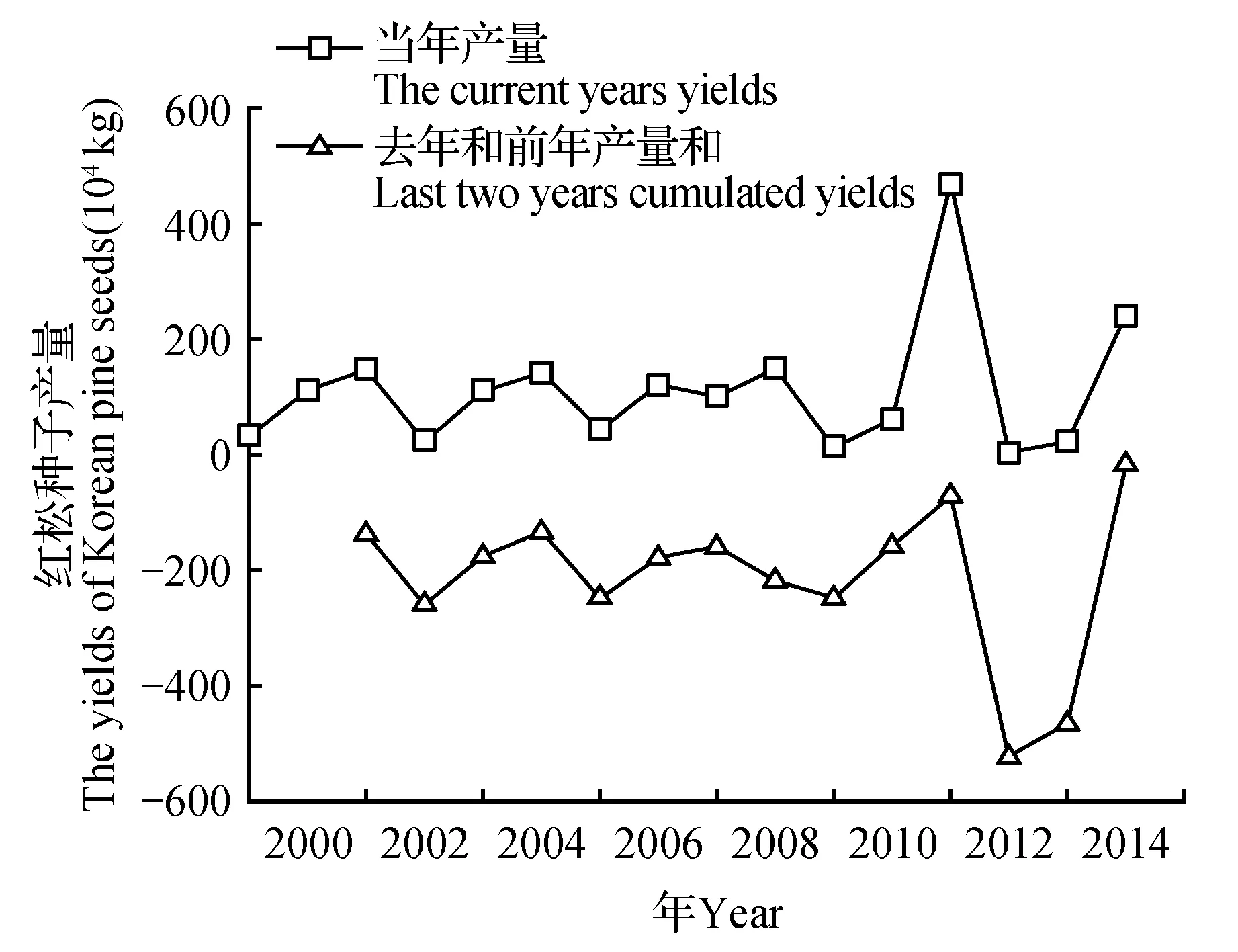
图2 丰林自然保护区阔叶红松林1999~2014年红松种子产量年际变化 当年为红松采摘年份;去年为当年的上一年;前年为去年的上一年;去年和前年产量和为去年红松种子产量与前年红松种子产量相加的累积产量。Fig.2 Yearly change of the yields of Korean pine seeds in the broad-leaved Korean pine forest in Fenglin National Nature Reserve during 1999-2014 The current year is the year of Korean pine seeds gathering; Last year is the year before the current year;The year before last year is the previous year of last year;Last two years cumulated yields are the cumulative yields of last year yields and The year before last year yields.
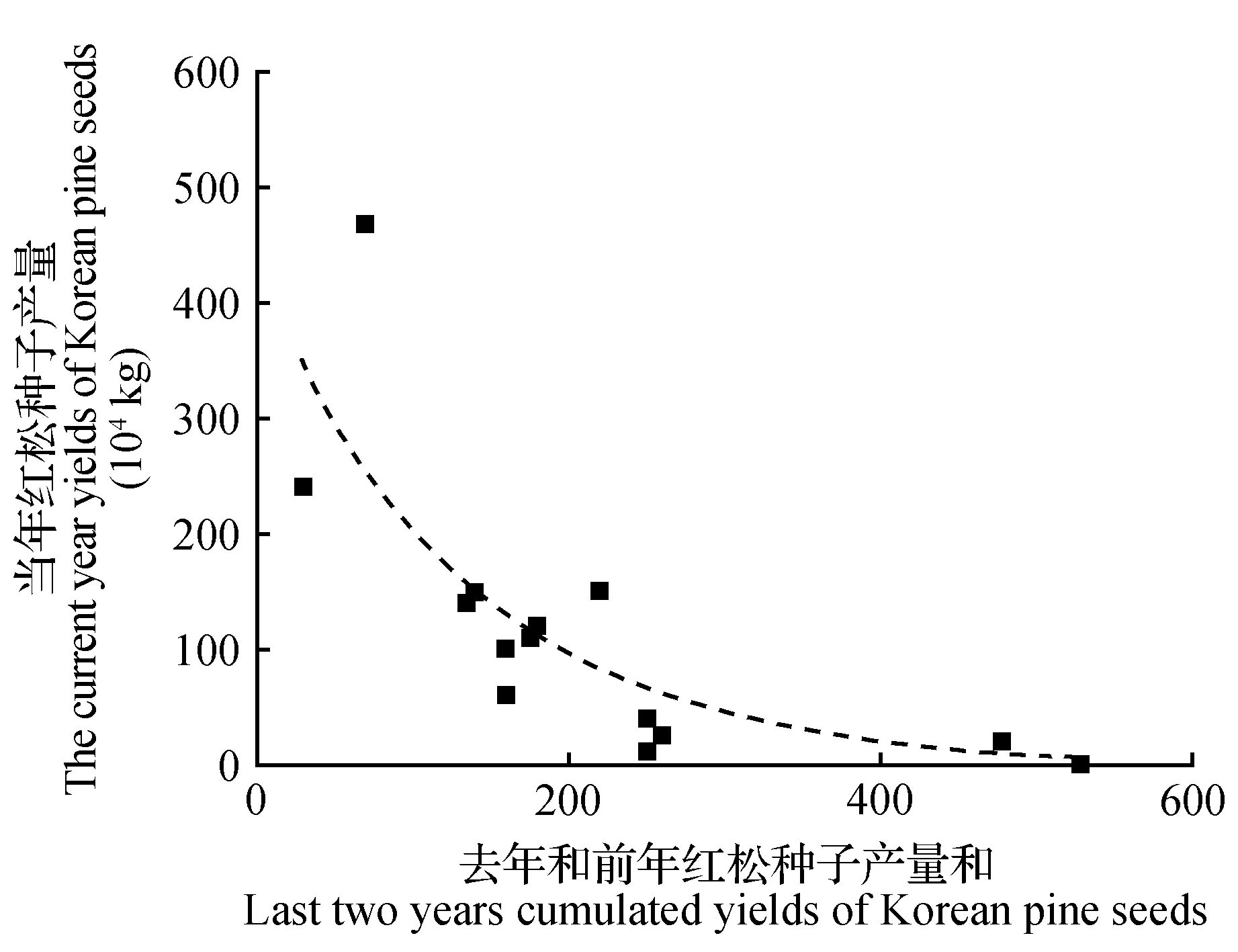
图3 丰林自然保护区2001~2014年当年红松种子产量与去年和前年种子产量和的关系Fig.3 Relationship between the current year yields and last two years cumulated yields of Korean pine seeds in Fenglin National Nature Reserve during 2001-2014
为了提取红松结实的资源限制分量,计算趋势产量,我们进一步分析了当年红松种子产量与去年和前年产量和的关系,发现两者并不是简单的线性关系,当年红松种子产量随去年和前年产量和呈指数曲线变化(图3)。我们用去年和前年产量和对当年种子产量进行拟合,计算红松结实的资源限制分量,得出趋势产量:Yt=443.87×e-0.007 78×(Yi-1+Yi-2)(Yi-1和Yi-2分别为去年和前年的种子产量,R2=0.60),并分解出气候产量。分析认为趋势产量可看作是红松种子产量变化的固有规律,气候产量则是对趋势产量的调整(图4),在不同气候条件影响下,这种调整不但包括增加实际产量也包括减少实际产量。
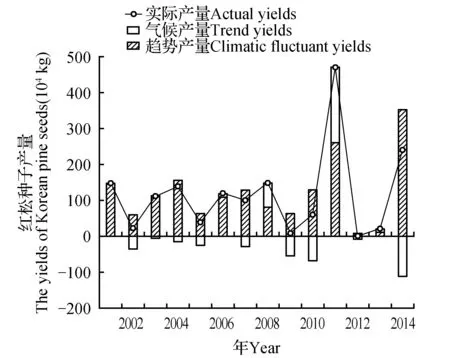
图4 丰林自然保护区2001~2014年红松结实趋势产量和气候产量的关系Fig.4 Relationship between the trend yields and climatic fluctuant yields of Korean pine seeds in Fenglin National Nature Reserve during 2001-2014
2.3 气象条件对红松结实的影响
本文根据刘一樵[21]和王振宇[2]等研究,结合2012~2014年对五营林区红松物候的实际观测结果,将红松生殖发育过程分为花原基形成(前年6~8月)、花原基越冬(前年11月~去年2月)、花芽形成(去年5月)、开花授粉幼果形成(去年6月)、幼果越冬(去年12月~当年2月)、幼果受精(当年6月)、胚成熟(当年7月)和种子发育成熟(当年9月)共8个阶段,并结合红松8个生殖发育阶段分析气象因子对红松气候产量的影响。结果表明(表1),气温、降水和日照对红松有性生殖过程的不同阶段均有不同程度的影响。
2.3.1 温度变化对红松结实的影响
相关分析显示(表1),当年红松气候产量与前年夏季6~8月(花原基形成期)平均气温呈极显著负相关,其中与前年6月平均气温呈显著负相关。与此结论一致,同样拥有3年生殖周期的柏木陆均松(D.cupressinum)、欧洲山毛榉(FagussylvaticaL.)和美国食松(Pinusedulis)在种子年发生年的前年夏季也出现了温度异常偏低现象[23~25];当年红松气候产量与去年4月平均气温(花芽形成期前期)呈显著负相关;与去年6月(开花授粉幼果形成期)平均气温呈极显著正相关,有研究表明高温可能会减少红松授粉不足现象,增加座果率,提高红松种子产量[26];当年红松气候产量与当年7月(胚成熟期)平均气温呈极显著正相关,表明7月高温有利于红松种子胚成熟[2];此外,当年红松气候产量还与当年9月(种子发育成熟期)平均气温呈显著负相关。
2.3.2 前两年6月温差与红松结实的关系
分析发现红松气候产量与前年夏季(花原基形成期)、前年6月、去年夏季、去年6月(开花授粉幼果形成期)平均气温的相关系数大多都显著相关且符号相反。计算去年夏季和前年夏季平均温度的差值(Δt=Tb6-b8-Ta6-a8)、去年6月和前年6月平均温度的差值(Δt6=Tb6-Ta6),它们与当年红松气候产量均呈极显著正相关(rΔt=0.73,rΔ6=0.74,P<0.01)。Δt6显著性略大,对红松产量具有很好的预测性,对图2中红松种子产量异常变化年份给出了很好的解释,如2010和2014年产量异常减少,Δt6也异常减小,分别为-3.9℃和-0.9℃。2008和2011年产量异常增加,Δt6也异常增大,分别为3.1℃和6.1℃,绝对值均远远大于0.04℃的平均值。
2.3.3 降水变化对红松结实的影响
相关分析显示(表1),当年红松气候产量与前年夏季6~8月(花原基形成期)降水、前年6月降水均呈极显著正相关,Mutke[11]研究也发现地中海石松在其花原基形成年份良好的水分条件有利于其结实。Smaill[12]的研究结论相同,并分析认为降水促进了氮的利用,提高了产量。当年红松气候产量与去年6月(开花授粉幼果形成期)降水不相关,根据相关研究[27]降水量的确对植物的开花授粉会产生一定的影响,原因可能是红松开花授粉期一般较短,降水对授粉影响,只与授粉当日降水指标有关,并且只有达到一定级别的降水量才能影响开花授粉;当年红松气候产量还与去年12月(处于幼果越冬期之内)降水呈极显著正相关。
2.3.4 日照时数变化对红松结实的影响
相关分析显示(表1),当年红松气候产量与前年5月(花原基形成期前期)日照时数正相关达到显著水平;与前年夏季6~8月(花原基形成期)日照时数呈显著负相关,其中与前年6月日照时数相关达到极显著水平,可见光周期的变化可能对红松顶芽中花原基形成有着重要影响。当年红松气候产量还与去年冬季(前年12月~去年2月)日照时数呈显著负相关。
表1丰林自然保护区阔叶红松林2001~2014年红松气候产量与气象因子的关系
Table1ThecorrelationbetweenclimaticfluctuantyieldsofKoreanpineandmeteorologicalfactorinthebroad-leavedKoreanpineforestinFenglinNationalNatureReserveduring2001-2014year
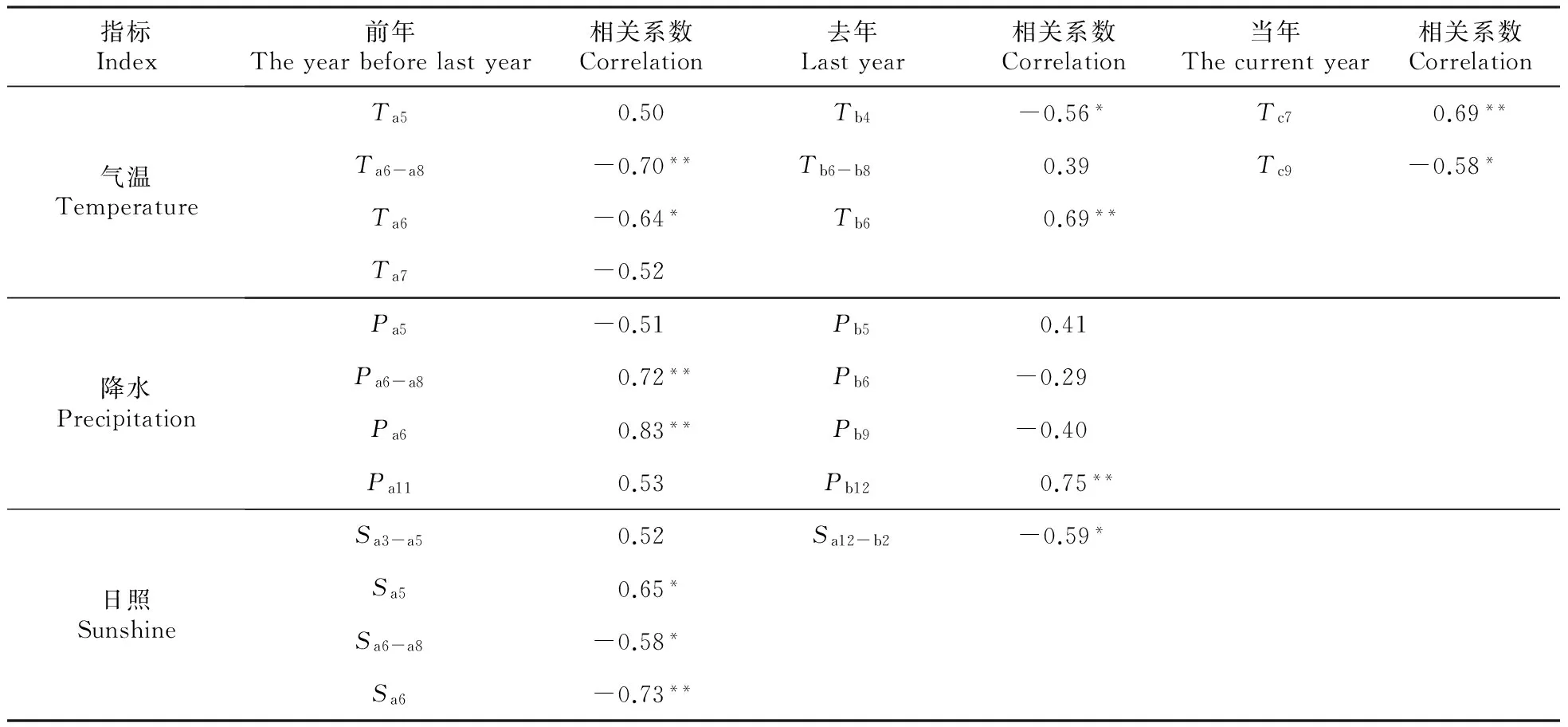
指标Index前年Theyearbeforelastyear相关系数Correlation去年Lastyear相关系数Correlation当年Thecurrentyear相关系数Correlation气温TemperatureTa50.50Tb4-0.56*Tc70.69**Ta6-a8-0.70**Tb6-b80.39Tc9-0.58*Ta6-0.64*Tb60.69**Ta7-0.52降水PrecipitationPa5-0.51 Pb50.41Pa6-a80.72**Pb6-0.29Pa60.83**Pb9-0.40Pa110.53Pb120.75**日照SunshineSa3-a50.52Sa12-b2-0.59*Sa50.65*Sa6-a8-0.58*Sa6-0.73**
注:当年(c)为红松采摘年份;去年(b)为当年的上一年;前年(a)为去年的上一年;字母下角数字代表月份;*在P=0.05水平上显著;**在P=0.01水平上显著。
Note:The current year(c) is the year of Korean pine seeds gathering; Last year(b) is the year before the current year;The year before last year(a) is the previous year of last year;The numbers at the right foot of letters represent month;*,P<0.05;**,P<0,01,only significant relationships are shown.
3 讨论
3.1 红松资源限制机制
本研究中,红松种子产量与去年和前年产量和呈显著负相关,如果去年和前年产量之和增加,当年种子产量则相应减少,表明红松结实受到了自身限制,意味着红松结实存在资源限制机制。Sanguinetti[7]对智利南洋杉(Araucariaaraucana)的研究发现树木在个体水平上,种子产量同样存在0~2年负的自相关。在关于开心果[28]的研究中也发现了类似结论。目前,关于种子年资源限制机制的机理研究还没有统一的定论。这种不同年份产量间的限制关系究竟是发生于年际间,即种子年大量消耗林木储存的资源减少了下一年结实的可利用资源;还是发生于同一年份内,即花原基形成、幼果形成和球果成熟等不同生殖阶段间的资源限制,进而影响了不同年份的产量,尚有待于进一步研究。Drobyshev[8]发现欧洲山毛榉的径向生长只有在种子年才受到抑制,但资源消耗在下一年之前即可得到恢复,可见种子年资源限制跨年存在的可能性较低。并且我们假设红松存在这种年际间结实的资源限制,种子年后储存资源减少限制了下一年种子产量,红松年年结实,资源年年消耗,将不会出现下一次的种子年现象,与观测结果相矛盾。因此资源限制更可能发生在同年份内不同生殖阶段间。有研究表明地中海石松(Pinuspinea)[11]种子年的发生与花原基形成年份种子的低产量密切相关,说明花原基形成与同一年球果成熟间可能存在资源限制关系。Sala[6]进行的白皮松(Pinusalbicaulis)种子年营养消耗试验表明:在种子年,N或P减少只发生在繁殖枝的部分,然而在下一年营养在所有枝上都减少,即球果成熟在营养利用中可能具有优先权,即为单向限制。而这种单向限制很可能与植物内源激素的动态平衡有关[29]。并且机理原因可能是促进果实成熟而不是抑制花原基形成。因为假设种子年抑制了花原基形成,那么即使进行施肥也不会使种子产量增加。然而如果种子年的作用是通过优先促进果实的成熟,降低了营养可利用量而影响了种子产量,施肥在增加当年产量的同时会大幅促进花原基形成量而增加2年后的产量,在Smaill[12]对山毛榉(Nothofagussolandrivar.cliffortioides)的氮肥施入实验中证实了这种观点。
通过分析,我们认为不同年份间种子产量的限制关系的根本原因可能是球果成熟对同年其它生殖发育的单向自我资源限制关系。假设每年红松吸收和储存的营养为定值,由于花原基形成(2年后结实)、开花及幼果形成(1年后结实)和球果成熟(当年结实)这三个生殖过程在一年中同时存在,如果综合条件有利于红松球果大量成熟,出现种子年现象,那么花原基形成、开花及幼果形成就会受到资源限制,进而影响之后年份的红松种子产量。红松的这种开花结实规律则可能是红松资源分配的一种权衡策略。
3.2 红松资源匹配机制和气候敏感期
本研究中,当年红松种子产量与去年和前年产量和趋势并不完全一致,说明红松内部的资源限制机制并不是影响红松种子产量的唯一因素,还可能存在资源匹配机制。Schauber等[23]在对新西兰的17种不同植物研究中发现同一地域它们的结实具有种间同步性,证明了资源匹配机制的存在。本研究中,当年红松气候产量在部分生殖发育阶段内对气候变化均表现敏感,与单个或者多个气象因子相关都达到了显著,说明红松结实也存在资源匹配机制。我们把这些关键的生殖发育阶段称为气候敏感期。红松结实气候敏感期和敏感气象指标可用于未来红松种子产量预测,也可结合当地红松生殖发育期通过对其他地区产量预测加以检验和应用。
综合分析,花原基形成期前期(前年5月)日照,花原基形成期(前年6~8月)平均气温、降水和日照,花芽形成期前期(去年4月)平均气温,开花授粉幼果形成期(去年6月)平均气温,幼果越冬期(前年12~去年2)去年12月降水,胚成熟期(当年7月)平均气温和种子发育成熟期(当年9月)平均气温均为红松结实的气候敏感期和敏感气象指标。平均气温的影响时间范围(对3年生殖发育均有显著影响)要大于降水和日照。花原基形成期是红松结实最为关键的气候敏感期,红松气候产量与此阶段气温和日照呈极显著负相关,与降水呈极显著正相关。我们认为红松结实的资源匹配机制与气候资源的可利用性有关,气象指标对结实的影响也随着气候资源可利用性的变化而变化。
3.3 红松结实的气候脆弱性
Kelly研究发现去年夏季与前年夏季温度的差值(Δt)对种子年现象具有很好的预测效果[10]。本研究中也得到类似结论,去年和前年夏季平均气温差值Δt、去年和前年6月份平均气温差值Δt6均与红松气候产量呈极显著正相关,Δt6显著性略大。分析认为原因是当年红松气候产量与前年6月(处于花原基形成期)和去年6月(开花授粉幼果形成期)平均气温相关系数的符号相反,即平均气温低有利于花原基形成,而平均气温高则有利于开花授粉幼果形成,Δt6越大气候产量越高。因此红松的生殖发育只适应于一定的温度范围内,温度过高将影响花原基的形成,温度过低则可能会对开花授粉幼果形成不利。由此可推论,在当今极端气候频发的背景下,气温年际间变异系数将增大,Δt6极端变化的概率逐渐加大,这将导致红松种子产量也将趋于极端变化。
我们认为,红松种子产量的高低主要取决于红松球果数量和种子发育质量两方面。而红松花原基形成的数量和质量作为基数对未来最终红松球果数量具有决定性影响,是红松种子产量的重要基础。五营区平均气温在1987年左右发生了突变,由偏冷期跃变为偏暖期[30]。突变前后五营6月份平均气温(1958~1986年,1987~2015年)分别为16.6和17.5℃,突变后升高了0.9℃。近16年来,丰林保护区红松高产年份(大于150万千克)的前年6月平均气温(处于花原基形成期)为16.6℃,与突变前平均气温相同;而其它低产年份则为18.0℃,比突变前高1.4℃,比突变后也高出0.5℃,受到了高温的影响。这表明在气候变暖的大背景下,部分年份6月异常高温已经超出了花原基形成的最适温度范围,导致了种子产量的下降。Mutke对地中海石松40年观测记录分析显示,气候变暖也导致了石松产量的显著减少,并可能发生在广阔的针叶树种中[11]。可见随着气候变化,红松种子产量将成为原始红松林更新重要的瓶颈,对气候变暖和极端气候具有一定的脆弱性。
3.4 植物种子年机制探讨及森林管理应用
本研究中,原始阔叶红松林红松种子年现象受到了资源限制机制和资源匹配机制的共同作用,影响了红松有性生殖不同阶段的发育,且资源限制影响要大于资源匹配机制。这与Lyles[5]和Smaill[12]的观点一致。这可能是因为原始阔叶红松林林分稳定,对气候变化的影响具有较强的适应性。但是随着气候变化加剧,资源匹配影响可能逐渐增大(如图4)。当气候因子达到一定阈值,则对红松结实具有决定性影响。我们认为,红松结实规律可能有2种情况:在平常年份,外界环境波动不大,资源限制机制占主导,而资源匹配机制不明显,红松结实周期即为3年的生殖周期,并且较为稳定;在气象灾年(如2007和2008年极端高温干旱),外界环境尤其是在植物气候敏感期内波动剧烈,资源匹配机制将占主导,红松种子产量会随着环境而变化异常,结实规律随之调整,周期不稳定。本研究在分析种子年机制的基础上还提供了趋势产量和气候产量的计算指标,为今后红松种子产量的预测提供了重要依据。将有利于红松种子资源管理,为种子年的种子储存和控制采摘提供参考,以保证红松林的正常更新和相关动物的生存及生态系统多样性的维持。
值得指出的是,本研究基于当年种子产量与去年和前年种子产量和的显著负相关关系以及种子产量与主要气象因子的关系推论出红松种子年现象可能受资源限制与资源匹配的共同影响。而确切详细的机理解释有待进一步深入研究,如红松结实规律的年际变化与树木生长指标、营养状况(N、碳水化合物等)、激素水平等之间的关系,与营养生长和生殖生长之间的关系,与各生殖构件之间的关系,以及与环境因子的关系等均需进行深入细致的研究。
1.中华人民共和国国务院.国家重点保护野生植物名录(第一批)[J].植物杂志,1999(5):4-11.
The State Council of the People’s Republic of China.National key preserved wild plants[J].Plant Journal,1999(5):4-11.
2.王振宇,刘文欢,谢伟,等.红松开花结实与气象因子相关规律的研究[J].东北林业大学学报,1992,20(S):118-120.
Wang Z Y,Liu W H,Xie W,et al.Research on the correlation between the Korean pine flowering and meteorological factors[J].Journal of Northeast Forestry University,1992,20(S):118-120.
3.赵振卿,王建华.红松种子产业的开发前景及措施[J].农村实用科技信息,2010(4):34.
Zhao Z Q,Wang J H.The development prospect and measures of Korean pine seeds industry[J].Rural Practical Science and Technology Information,2010(4):34.
4.Kelly D.The evolutionary ecology of mast seeding[J].Trends in Ecology & Evolution,1994,9(12):465-470.
5.Lyles D,Rosenstock T S,Hastings A,et al.The role of large environmental noise in masting:general model and example from pistachio trees[J].Journal of Theoretical Biology,2009,259(4):701-713.
6.Sala A,Hopping K,Mcintire E J B,et al.Masting in whitebark pine(Pinusalbicaulis) depletes stored nutrients[J].New Phytologist,2012,196(1):189-199.
7.Sanguinetti J,Kitzberger T.Patterns and mechanisms of masting in the large seeded southern hemisphere coniferAraucariaaraucana[J].Austral Ecology,2008,33(1):78-87.
8.Drobyshev I,Övergaard R,Saygin I,et al.Masting behaviour and dendrochronology of European beech(FagussylvaticaL.) in southern Sweden[J].Forest Ecology and Management,2010,259(11):2160-2171.
9.Woodward A,Silsbee D G,Schreiner E G,et al.Influence of climate on radial growth and cone production in subalpine fir(bieslasiocarpa) and mountain hemlock(Tsugamertensiana)[J].Canadian Journal of Forest Research,1994,24(6):1133-1143.
10.Kelly D,Geldenhuis A,James A,et al.Of mast and mean:differential-temperature cue makes mast seeding insensitive to climate change[J].Ecology Letters,2013,16(1):90-98.
11.Mutke S,Gordo J,Gil L.Variability of Mediterranean stone pine cone production:yield loss as response to climate change[J].Agricultural and Forest Meteorology,2005,132(3-4):263-272.
12.Smaill S J,Clinton P W,Allen R B,et al.Climate cues and resources interact to determine seed production by a masting species[J].Journal of Ecology,2011,99(3):870-877.
13.Mccarthy J J.Climate Change 2001:Impacts,adaptation,and vulnerability:contribution of working group II to the third assessment report of the intergovernmental panel on climate change[M].Cambridge:Cambridge University Press,2001:81-111.
14.Mooney K A,Linhart Y B,Snyder M A.Masting in ponderosa pine:comparisons of pollen and seed over space and time[J].Oecologia,2011,165(3):651-661.
15.李俊清,崔国发,臧润国.小兴安岭五营林区森林生态系统经营研究[J].北京林业大学学报,2000,22(4):25-34.
Li J Q,Cui G F,Zang R G.Studies on forest ecosystems management of Wuying forest areas in the lesser Xing’an mountains of Northeast China[J].Journal of Beijing Forestry University,2000,22(4):25-34.
16.王天明.丰林自然保护区阔叶红松林生物多样性研究[D].哈尔滨:东北林业大学,2004.
Wang T M.Study on biodiversity of broad-leaved Korean pine forest in Fenglin Nature Reserve[D].Harbin:Forestry College of Northeast Forestry University,2004.
17.Nicholls N.Increased Australian wheat yield due to recent climate trends[J].Nature,1997,387(6632):484-485.
18.Peltonen-sainio P,Jauhiainen L,Trnka M,et al.Coincidence of variation in yield and climate in Europe[J].Agriculture,Ecosystems & Environment,2010,139(4):483-489.
19.El-maayar M,Lange M A.A Methodology to infer crop yield response to climate variability and change using long-term observations[J].Atmosphere,2013,4(4):365-382.
20.丘阳,高露双,张雪,等.气候变化对阔叶红松林不同演替阶段红松种群生产力的影响[J].应用生态学报,2014,25(7):1870-1878.
Qiu Y,Gao L S,Zhang X,et al.Effect of climate change on net primary productivity of Korean pine(Pinuskoraiensis)at different successional stages of broad-leaved Korean pine forest[J].Chinese Journal of Applied Ecology,2014,25(7):1870-1878.
21.刘一樵,李士汤.森林植物学[M].北京:中国林业出版社,1993.
Liu Y Q,Li S T.Tree botany[M].Beijing:China Forestry Publishing,1993.
22.吴丽,陈薇,马杰,等.1961~2010天水市光、温、水资源变化特征及其与粮食产量的关系[J].中国农学通报,2014,30(17):245-249.
Wu L,Chen W,Ma J,et al.A study on variation characteristic of light,temperature and precipitation resource and their relationship with grain yield in Tianshui City from 1961 to 2010[J].Chinese Agricultural Science Bulletin,2014,30(17):245-249.
This study was founded by National Key Basic Research 973 Program(2010CB951301);National Natural Science Foundation of China(31270494)
introduction:CHENG Chun-Xiang(1982—),female,doctoral candidate,Engaged in researches of botany.
date:2016-11-08
SensitivityofFruitingforPinuskoraiensistoClimateChangeandMechanismsofMastingintheOriginalBroad-leavedKoreanPineForestinNorthXiaoxing’anMountain,China
CHENG Chun-Xiang1,2MAO Zi-Jun1*JIN Shi-Bo2SONG Guo-Hua3SUN Peng-Fei2SUN Tao1,4
(1.Key Laboratory of Forest Plant Ecology,Northeast Forestry University,Harbin 150040;2.Wuying Meteorological Bureau/Wuying Forest Ecosystem and Agro-meteorology Experimental Research Station,Yichun 153033;3.Fenglin National Nature Reserve Administrative Bureau,Yichun 153033;4.Institute of Applied Ecology,CAS,Shenyang 110016)
To find out sensitivity of fruiting for Korean pine(Pinuskoraiensis) to climate change and mechanisms of masting, we studied Korean pine in the original broad-leaved Korean pine forest which has high stand stability and the yields of Korean pine seeds in nearly 16 years. We decomposed the yields of Korean pine seeds into trend yields and climatic fluctuant yields by resouce switching theory, resource matching theory and the suggestion of Nicholls. Then, we analyzed the influence of weather on climatic fluctuant yields. The results reveal:(1)The current year yields of Korean pine seeds significantly negative correlated with the combined yields of the last 2 years and showed exponential relationship with it, which supports the resource switching hypothesis. And resource switching probably acts as Korean pine cone maturating inhibition among the reproductive processes in a mast year. (2)The temperature difference between the two previous Junes(ΔT6) better predicts the yields of Korean pine seeds. (3)The period of bud meristerms development for Korean pine fruiting was the most key climate-sensitive phase. Negative temperature and sunshine duration, coupled with positive precipitation in the period significantly correlated with climatic fluctuant yields of Korean pine. (4)Positive temperature anomaly in some years was beyond the optimum temperature range of the development of bud meristerms and caused reproduction of Korean pine to fall. (5)The yields of Korean pine seeds in many periods of reproductive development was sensitive to climate change, indicating that resource switching and resource matching may be exhibited by a single masting. These findings suggest Korean pine reproduction are vulnerability to climate warms and may be an increasingly important bottleneck for future Korean pine regeneration, especially in areas with greater increases in temperature.
Pinuskoraiensis;Xiaoxing’an Mountain;masting;climatic fluctuant yields;climate sensitivity
科技部973国家重点基础研究项目发展计划项目(2010CB951301);国家自然科学基金资助项目(31270494)
程春香(1982—),女,博士研究生,主要从事植物学研究。
* 通信作者:E-mail:zijunm@nefu.edu.cn
2016-11-08
* Corresponding author:E-mail:zijunm@nefu.edu.cn
S791.247
A
10.7525/j.issn.1673-5102.2017.01.016
