古尔班通古特沙漠南缘3种生活型草本植物生物量分配及相关生长关系
2017-11-09汪溪远
赵 晶 汪溪远
(新疆大学资源与环境科学学院,绿洲生态重点实验室,乌鲁木齐 830046)
古尔班通古特沙漠南缘3种生活型草本植物生物量分配及相关生长关系
赵 晶 汪溪远*
(新疆大学资源与环境科学学院,绿洲生态重点实验室,乌鲁木齐 830046)
为了解古尔班通古特沙漠不同生活型草本植物的生物量分配特征,通过取样调查当地34种草本植物的生物量研究表明:(1)采集到的每个物种样本的总生物量大多小于4 g,其中多数个体(大于80%)的地上与地下生物量分别在2和0.5 g以内,根冠比主要集中在0.25以内;(2)类短命植物向地下部分分配较多生物量,根冠比较大,集中分布在1.67左右;短命和一年生长营养期草本向地下部分配的生物量远小于类短命植物,两者的根冠比较小,主要集中在0.15左右;(3)短命植物地下与地上部生物量间为等速生长关系,而类短命植物和一年生长营养期草本植物地下与地上生物量间为异速生长关系,其中类短命植物随个体增大向地下部分配生物量的比例增多,而一年生长营养期草本则相反。综上所述,短命、类短命和一年生长营养期草本植物地下与地上生物量间具有不同的分配特征,与各自独特的生活史特征相一致。
古尔班通古特沙漠;草本植物;生活型;生物量分配
生物量分配是指将营养器官同化的资源用于根、茎、叶、花、果等器官的比例,是植物权衡生殖与生存策略的结果,体现了植物的生活史特征[1~3]。生物量向植物不同器官的分配主要受物种、个体发育和环境因子的影响[4]。生物量在各器官间的分配比例和异速生长方程是分析生物量分配特征和类型的2种常用方法[5~8]。异速生长分析即相关生长分析,可用于描述某些生物学特征(如个体形态指标、器官生物量等)与个体大小之间的幂函数关系。目前,异速生长关系在刻画生物体构件结构与功能特征上得到了大量应用,其中以植物体构件(器官)生物量间的相关生长关系研究最为常见[9]。通过分析全球植物物种地上和地下生物量数据库(包括不同生境下的草本植物与树木,双子叶植物、单子叶植物和裸子植物)的研究表明,在自然条件下,种间和种内个体水平的地上与地下生物量间具有相同的分配速率[10]。
短命植物(ephemerals)、类短命植物(ephemeroids)和一年生长营养期植物(annuals)是古尔班通古特沙漠植物群落草本层的主要构建者,同时也是中国荒漠植被区系重要而独特的组成部分[11]。短命和类短命植物利用春季或夏季短期内较丰富的环境资源迅速完成生活周期,并留下种子越冬。其中,一年生的为短命植物,多年生的是类短命植物,其地上部分当年死亡并在地下部留下根茎次年再萌发生长[12~13]。短命和类短命植物出芽早且生长迅速,在其它荒漠植物尚未返青前,能够充分“填补”固定或半固定沙丘荒漠灌丛间的裸地,从而充分利用有限的资源,例如水分完成生命周期[14]。另外,短生长周期的特点使其在N、P、K、Ca等养分元素的积累和循环方面起到重要的“春坝”作用,即植物将养分暂时存在生物量中,随后再将其归还到土壤,这种作用对荒漠生态系统的平衡和稳定具有重大意义[15~16]。一年生长营养期草本植物出芽则较晚,能够保持较长生长期(4~8月),是一类旱生植物类群,尽管一年生草本植物物种较少,但在夏末和秋初时期,其较大的密度在维护荒漠生态系统稳定方面起到重要作用[11]。
目前,针对荒漠地区的短命植物、类短命植物及一年生草本的研究已有很多,如邱娟等对准噶尔荒漠早春短命植物的生物量分配特点进行了分析[17],李雪华等对科尔沁沙地70种草本植物个体和构建生物量进行了比较研究[18],艾沙江等对天山北麓类短命植物新疆郁金香生物量分配特点进行了分析[19],陶冶等对准噶尔荒漠6种类短命植物的生物量分配特征进行了研究[20]。不难看出,以往的研究多集中探讨某一类生活型植物的生物量分配特点,较少将不同生活型(短命、类短命和一年生长营养期草本)植物生物量分配特征进行对比研究,因此,对这3种生活型植物生物量分配规律及差异性的认识仍不清楚。为此,本研究对古尔班通古特沙漠南缘生长的短命、类短命及一年生草本植物按物种进行生物量的调查取样,通过分析地上、地下各器官的生物量、分配比例以及异速生长关系,探讨不同生活型植物间的生物量分配特点。
1 研究方法
1.1 研究区概况
古尔班通古特沙漠南缘地势呈东北高西南低,为温带干旱荒漠气候,干旱少雨。在古尔班通古特沙漠南缘的沙丘类型主要为沙垅,呈南北走向较多,一般高度在10~50 m。古尔班通古特沙漠南缘的主要植被类型为灌木、短命和类短命植物,其中灌木主要有梭梭(Haloxylonammodendron)、蛇麻黄(Ephedradistachys)、沙拐枣(Calligonummongolicum)、琵琶柴(Reaumuriasoongorica)等。研究区内以白梭梭(H.persicum)为优势种的植物群落占据沙丘中上部,蛇麻黄(E.distachya)群落主要分布在丘间低地和沙丘中下部,两种群落下层普遍生长有一年和多年生的草本及小半灌木。短命和类短命植物发育良好,主要种有尖喙牻牛儿苗(Erodiumoxyrrhynchum)、囊果苔草(Carexphysodes)、粗柄独尾草(Eremurusinderiensis)、角果藜(Ceratocarpusarenarius)等[21]。
1.2 野外数据采集
2015年4月底,在古尔班通古特沙漠南缘(44°12′~44°21′N,87°50′~87°54′)选取具有代表性的沙丘5处,通过样方调查法确定草本层的物种组成。经统计,研究区内共有34种草本植物,其中短命植物24种,类短命植物5种,一年生草本植物5种(表1)。植物采集均在其成熟期(开花末期)进行(即2015年5月中旬、5月底及6月底)不考虑繁殖器官的生物量分配比例。依据前期的样方调查结果,为每一个物种选择一处10 m×10 m典型样地,并划分为4块5 m×5 m的样方中,选择一个2.5 m×2.5 m的小样方进行植物取样。利用全株挖掘获取样地内全部物种的生物量[22],每个物种获取至少10株健康且完整的个体,植株个体较小的物种,选取5~10株合并为一个样品进行称重。根系挖掘的广度和深度因物种根系形态特征而异,短命植物个体矮小,根系分布较浅,一般在30 cm以内,因此,挖掘深度通常在30 cm,半径在10~20 cm;对类短命植物根系的挖掘方法见文献[20];对一年生植物的挖掘深度一般在30~50 cm,半径约20~30 cm。在获取包含根系的土块后,仔细除去附着在目标植物根系上的土壤及其他植物的根系,同时去除处于半分解状态的死根,整个过程保证地下根系的完整性。随后,将获得的植株装入封口袋,并放入冷藏箱带回实验室进行分割。地上部分分为茎、叶,根系样品用冷水冲洗干净后与地上部分分装在已做好标记的信封内,并置于75℃烘箱内烘48 h。将每株植物的地下根系、地上叶片及茎生物量单独称重、记录。所用天平精度为0.000 1 g。
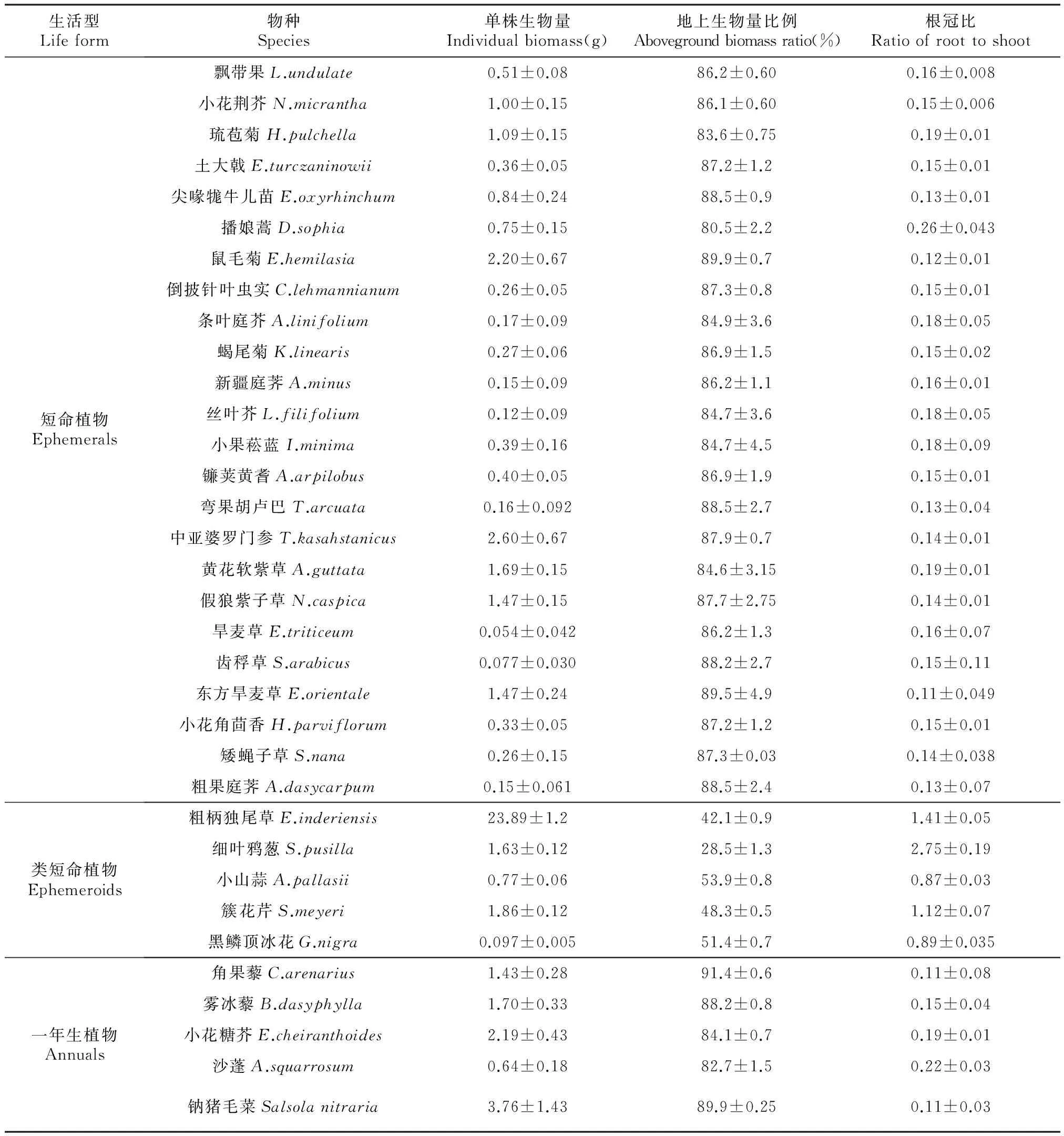
表1 34种草本植物的生物量及根冠比
1.2 数据分析与作图
常规数据的整理及作图分别在Excel 2010和Origin 9.0软件中完成。异速生长指数(a)、截距(lgβ)的计算,以及等速生长检验、共斜率检验以及a的差异性分析均使用SMATR软件包在R软件中完成。利用Ⅱ类线性回归(reduced major axis regression,RMA)计算回归指数(a)、95%置信区间(95% CI)及决定系数R2[9]。

图1 34种草本植物地上生物量、地下生物量及总生物量的频度分布Fig.1 Frequency distribution of individual aboveground biomass,belowground biomass and total biomass of 34 herbs
2 结果与分析
2.1 草本植物个体的生物量分配特点
研究区共采集到34种草本植物,其中短命植物24种,类短命植物5种,一年草本5种。每种植物平均采集不少于10株,共410株。大部分荒漠草本植物单株生物量较小(图1)。地上生物量大部分在2 g以内(占82%),其中地上生物量小于1 g的植株数占70%(图1A)。地下生物量(根系)超过80%植株个体在0.5 g以内,其中小于0.25 g的植株个体占71.5%(图1B)。总生物量大多小于4 g,其中小于2 g的植株占到78%(图1C)。草本植物根冠比主要集中在0.25以内,所占比例高达72%(图2)。
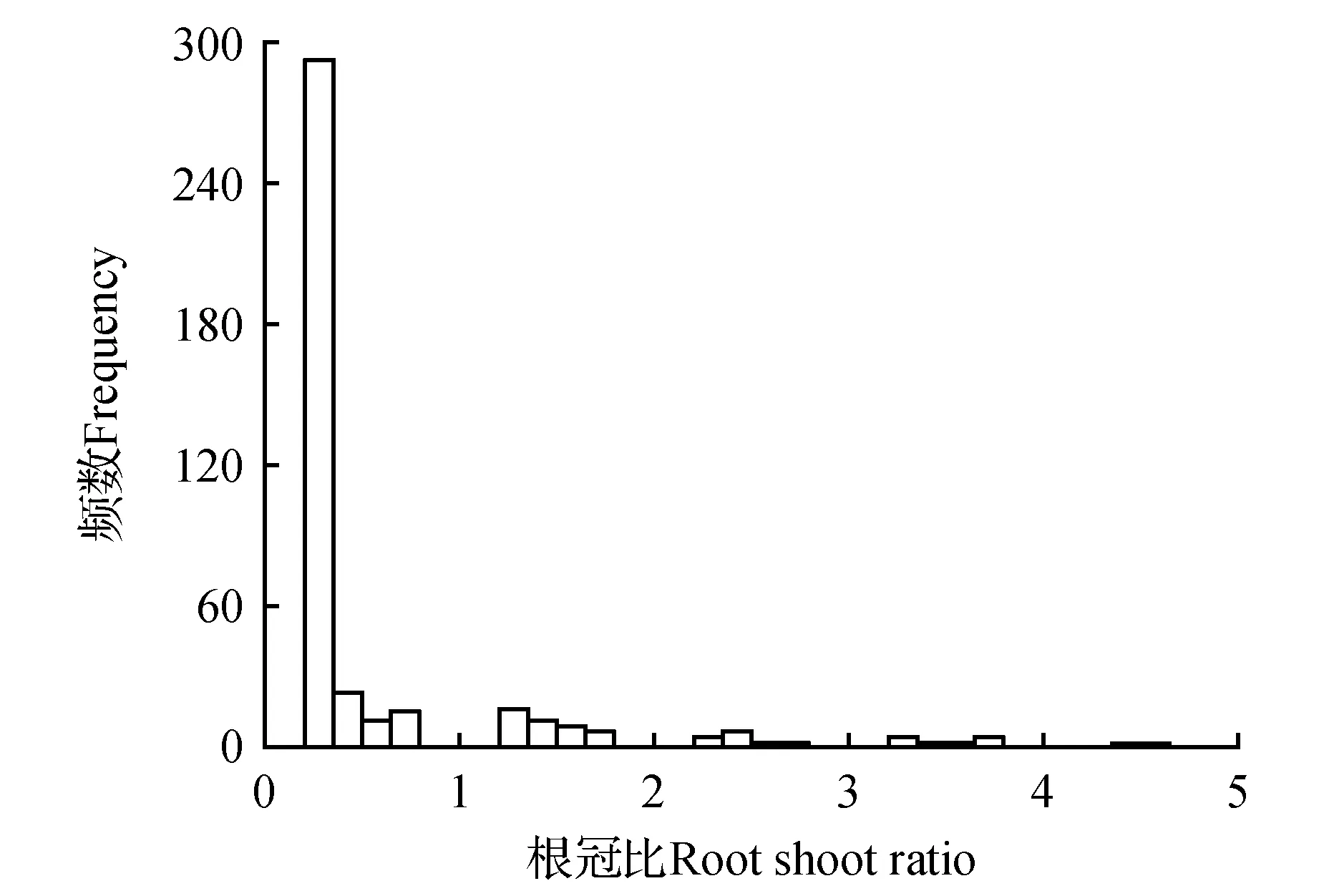
图2 34种草本植物根冠比(R/S)的频度分布Fig.2 Frequency distribution of root to shoot ratio of 34 desert herbs
类短命植物根冠比较大,如粗柄独尾草和细叶鸦葱的冠比分别为1.41、2.75;此外,类短命植物小山蒜的根冠比虽较小,但仍远远大于短命植物和一年生植物,后两者的根冠比一般在0.11~0.26(表1)。此外,大部分短命和一年生草本植物地上生物量分配比例较高,均在80%以上,而类短命植物地上生物量分配比例则较小,其中小山蒜的地上生物量分配比例最高,为53.9%,细叶鸦葱最小,约为28.5(表1)。
2.2 3种生活型草本植物的生物量分配特征
短命、类短命和一年生草本植物的地上生物量、地下生物量和总生物量均表现出较大的数值差异,其中短命植物、一年生植物、类短命植物地上生物量的中值分别为0.441、0617和0.649 g;类短命植物地下生物量中值为1.622,远大于短命植物(0.063 g)和一年生植物(0.082 g)。植株个体总生物量以类短命植物最大,中值为1.622 g,短命植物和一年生植物个体总生物量差异不大,分别为0.035、0.038 g(表2)。类短命植物根冠比最大,均值为1.676;短命和一年生草本植物的根冠比远小于类短命植物,均值分别为0.157、0.148,且两者差异不显著(图3)。
表23种生活型草本植物的地上、地下与总生物量生物量
Table2Abovegroundbiomass(AGB),belowgroundbiomass(BGB)andtotalbiomassofthreelife-formherbaceousplants
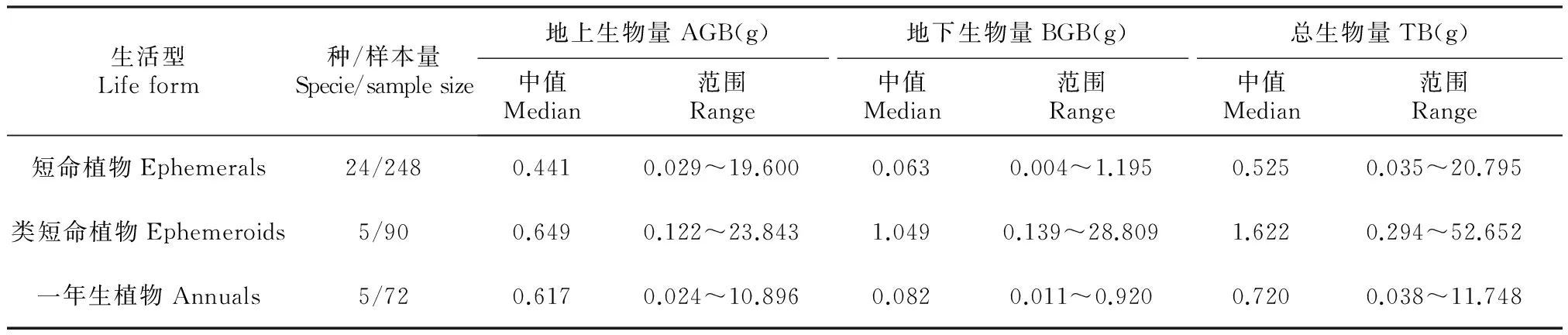
生活型Lifeform种/样本量Specie/samplesize地上生物量AGB(g)地下生物量BGB(g)总生物量TB(g)中值Median范围Range中值Median范围Range中值Median范围Range短命植物Ephemerals24/2480.4410.029~19.6000.0630.004~1.1950.5250.035~20.795类短命植物Ephemeroids5/900.6490.122~23.8431.0490.139~28.8091.6220.294~52.652一年生植物Annuals5/720.6170.024~10.8960.0820.011~0.9200.7200.038~11.748

图3 3种生活型草本植物的根冠比 不同小写字母表示生活型间根冠比在P<0.05水平上差异显著。Fig.3 Root to shoot biomass ratio of three life-form herbaceous plants Different lowercase letter indicates a significant difference P<0.05 among life-form plants.
2.33种生活型草本植物地上与地下生物量的相关生长关系
3种生活型草本植物Root-Shoot间具有极显著(P<0.01)的相关生长关系(图4,表3),且相关生长指数差异显著,其中一年生长营养期草本的相关生长指数最大,其次为短命植物,类短命植物的最小,分别为1.117、1.002和0.949。等速生长检验表明,短命植物Root-Shoot间相关生长指数与1.0差异不显著,属于等速生长;类短命植物Root-Shoot间的相关生长指数显著小于1.0,也即属于地下部分生物量分配速率大于地上部分的异速生长关系;一年生长营养期草本Root-Shoot间的相关生长指数显著大于1.0,即向地上部分分配生物量的速率大于地下部。

图4 3种生活型草本植物根部与地上部生物量间的相关生长关系Fig.4 Allometric relationship between root biomass and leaf biomass of three life-form herbaceous plants
表33种生活型草本植物根部与地上部生物量间的相关生长指数、等速生长及共斜率检验
Table3Allometricrelationship,thetestofallometryandthetestofcommonslopebetweenshootandrootbiomassofthreelife-formherbaceousplants
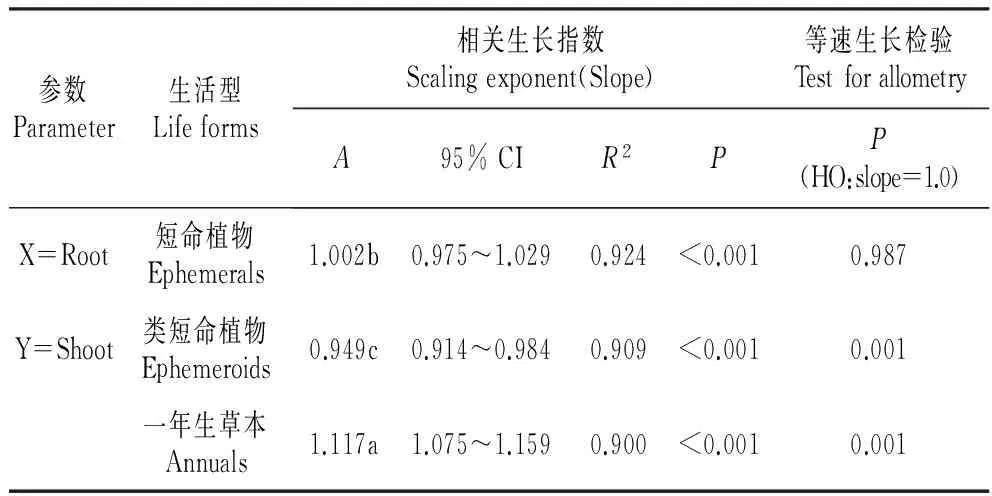
参数Parameter生活型Lifeforms相关生长指数Scalingexponent(Slope)等速生长检验TestforallometryA95%CIR2PP(HO:slope=1.0)X=Root短命植物Ephemerals1.002b0.975~1.0290.924<0.0010.987Y=Shoot类短命植物Ephemeroids0.949c0.914~0.9840.909<0.0010.001一年生草本Annuals1.117a1.075~1.1590.900<0.0010.001
注:相关生长指数(a)一列的不同字母表示物种间a差异显著(P<0.05)。
Note:Different litters indicate significant difference(P<0.05) of allometric scaling exponents among three life-form herbaceous plants.
3 讨论
3.1 不同生活型草本植物的生物量分配特征
生活型是植物对生态因素综合影响下长期适应的结果,是不同物种在相同环境条件下生存所发生的趋同适应,它是生物的一种生态分类单位[23]。植物在长期的自然选择过程中,通过调节自身的资源配置以适应特定的生长环境,形成了对一定环境变化的复杂性和多样性的适应机制。根据最优分配理论,生物量分配使植物的资源限制最小化。因此,在适应生态因素综合影响下,不同生活型植物的生物量分配存在较大差异[20,24]。
短命植物、类短命植物和一年生植物是古尔班通古特沙漠中常见的3种草本植物类群。本研究中3种生活型草本植物个体的地上、地下及总生物量表现出较大的数值差异,但仍可以看出短命和一年生植物向地下分配的生物量远小于类短命植物。进一步分析发现,短命植物和一年生植物的根冠比较小,大多小于0.25,主要集中在0.15左右。这与陶冶等,艾沙江等前人的研究结果类似;陶冶等对准噶尔荒漠6种类短命植物生物量分配特征的研究表明:在开花末期6种类短命植物的根冠比在0.36~3.07[20]。艾沙江等对天山北麓类短命植物新疆郁金香(Tulipasinkiangensis)生物量分配的研究表明:在果实成熟期新疆郁金香的根冠比约为1.0[19]。大部分短命植物和一年生草本的地下生物量分配比例很小,根冠比常在0.50以下[17~18]。类短命植物的根冠比远大于短命和一年生植物,一般大于0.65,集中分布在1.67周围。
由此可见,与短命植物和一年生草本相比,类短命植物向根系投入较大比例的生物量,这样能为植株存储足够的有机质,以保证来年的再次萌发,体现了类短命植物地上部分短生、地下部分多年生的生存策略。短命植物和一年生草本均为当年完成生活史,不需要在地下根系存储来年萌发所需的能量物质,因而两者的根冠比低于类短命植物。另外,短命植物属干旱逃避型植物,而一年生长营养期草本为旱生类型,同时,两者具有不同的萌发和生长时期。尽管如此,短命植物与一年生草本却具有相似的地上与地下生物量分配比例。这表明短命植物与一年生草本植物具有相似或相同的生物量分配特征,体现了2种生活型类群在生物量分配上的趋同适应。
一般认为,高的根生物量分配比例是荒漠演替过程中植物响应地下资源亏缺的普遍特征[25]。将较多的生物量分配到地上部的支撑、生长和繁殖构件,从而减少地下根系的投入是保证植物进行有性繁殖和种群扩散的重要途径[18]。短命植物为避开恶劣的干旱期,因此大部分短命和一年生长营养期草本植物的地上生物量分配比例较高(在80%以上),以便在短时间内资源(主要是水)较为丰富的时期完成生活史。而类短命植物的地上生物量分配比例则较小,其生长过程中常遭遇资源贫乏的时期,面临较大的环境压力。因此需要通过调整形态特征(如叶面积、根长等)来适应环境的变化[26~27]。
3.2不同生活型草本植物地上与地下生物量间的相关生长关系
本研究中,仅短命植物地上与地下生物量间呈等速生长关系,类短命植物和一年生长营养期草本植物地上与地下生物量间均为异速生长关系,其中类短命植物地下与地上生物量间的相关生长指数均显著小于1.0,一年生长营养期草本植物地下与地上生物量间的相关生长指数均显著大于1.0。这个结果不同于通常相关生长关系理论认为的在自然条件下,种内和种间植物个体水平上的地上与地下生物量间具有相同的分配速率(幂指数为1.0,属于等速生长),且这种生长关系不随植物种类(裸子植物或被子植物)和环境的变化而改变[10]。而与Wang等在个体水平上对中国草地地上与地下生物量分配格局的研究结论相同。这说明在干旱区水资源的多寡是决定植物生物量分配的关键性因素,是植物适应干旱环境的一种特殊形态。
通常认为,植物地上、地下生物量间的相关生长关系与物种种类、生活型无关[9~10,20,28]。然而,本研究的结果与这一结论并不完全相同。本研究中类短命植物(a<1.0,P<0.01)和一年生长营养期草本(a>1.0,P<0.01)分别呈现个体越大地下生物量分配比例越大且越少的格局。而短命植物地下与地上生物量间的分配速率与个体大小无关(等速生长)。这可能与物种间系统分类地位有关,即植物在适应温带荒漠气候环境过程中,形成了不同的资源分配策略(如地上部分每年枯死,根、茎结构和功能多样性等),而且这可能是由植物自身某些遗传因素所决定。具体的原因还有待进一步研究证实。
4 结论
短命植物与一年生长营养期草本地下与地上生物量间具有相似的分配特征,但两者地下与地上部生物量间的相关生长关系存在显著差异。类短命植物向根系投入较大比例的生物量,并呈现个体越大地下生物量分配比例越大的趋势。总的来说,3种生活型草本植物呈现的生物量分配特点与其生活史特征相一致,体现了植物经过长期进化形成了与环境相适应的生物量分配特征。
1.杨冬梅,毛林灿,彭国权.常绿和落叶阔叶木本植物小枝内生物量分配关系研究:异速生长分析[J].植物研究,2011,31(4):472-477.
Yang D M,Mao L C,Peng G Q.Within-twig biomass allocation in evergreen and deciduous broad-leaved species:allometric scaling analyses[J].Bulletin of Botanical Research,2011,31(4):472-477.
2.Selzer L J,Lencinas M V,Martínez-Pastur G J,et al.Light and soil moisture effects on biomass and its allocation inOsmorhizadepauperataPhilippi(Apiaceae)[J].Ecological Research,2013,28(3):469-480.
3.陶冶,张元明.3种荒漠植物群落物种组成与丰富度的季节变化及地上生物量特征[J].草业学报,2011,20(6):1-11.
Tao Y,Zhang Y M.Seasonal changes in species composition,richness and the aboveground biomass of three community types in gurbantunggut desert,Northwestern China[J].Acta Prataculturae Sinica,2011,20(6):1-11.
4.Poorter H,Nagel O.The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water:a quantitative review[J].Functional Plant Biology,2000,27(6):595-607.
5.Mokany K,Raison R J,Prokushkin A S.Critical analysis of root:shoot ratios in terrestrial biomes[J].Global Change Biology,2006,12(1):84-96.
6.Wang L,Niu K C,Yang Y H,et al.Patterns of above- and belowground biomass allocation in China’s grasslands:evidence from individual-level observations[J].Science China Life Sciences,2010,53(7):851-857.
7.Poorter H,Sack L.Pitfalls and possibilities in the analysis of biomass allocation patterns in plants[J].Frontiers in Plant Science,2012,3:259.
8.Wang L M,Li L H,Chen X,et al.Biomass allocation patterns across China’s terrestrial biomes[J].PLoS One,2014,9(4):e93566.
9.程栋梁.植物生物量分配模式与生长速率的相关规律研究[D].兰州:兰州大学,2007.
Cheng D L.Plant allometric study of biomass allocation pattern and biomass production rates[D].Lanzhou:Lanzhou University,2007.
10.Enquist B J,Niklas K J.Global allocation rules for patterns of biomass partitioning in seed plants[J].Science,2002,295(5559):1517-1520.
11.张立运,陈昌笃.论古尔班通古特沙漠植物多样性的一般特点[J].生态学报,2002,22(11):1923-1932.
Zhang L Y,Chen C D.On the general characteristics of plant diversity of Gurbantunggut sandy desert[J].Acta Ecologica Sinica,2002,22(11):1923-1932.
12.兰海燕,张富春.新疆早春短命植物适应荒漠环境的机理研究进展[J].西北植物学报,2008,28(7):1478-1485.
Lan H Y,Zhang F C.Reviews on special mechanisms of adaptability of early-spring ephemeral plants to desert habitats in Xinjiang[J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(7):1478-1485.
13.Mckenna M F,Houle G.Why are annual plants rarely spring ephemerals?[J].New Phytologist,2000,148(2):295-302.
14.王雪芹,蒋进,雷加强,等.短命植物分布与沙垄表层土壤水分的关系—以古尔班通古特沙漠为例[J].应用生态学报,2004,15(4):556-560.
Wang X Q,Jiang J,Lei J Q,et al.Relationship between ephemeral plants distribution and soil moisture on longitudinal dune surface in Gurbantonggut desert[J].Chinese Journal of Applied Ecology,2004,15(4):556-560.
15.Warren M,Zou X M.Seasonal nitrogen retention in temperate hardwood forests:the “Vernal Dam” hypothesis and case studies[J].Acta Phytoecological Sinica,2002,27(1):11-15.
16.袁素芬,唐海萍.短命植物生理生态特性对生境的适应性研究进展[J].草业学报,2010,19(1):240-247.
Yuan S F,Tang H P.Research advances in the eco-physiological characteristics of ephemerals adaptation to habitats[J].Acta Prataculturae Sinica,2010,19(1):240-247.
17.邱娟,谭敦炎,樊大勇.准噶尔荒漠早春短命植物的光合特性及生物量分配特点[J].植物生态学报,2007,31(5):883-891.
Qiu J,Tan D Y,Fan D Y.Characteristics of photosynthesis and biomass allocation of spring ephemerals in the Junggar desert[J].Journal of Plant Ecology,2007,31(5):883-891.
18.李雪华,李晓兰,蒋德明,等.科尔沁沙地70种草本植物个体和构件生物量比较研究[J].干旱区研究,2009,26(2):200-205.
Li X H,Li X L,Jiang D M,et al.A comparative study of the individual biomass and modular biomass of 70 herbaceous species found in the Horqin sandy land[J].Arid Zone Research,2009,26(2):200-205.
19.艾沙江·阿不都沙拉木,谭敦炎,吾买尔夏提·塔汉.新疆郁金香营养生长、个体大小和开花次序对繁殖分配的影响[J].生物多样性,2012,20(3):391-399.
Abdusalam A,Tan D Y,Tahan O.Effects of vegetative growth,plant size and flowering order on sexual reproduction allocation ofTulipasinkiangensis[J].Biodiversity Science,2012,20(3):391-399.
20.陶冶,张元明.准噶尔荒漠6种类短命植物生物量分配与异速生长关系[J].草业学报,2014,23(2):38-48.
Tao Y,Zhang Y M.Biomass allocation patterns and allometric relationships of six ephemeroid species in Junggar Basin,China[J].Acta Prataculturae Sinica,2014,23(2):38-48.
21.刘乾.古尔班通古特沙漠南缘植物群落多样性与植被覆盖变化分析[D].乌鲁木齐:新疆农业大学,2009.
Liu Q.Study on the plant community diversity and vegetation coverage changes in the south of Gurbantunggut desert[D].Urumchi:Xinjiang Agricultural University,2009.
22.Ravindranath N H,Ostwald M.Methods for below-ground biomass[M].//Ravindranath N H,Ostwald M.Carbon Inventory Methods Handbook for Greenhouse Gas Inventory,Carbon Mitigation and Roundwood Production Projects.Netherlands:Springer,2007:149-156.
23.颜忠诚.生态型与生活型[J].生物学通报,2001,36(5):4-5.
Yan Z C.Ecological and life style[J].Bulletin of Biology,2001,36(5):4-5.
24.Zhou X B,Zhang Y M,Niklas K J.Sensitivity of growth and biomass allocation patterns to increasing nitrogen:a comparison between ephemerals and annuals in the Gurbantunggut Desert,north-western China[J].Annals of Botany,2014,113(3):501-511.
25.Gill R A,Kelly R H,Parton W J,et al.Using simple environmental variables to estimate below-ground productivity in grasslands[J].Global Ecology and Biogeography,2002,11(1):79-86.
26.Poorter H,Niklas K J,Reich P B,et al.Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control[J].New Phytologist,2012,193(1):30-50.
27.李志波,李平衡,王权,等.梭梭和多枝柽柳的末端小枝异速生长特征研究[J].植物研究,2013,33(3):274-281.
Li Z B,Li P H,Wang Q,et al.Allometric characteristics of terminal twigs ofTamarixramosissimaandHaloxylonammodendron[J].Bulletin of Botanical Research,2013,33(3):274-281.
28.Niklas K J.Plant allometry:is there a grand unifying theory?[J].Biological Reviews,2004,79(4):871-889.
introduction:ZHAO Jing(1988—),male,master student,the main research direction is ecological planning and management.
date:2016-11-14
AllocationPatternsandAllometricRelationshipsbetweenAbove-andBelow-groundBiomassofThreeLife-formHerbaceousPlantsintheGurbantünggütDesert
ZHAO Jing WANG Xi-Yuan*
(Key Laboratory of Oasis Ecology,College of Resources and Environment Science,Xinjiang University,Urumqi 830046)
We took biomass survey samples on herbaceous plants along the southern margin Gurbantünggüt desert to study the biomass allocation features of herbaceous plants of different life-forms in Gurbantünggüt Desert. Results showed that: (1)most of the total biomass of the sampled 34 species of herbaceous plants was less than 4 g, most of whose above-ground biomass(AGB) and below-ground biomass(BGB) was less than 2 and 0.5 g, respectively, and their root-shoot ratios(R/S) were less than 0.25; (2)Ephemeroids allocated more biomass to below ground parts and had a higher R/S, which distributed around 1.67 intensively; Ephemerals and annuals allocated much less biomass to belowground parts, both of which has smaller R/S around 0.15; (3)The growth speed of the biomass of up-ground and below-ground parts of ephemeral parts was equal, but that of the ephemeroids was unequal. The proportion of the ephemeroids biomass allocated to the below-ground parts was increased complying with the individual’s enlargement, while annuals biomass was on contrast. Therefore, the biomass of up-ground and below-ground parts of ephemeral, ephemeroids and annuals has different allocation features, which is consistent with the special plant life story features.
Gurbantünggüt Desert;herbaceous plants;life-form;biomass allocation
赵晶(1988—),男,硕士研究生,主要研究方向为生态规划与管理。
* 通信作者:E-mail:1076135474@qq.com
2016-11-14
* Corresponding author:E-mail:1076135474@qq.com
S567.2
A
10.7525/j.issn.1673-5102.2017.02.020
