世界蓑藓属植物分布式样及其系统演化和生物地理学意义
2017-11-09郭水良吴倩倩
郭水良 吴倩倩 于 晶 曹 同
(上海师范大学生命与环境科学学院,上海 200234)
世界蓑藓属植物分布式样及其系统演化和生物地理学意义
郭水良 吴倩倩 于 晶 曹 同*
(上海师范大学生命与环境科学学院,上海 200234)
蓑藓属(Macromitrium)是苔藓植物分类、生态学研究及生物多样性保护的重要类群。以“IndexMuscorum”中的14个藓类植物地理分布单位为分析区域,基于标本信息和相关文献,在分类修订的基础上,统计了世界蓑藓属274种植物在14个地理单位的分布数据,基于这些数据,应用聚类分析和除趋势对应分析法,分别得到了14个地理单位的聚类图和三维空间图,直观展示了世界蓑藓属植物的地理分布格局。发现蓑藓属植物区系在东亚(As2)、大洋洲(Oc)、南亚和东南亚(As3,As4)、非洲(Afr2,Afr3,Afr4)、中美洲—南美洲(Am2,Am3,Am4,Am5)、澳洲(Austr1,Austr2)存在区域分化。热带亚洲和热带美洲是世界蓑藓属植物的多样化中心。除长柄蓑藓(M.microstomum(Hook. & Grev.) Schwägr.)在14个地理单位均有分布外,大部分种类分布限于少数或仅一个地理单位。蓑藓属植物在东亚、热带亚洲、热带美洲和澳洲不仅种类存在明显分化,而且形态性状也有区域特点。根据蓑藓属植物的现代分布式样,探讨了其系统演化和生物地理学意义。
蓑藓属;地理分布;聚类分析;排序分析
蓑藓属(MacromitriumBridel)隶属于苔藓植物门(Bryophyta)藓纲(Bryopsida)木灵藓目(Orthotrichales)木灵藓科(Orthotrichaceae),是木灵藓科中的最大属。该属植物先后记录有946个种名,Crosby等报道有368种,是藓类植物中种数仅次于凤尾藓属(Fissidens460种)和真藓属(Bryum449种)的第三个大属[1]。
蓑藓属植物主要分布于热带和泛热带地区[2]。目前世界大部分地区已经完成了蓑藓属的分类修订,这些地区包括新西兰[3]、澳大利亚[4]、巴布亚新几内亚的Huon地区[5]、Malesia地区(包括热带东南亚和大洋洲的新几内亚)[6]、墨西哥[7]、中美洲[8]、南部非洲[9]、日本[10]和印度东部及邻近地区[11]。郭水良等在国家自然基金委的资助下,先后开展亚洲、非洲和南美洲地区的蓑藓属分类修订工作[12~23];近年来,Wilbram等[24]先后开展了非洲地区的蓑藓属植物分类修订。根据前人研究资料,以及本文作者对蓑藓属植物的分类修订,目前比较明确的蓑藓属植物有274种。
初步统计IndexMuscorum蓑藓属植物在世界藓类植物地理单位中的分布情况,发现绝大部分种类记录于一个或少数几个地理单位[2]。蓑藓属植物在世界范围的地理分布特点并不清楚,研究该属的地理分布式样将有利于更好地开展该属的分类与系统学研究。
1 研究方法
根据IndexMuscorum,世界藓类植物分布被划分为20个地理单元[2],分别是Am.1-Am.6,Eur.,As.1-As.5,Afr.1-Afr.4,Austr.1,Austr.2,Ant and Oc.(图1)。蓑藓属是泛热带分布类群,分布于As.2-As.4,Afr.2-Afr.4,Austr.1,Austr.2,Oc.,Am.1-Am.6共计15个地理单位。由于Am1仅有MacromitriumrichardiiSchwägr.一种分布于该区域最南缘的佛罗里达州(Florida,近Am3),所以本文的地理单位分析中不包括Am.1。
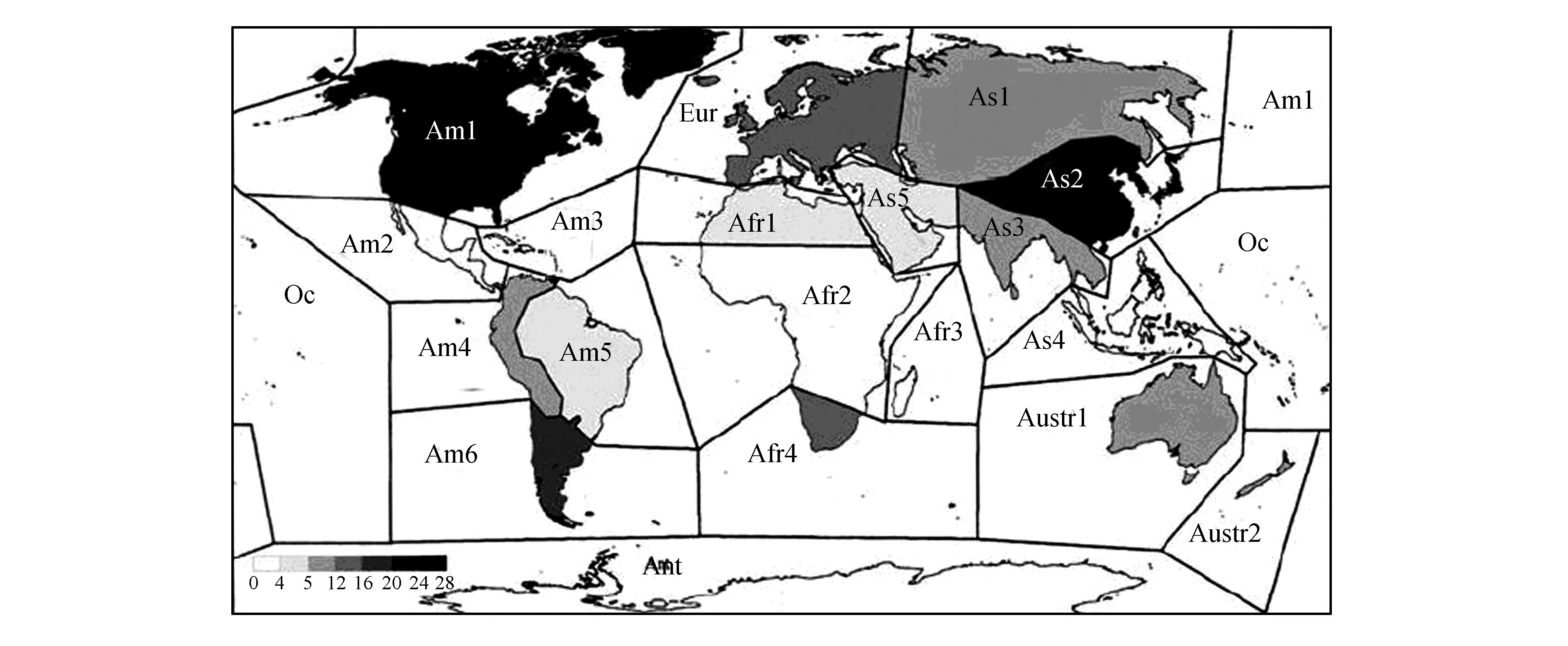
图1 世界藓类植物地理分布的20个地理单位Fig.1 Twenty geographical units in relation to global geographical distribution of mosses
根据文献资料,结合标本记录,获得蓑藓属274种植物在世界14个地理单元有(赋值1)、无(赋值0)分布信息,建立0、1数据矩阵(由于篇幅有限,数据存查)。应用Pcord 5.0进行聚类分析,选择Jaccard系数比较各地区蓑藓植物区系相似性,应用离差平方和法作为聚类策略,获得反映14个地理单位在蓑藓属植物组成相似性的聚类树状图[25],初步了解世界蓑藓属植物在14个地理单位的种类分化情况;同时应用该软件中的除趋势对应分析(Detrended Correspondence Analysis,DCA)分析模块,将“number of segments”设置成26,Rescaling threshold设置成0,获得反映14个地理单元蓑藓属植物分布的三维排序信息,再应用SPSS16.0对蓑藓属植物在14个地理单元的分布特点进行了三维图形展示。根据三维图上展示的各地理单元间的空间距离来探讨世界蓑藓属植物的地理区系关系。
2 结果与分析
统计蓑藓属植物在世界14个地理单元的分布数据,发现热带亚洲(As4)和热带美洲(Am4)种数最多,分别为48和47种,其次是热带美洲以亚马逊流域为主(Am5)的热带南美洲、热带大洋洲(Oc)和南亚(As3),种数分别为39、38和34;再次为中美洲Am2、东亚(As2)、澳洲的Austr1,马达加斯加及印度洋岛屿(Af3),种数分别为29、26、26和21,Austr2、Af2、Am6、Am3的种数均在13~16,另外,南部非洲(Af2)有7种。剩余的其他地理单元中,北美大陆仅M.richardii一种分布于Am1最南缘的佛罗里达洲外,而南极大陆(Ant)、欧亚大陆的大部分地区(Eur、As1)、中东(As5)和北非(Afr1)这五个地理单位没有蓑藓分布。全球范围内,蓑藓属植物在南半球占绝对优势。
对14个地理单位(Am1除外)的蓑藓属植物分布式样进行了聚类和排序分析,得到图2和3。图2中虚线处为划分阈值下,世界蓑藓属植物可以分成六个区域。

图2 基于蓑藓属植物分布数据的14个地理单位聚类树状图Fig.2 Clustering dendrogram of fourteen geographical units based on their species distribution of Macromitrium

图3 蓑藓属植物在世界14个地理单位分布式样的三维排序图Fig.3 Three-dimensional ordination plot revealing geographical distribution pattern of Macromitrium in fourteen geographical units 注:1=As.2,2=As.3,3=As.4,4=Af.2,5=Af.3,6=Af.4,7=Oc,8=Austr.1,9=Austr.2,10=Am.2,11=Am.3,12=Am.4,13=Am.5,14=Am.6
区域1:包括东亚(As2),该地区包括26种,代表性的种类有钝叶蓑藓(M.japonicum)、中华蓑藓(M.cavaleriei)、M.clastophyllum、M.comatum、M.formosae、M.gymnostomum、M.hainanese、M.holomitrioides、M.prolongatum、M.rhacomitrioides、M.subincurvum、M.taiheizanense、M.taiwanense、M.tosae、M.tuberculatum和M.uraiense,这16种仅限于本区域,占本区域总种数的61.54%,孢蒴多数为卵状椭圆形,蒴齿退化或单层。
区域2:包括南亚(As3)和东南亚(As4)两个地理单位,共记录有82种,其中长柄蓑藓(M.microstomum)、尼泊尔蓑藓(M.nepalense)、M.falcatulum、M.incurvifolium、M.lauterbachii、M.leptocarpum、M.minutum、南亚蓑藓(M.sulcatum)同时记录于这两个区域,其中尼泊尔蓑藓叶片细胞疣特点,枝叶表面由不规则的1~3层出芽状突起的细胞,这些细胞的背腹两面又有多疣;南亚蓑藓蒴帽大无毛,蒴齿两层等特点明显,有别于东亚蓑藓的种类。本组中64种仅记录于本区域中,占总种数的78.05%。
区域3:包括大洋洲(Oc)一个地理单位,共有38种,其中有30种仅记录于本区域,占种数的78.95%。代表性的种类有M.altum、M.bistratosum、M.brevisetum、M.cardotii、M.emersulum、M.francii、M.neo-caledonicum等。
区域4:包括Afr.2(中非)、Afr.3(马达加斯加岛)、Afr.4(南非)三个地理单元,本区共有蓑藓属植物36种,占世界蓑藓属植物总种数的13.1%。其中27种为该地区特有种,占该组总种数的75%。Afr.2单元记录的14种中9种为该区域特有,占该地区蓑藓属种数的64.3%;Afr.3记录的21种中14种为特有种,占该地区总种数的66.7%。Afr.2、Afr.3和Afr.4地处古热带植物区,蓑藓属植物区系成分相似性也较大。该区域内特有种Macromitriumlevatum和M.serpens在Afr.2、Afr.3和Afr.4三个单元内均有分布,该种的配子体叶片特征类似于M.nepalense,As4的M.tongense也有类似的特点,一方面反映了区域4内各地理单位之间的亲缘关系,也反映出与As3、As4的亲缘关系。
区域5:包括Austr.1(澳大利亚)、Austr.2(新西兰)和Am.6(智利—巴塔哥尼亚)3个地理单元,本区共有蓑藓属植物47种,占世界蓑藓属植物总种数的17.1%,其中该地区特有种33种,占世界本组总种数的70.2%,其中Austr.1单元特有种16种,Austr.2单元特有种9种。其中M.ligulare、M.longipes和M.longirostre为这三个地理单元共有,也是组5的特有种。
区域6:包括南美—中美洲的Am.2(墨西哥)、Am.3(加勒比地区)、Am.4(秘鲁)、Am.5(巴西)共四个地理单位。Am.2和Am.3属于加勒比植物地区,Am.4属于安第斯植物地区,Am.5属于巴西植物地区,这几个区都是新热带植物地区。本组共有蓑藓属植物90种,占世界蓑藓属植物总种数的32.85%,其中为本组所特有的有80种,占本组总种数的88.89%。其中M.cirrosum、M.longifolium、M.punctatum、M.swainsonii既是该组的特有种,也为四个地理单位所共有,另外,M.richardii在这4个单元内也均有分布,组外记录的仅为Am.3相邻的佛罗里达半岛。因此,这五种是本组的代表性区系成分。从聚类图上可以看出,本区域中,位于中美洲的墨西哥(Am.2)和加勒比地区(Am.3)具有相近的区系成分,两地区分布的34种藓类植物中,有9种共有;南美洲两个地理单位(Am4,5)共计有70种蓑藓植物,其中15种共有。
从三维排序图3中看出,大洋洲Oc(7)位于排序图的外围,反映出两者蓑藓属区域特色明显。
274种蓑藓植物中,分布最广的长柄蓑藓,出现在14个地理单位,其次是M.richardii,记录于Am1-5六个地理单位,再次是M.cirrosum、M.guatemalense、M.punctatum、M.swainsonii、M.longifolium、M.orthostichum、M.incurvifolium和M.involutifolium共计8个种分布于4个地理单位,其中前5个种见于热带美洲,M.orthostichum见于热带美洲、大洋洲、热带亚洲和马达加斯加。M.aureum、M.fuscescens等10种蓑藓记录于3个地理单位,M.acutirameum和M.angulatum等37种蓑藓记录于2个地理单位,而余下的217种蓑藓植物分别均报道自一个地理单位。图4示意性地反映了274种蓑藓属植物的地理分布特点,也反映出众多的种类仅见于一个地理单位,热带亚洲(As4)和大洋洲Oc(7)具有最多的地区特有种。地理单位数(x)与种数(y)的关系为:y=121.28*e-1.721 6*x(r=0.999,n=6,P<0.001)(图5),即绝大部分种类仅记录于少数地区。

图4 反映蓑藓属植物地理分布的聚类图Fig.4 Clustering dendrogram of 274 species of Macromitrium revealing their distribution pattern

图5 蓑藓植物种数与地理单位数关系Fig.5 Relationship between species number of Macromitrium and geographical units
3 讨论
世界蓑藓属植物在不同地理单位的特有和分化现象明显。亚洲地区的三个地理单位(As.2-4)共记录有90种蓑藓属植物,美洲的六个地理单位(Am.1-6)共记录有99种蓑藓,仅长柄蓑藓和M.orthostichum二种同时也见于美洲,两者区系相似性(Jaccard系数)仅为0.01,亚洲的90种蓑藓属植物与非洲的36种蓑藓中,也仅有M.orthostichum,M.microstomum和M.mauritianum3种相同,区系相似系数也仅为0.05。
在地质历史上,本文的As.2处于劳亚古陆,而南美洲(Am.2-4)、非洲(AF.2-4)、澳洲(Austr.1-2)、南亚次大陆(As.3)是冈瓦纳古陆(Gondwanaland)。在蓑藓属植物区系上,东亚(As.2)的25种蓑藓属植物与美洲的99种蓑藓属植物中,仅长柄蓑藓一种共有,区系相似系数仅为0.008。而且,长柄蓑藓在东亚也主要记录于该地理单位的东南边缘范围(中国的海南岛、台湾岛和云南的西双版纳),并非典型的东亚成分。因此,蓑藓属植物东亚成分的区系强烈极为分化。因此推测蓑藓属植物可能起源于冈瓦纳古陆与劳亚古陆分离之前,两者独立演化形成了目前的区系。
在地质上,南亚次大陆(As.3)曾是冈瓦纳古陆的一部分。在地理区系上,As.3属于南半球区。在三维散点图上,As.3的位置处于As.2与Af.3(及Af.4)之间。这可能是因为As.3与冈瓦纳古陆分离后,在新生代的始新世时期和亚洲撞合,形成南亚次大陆的一部分,并与As.2相邻,受到了物种迁移的影响。如As.2中分布的种类在As.3也均有分布,而非洲板块的种类也有部分见于南亚(M.microstomum,M.sulcatum,M.sulcatumvar.ceylanium)。
地理分布特点的分析也给出了系统演化上的信息。根据分支系统学的性状极性判别的生物地理学标准,生物的扩散往往由近到远,因此分布广泛的种类往往是起源较早的类群。长柄蓑藓是记录于除北美(Am1)外的其余14个地理单位,这种地理分布特点暗示该种在起源上具有原始性。长柄蓑藓的配子体叶片细胞光滑、上部细胞近圆形,中下部细胞长方形或长条形,叶椭圆状铍针形,全缘;孢蒴椭圆形、有蒴齿;蒴帽和蒴柄光滑;孢子同型。因此,这些性状是原始的。相对于这些性状,以下性状是进化的:叶片细胞具疣(例如钝叶蓑藓、大帽蓑藓M.tosae、贵州蓑藓M.fortunati等)、枝叶表面由不规则的1-3-层出芽状突起的细胞,这些细胞的背腹两面又有多疣(尼泊尔蓑藓、M.serpens,M.tongense)、叶先端边缘细啮齿状(M.rhacomitrioides、M.fulgescens、M.sejunctum)、叶基边缘细胞有大形齿状细胞(M.carionis、M.fragilicuspis、M.guatemalense、M.oblongum等)、孢蒴形态特殊(M.sulcatum)、蒴齿退化(海南蓑藓、M.formosae,M.gymnostomum,M.heterodictyon,M.holomitrioides,M.robinsonii,M.schmidiiandM.taiheizanenseNog.),蒴帽有毛、蒴柄粗糙或有疣(M.orthostichum、M.longifolium等)。配子体叶片形态特殊的一些类群也是进化或特化的,例如分布于南亚的M.densum(叶形短舌状,先端强烈兜形),或者分布于南美的M.standleyi(叶形特别细长,长达10 mm),或似石松状反折的叶(M.perreflexum)、具双层叶片细胞的种类(M.bistratosum、M.gracile),或者叶先端具长突尖(M.blumei)等。
Shaw曾指出,相对于其他高等植物,众多的藓类植物应该是跨洲际分布的[26]。但是,作为藓类植物中的第三大属,有高达79.2%的种类仅局限于某一地理单位,有93.43%的种限于某一洲的范围,跨洲际分布的仅占6.57%,仅长柄蓑藓和M.orthostichum跨三个以上洲。这一方面与蓑藓属植物绝大多数为树附生植物,与植被类型等密切有关,影响了该属植物的广泛分布。还有一种情况是,目前有关蓑藓属植物的分类修订多数限于某一国家,或者限于某个洲有关,或者目前该属的分类学家可能采纳了“小种”的概念。如果开展全球范围进一步开展分类修订,有可能发现新的异名,从而增加该属植物洲际分布的类群。
有关植物的现代地理分布格局,有替代分布学说和扩散学说。从蓑藓属植物的地理分布式样来分析,位于起源于劳亚古陆的东亚(As.2)的蓑藓植物与起源于冈瓦纳古陆的南亚(As.3)、澳洲(Austr1-2)、非洲(Af.2-4)、南美洲(Am.4-6)的蓑藓属植物种类构成明显不同,从大陆漂移的角度,也能够说蓑藓属植物的现代分布式样,即蓑藓属植物在大陆漂移前已经起源,最早产生的种类配子体叶片细胞光滑、蒴柄和蒴帽光滑、有蒴齿的种类(例如长柄蓑藓)及其近缘种,然后在不同的板块上分别独立演化,形成了现代各大板块种类构成明显不同的类群。
如果按扩散学说,按全球大气环流的方向,蓑藓属植物在南半球与北半球会形成种类构成明显不同的区系特点。但是,在南半球的各大板块,应该有相似度较高的种类组成。但是,非洲的36种蓑藓属植物与南美洲的99种蓑藓属植物中,也仅M.orthostichum、M.microstomum、M.urceolatum和M.acutirameum4种相似,区系相似系数(Jaccard系数)仅为0.03,南亚块板(As.3)与非洲板块(Af2-4)的区系也仅M.sulcatum,M.sulcatumvar.ceylanium和M.microstomum相似,区系的相似系数也仅为0.05。这种洲际间的低比例区系相似性,用替代学说或许更能够好地进行解释。例如,澳洲与南美洲曾一直连接到第三世纪中叶,随着大陆的分裂和漂移才逐渐分开,这为M.gracile、M.microstomum、M.ligulare、M.longipes和M.longirostre在南美洲和澳洲的间断分布提供了现代分布的依据。
1.Crosby M R,Magill R E,Allen B,et al.A checklist of the mosses[M].St.Louis:Missouri Botanical Garden,1999.
2.Van Der Wijk R,Margandant W D,Florschüetz P A.Index muscorum,vol.Ⅲ[M].Utrecht:International Bureau for Plant Taxonomy and Nomenclature,1967.
3.Vitt D H.The New Zealand species of the pantropical genusMacromitrium(Orthotrichaceae:Musci):taxonomy,phylogeny and phytogeography[J].Journal of the Hattori Botanical Laboratory,1983,54:1-94.
4.Vitt D H,Ramsay H P.TheMacromitriumcomplex in Australasia(Orthotrichaceae:Bryopsida).Part Ⅰ.Taxonomy and phylogenetic relationships[J].Journal of the Hattori Botanical Laboratory,1985,59:325-451.
5.Vitt D H,Koponen T,Norris D H.Bryophyte flora of the Huon Peninsula,Papua New Guinea.LV.Desmotheca,Groutiella,MacrocomaandMacromitrium(Orthotrichaceae,Musci)[J].Acta Botanica Fennica,1995,154:1-94.
6.Eddy A.A handbook of Malesian Mosses:splachnobryaceae to leptostomataceaev.3[M].London:Natural History Museum Publications,1996:1-277.
7.Vitt D H.Orthotrichaceae[M].//Sharp A J,Crum H,Eckel P M.The moss flora of Mexico.New York:Memoirs of the New York Botanical Garden,1994,69:590-656.
8.Allen B.Moss flora of central America.Part 2:Encalyptaceae-Orthotrichaceae[M].St Louis,Mo.:Missouri Botanical Garden,2002.
9.Magill R E,Van Rooy J.Bryophyta.Part 1 musci,fascicle 3,Erpodiaceae-Hookeriaceae[M]//Leistner O A.Flora of southern Africa[M].Pretoria:National Botanical Institute,1998:462-475.
10.Noguchi A.Musci japonici.Ⅶ.The genusMacromitrium[J].Journal of the Hattori Botanical Laboratory,1967,30:207-230.
11.Gangulee H C.Mosses of Eastern India and adjacent regions Fascicle 5[M].Calcutta,India(Foreign distributor:Otto Koeltz,Koenigstein-Taunus,Germany),1976:1135-1462.
12.Guo S L,He S.A new species ofMacromitrium(Orthotrichaceae) from Hainan,China[J].The Bryologist,2008,111(3):505-509.
13.Guo S L,Cao T,Tan B C.Three new species records of Orthotrichaceae (Bryopsida)in China,with comments on their type specimens[J].Cryptogamie Bryologie,2007,28(2):149-158.
14.Guo S L,Cao T,Tan B C,et al.Taxonomic notes on Asian species of Orthotrichaceae(Bryopsida):Macromitriumwith gymnostomous capsules[J].Garden's Bulletin Singapore,2007,58(2):155-178.
15.Guo S L,Enroth J,Koponen T.Bryophyte flora of Hunan province,China.11.Orthotrichaceae (Music)[J].Annual Botanica Fennici,2007,44:1-34.
16.Guo S L,Tan B C,Virtanen V.Taxonomic and morphometric comments onMacromitriumblumei,M.zollingeriandM.annamense(Orthotrichaceae,Bryophyta)[J].Nova Hedwigia,2006,82:(3-4):467-482.
17.Guo S L,He S.Macromitriumcavaleriei,a little known moss from China with four new synonyms(Musci:Orthotrichaceae)[J].Journal of Bryology,2008,30(4):264-270.
18.Guo S L,Ma Y H,Cao T,et al.A synopsis ofMacromitrium(Orthotrichaceae) in China[J].Cryptogamie,Bryologie,2012,33(4):341-355.
19.Yu J,Yong K T,Guo S L.Taxonomic notes onMacromitriumdensum(Bryopsida) with a new synonym,M.brevissimum[J].Nordic Journal of Botany,2014,32(4):437-440.
20.Yu J,Guo S L,Ma Y H,et al.Macromitriumousiense,a neglected Chinese moss species(Orthotrichaceae,Bryopsida) with new synonym and records[J].Nordic Journal of Botany,2013,31(3):339-343.
21.Yu J,Guo S L,Ma Y H,et al.Taxonomic and morphometric comments onMacromitriumtosaeBesch.(Orthotrichaceae),with its four new synonyms[J].The Bryologist,2012,115(3):388-401.
22.Guo S L,Yu J,Ma Y H.Typification of an endangered mossMacromitriumfortunatii(Orthotrichaceae)[J].Chenia,2013,11:95-101.
23.Lou Y X,He S,Guo S L.Using macroclimatic models to estimate the distribution ranges of taxonomically challenging taxa,an example withMacromitriumcavalerieiCardot & Thér.(Orthotrichaceae)[J].Journal of Bryology,2014,36(4):271-278.
24.Wilbraham J.Annotated checklist and keys to the Orthotrichaceae of Malawi,together with new country records for East Africa[J].Journal of Bryology,2015,37(2):87-95.
25.郭水良,于晶,陈国奇.生态学数据分析—方法、程序与软件[M].北京:科学出版社,2015.
Guo S L,Yu J,Chen G Q.Ecological data analysis-methods,programs and software[M].Beijing:Science Press,2015.
26.Shaw J.Biogeographic patterns and cryptic speciation in bryophytes[J].Journal of Biogeography,2001,28(2):253-261.
National Natural Science Foundation of China(31370233,31570208,30970184,30570121)
introduction:GUO Shui-Liang(1964—),male,professor,research on plant systematic and taxonomy,plant ecology.
date:2016-11-08
GeographicalDistributionPatternofMacromitriumintheWorldandItsBiogeographicalSignificance
GUO Shui-Liang WU Qian-Qian YU Jing CAO Tong*
(College of Life and Environmental Sciences,Shanghai Normal University,Shanghai 200234)
The genusMacromitrium(Orthotrichaceae, Bryophyta) is an important taxon in bryological systematics, ecology and biological conservation. On the basis of our taxonomical revision of the genus, we got the distribution data of 274 species of the genus in 14 geographical unites(suggested by “Index Muscorum”) in the world.By using Clustering Analysis and Detrended Correspondence Analysis(DCA), we got a dendrogram and a three-dimensional scatter plots to reveal the distribution pattern of the genus visually. The results showed a distinctive floristic differentiation ofMacromitriumamong As2, Oc, As3-4, Afr2-3-4, Am2-3-4-5, and Austr1-2, with highest diversity in Tropical Asia and Tropical America. ExceptM.microstomum(Hook.& Grev.) Schwägr., all other species were recorded from a single or a few geographical unites. Among different global regions,Macromitriumdifferentiated not only in its floristic elements, but also in their morphological characters. We discussed the systematic and biogeographical significances of the genus according its global distribution pattern.
Macromitrium;geographical distribution pattern;Clustering Analysis;Detrended Correspondence Analysis(DCA)
国家自然科学基金(31370233,31570208,30970184,30570121)
郭水良(1964—),男,教授,主要从事植物系统与分类学、植物生态学。
* 通信作者:E-mail:ct1946@263.net
2016-11-08
* Corresponding author:E-mail:ct1946@263.net
Q949.35+2
A
10.7525/j.issn.1673-5102.2017.02.002
附表

表1 274种蓑藓属植物在15个地理单位的分布情况
续表Continuedtable

种名NamesAs2As3As4Afr2Afr3Afr4OcAustr1Austr2Am1Am2Am3Am4Am5Am6M.eriomitrium000000000000010M.erubescens001000000000000M.evrardii010000000000000M.exsertum000000010000000M.falcatulum011000000000000M.fimbriatum000010000000000M.flavopilosum000000000010000M.formosae100000000000000M.fortunatii101000000000000M.fragile000000000000100M.fragilicuspis000000000010000M.francii000000100000000M.frondosum000000000000100M.frustratum000000000010000M.funicaule000010000000000M.funiforme000000010000000M.fuscescens000000100011000M.fuscoaureum000000000010000M.galipense000000000010110M.glaziovii000000000000010M.globirameum000000100000000M.gracile000000001000010M.gracilipes000000100000000M.greenmanii000000000010000M.grossirete000000001000000M.guatemalense000100000010101M.hainanese100000000000000M.hamatum010000000000000M.helmsii000000001000000M.hemitrichodes000000011000000M.herzogii000000000000100M.hildebrandtii000010000000000M.hoehnei000000000000010M.holomitrioides100000000000000M.homaloblastum000000000000100M.hortoniae000000010000000M.incrustatifolium010000000000000M.incurvifolium011000110000000M.intortifolium000000000000010M.involutifolium100010111000000M.japonicum110000000000000M.kinabaluense001000000000000M.koghiense000000100000000M.krausei000000000000001M.laevigatum000000001000000M.lanceolatum000010000000000M.lauterbachii011000000000000M.lebomboense000001000000000M.leprieurii000000000010000M.leptocarpum011000000000000M.leratii000000100000000M.leratioides000000100000000M.levatum000111000000000M.ligulaefolium000000010000000M.ligulare000000011000001M.liliputanum000100000000000M.lonchomitrioides000000001000000M.longibrachteatum010000000000000M.longicaule001000000000000M.longifolium000000000011110M.longipes000000011000001M.longipilum001000000000000M.longirostre000000011000001M.lorifolium010000000000000M.ludovicae000000100000000M.macrosporum001000000000000M.macrothele000000000000100M.marginatum001000000000000
续表Continuedtable

种名NamesAs2As3As4Afr2Afr3Afr4OcAustr1Austr2Am1Am2Am3Am4Am5Am6M.masafuerae000000000000001M.mauritianum001110000000000M.mcphersonii000000000010100M.megalocladon001000000000000M.megalosporum000100000000000M.melanostomum100000000000000M.melinii000000000000100M.menziesii000000100000000M.microcarpum000000000000001M.microstomum111011010000111M.minutum011000000000000M.moorcroftii110000000000000M.nanothecium000010000000000M.nematosum000000000000010M.neocaledonicum000000100000000M.nepalense111000000000000M.nietneri010000000000000M.nigricans010000000000000M.noguchianum001000000000000M.norrisianum001000000000000M.oblongum000000000000110M.ochraceoides001000000000000M.ochraceum001000000000000M.okabei000000100000000M.onraedtii000010000000000M.orthophyllum000000001000000M.orthostichum001010100000100M.ovale000000000000100M.pallidum000010000000000M.papillisetum001000000000000M.paridis000000100000000M.parvifolium001000000000000M.parvirete000000000010000M.patens000000000010000M.pellucidum000000000000110M.peraristatum000000010000000M.perdensifolium001000000000000M.perfragile000000000000010M.perichaetiale000000000001010M.perpusillum000000000010000M.perreflexum000000000000100M.petelotii001000000000000M.picobonitum000000000010000M.pilicalyx000000100000000M.piliferum000000100000000M.pilosum001000000000000M.plicatum000000100000000M.proliferum000000000000100M.prolongatum100000000000000M.prorepens000000001000000M.proximum000010000000000M.pulchrum000000100000000M.pullenii001000000000000M.punctatum000000000011110M.ramsayae000000100000000M.recurvifolium001000001000000M.refractifolium000000000000100M.regnellii000000000000110M.renauldii000000100000000M.repandum000000010000000M.retusulum000000000010100M.retusum000000001000000M.rhacomitrioides100000000000000M.rhaphidophyllum000000000001100M.richardii000000000111110M.rigbyanum010000000000000M.rimbachii000000000000100M.rugulosum000000000000010M.sanctaemariae000010000000000M.sarasinii000000100000000
续表Continuedtable

种名NamesAs2As3As4Afr2Afr3Afr4OcAustr1Austr2Am1Am2Am3Am4Am5Am6M.savatieri000000100000000M.schiffneri000000000000010M.schmidii010000000000000M.scoparium000000000001010M.sejunctum000000000010000M.semipapillosum000010000000000M.semperi001000000000000M.seriatum000100000000000M.serpens000111000000000M.similirete001000000000000M.soulae000010000000000M.speirostichum000000100000000M.st-johnii000000100000000M.standleyi000000000010000M.standleyivar.subundulatum000000000011000M.stephanodictyon001000000000000M.stoneae000000010000000M.streimannii001000000000000M.striatum001000000000000M.strictfolium000000000000010M.stricticuspis000000000000100M.subbrevihamatum000000000000010M.subcirrhosum000000000010000M.subdiscretum000000000000100M.subhemitrichodes000000110000000M.subincurvum100000000000000M.sublaeve000000000000110M.sublongicaule001000000000000M.submucronifolium000000001000000M.subpaucidens000000000000100M.subperichaetiale000000000001000M.subsessile000000100000000M.substrictifolium000000000000010M.subtortum010000000000000M.subulatum000000010000000M.subvillosum000000100000000M.sulcatum011010000000000M.sulcatumvar.ceylanicum010001000000000M.swainsonii000000000011110M.tahitisecundum000000000000001M.taiheizanense100000000000000M.taiwanense100000000000000M.taoense000000100000000M.thwaitesii010000000000000M.tocaremae000000000000100M.tongense000000110000000M.tortifolium000100000000000M.torulosum010000000000000M.tosae110000000000000M.trachypodium000000000000100M.trichophyllum000000000000100M.trollii000100000000000M.tuberculatum100000000000000M.turgidum010000000000000M.tylostomum001000000000000M.ulophyllum000000000000100M.undatum000000000000010M.uraiense100000000000000M.urceolatulum000010000000000M.urceolatum000100000000001M.venezuelense000000000000100M.vesiculosum010000000000000M.vitianum000000100000000M.viticulosum000000010000000M.wattsii000000010000000M.weissioides000000010000000M.xenizon000000000000010M.yuleanum001000000000000M.zimmermannii001000000000000
