Comparison between Betalain and Flavonol Accumulation in Tricolor Bougainvillea peruviana ‘Thimma’ Based on Transcriptomes
2017-11-09KeyLaboratoryofPlantPhysiologyBiochemistryFujianInstituteofSubtropicalBotanyXiamen36006TheFirstAffiliatedHospitalofXiamenUniversityXiamen36003
(.Key Laboratory of Plant Physiology & Biochemistry,Fujian Institute of Subtropical Botany,Xiamen 36006; .The First Affiliated Hospital of Xiamen University,Xiamen 36003)
ComparisonbetweenBetalainandFlavonolAccumulationinTricolorBougainvilleaperuviana‘Thimma’BasedonTranscriptomes
(1.Key Laboratory of Plant Physiology & Biochemistry,Fujian Institute of Subtropical Botany,Xiamen 361006;2.The First Affiliated Hospital of Xiamen University,Xiamen 361003)
Bougainvilleaperuviana‘Thimma’, accumulating betalains, not anthocyanins occurring in majority of plants. The cultivar was characterized with either white, magenta, and white-magenta(variegated) bracts in the same branches. We first used magenta(Yp), bicolor(Ym) and white(Yw) open inflorescences for transcriptome analysis by sequencing. Candidate genes involving in pigment metablisms were validated by real-time PCR. We obtained 73,325 genes with average length of 616 bp. Out of DEGs, 327 candidate genes of betalain-biosynthesis, 308 flavonoid-biosynthesis genes and 466 anthocyanin- accumulation genes were detected. Eight candidate pigment-related genes were verified, four were responsible for betalain production(PPO, CYP76AD1, cDOPA-5-GT, DODA), and four responsible for flavonoids production(FLS, DFR, LDOX, 3-GT). Expression strength of four betalain production genes was well consistent with betalain-accumulation in Yp, Ym and Yw. Full later three anthocyanidin-producing genes(DFR, LDOX, 3-GT) were first verified and the expression varied in three samples. Expression of all eight candidate genes was well consistent with RNA- sequencing data.B.peruvianacould provide a good tool to study physiological, phytochemical and molecular mechanisms inBougainvillea, effectively eliminating the interference of some factors.
Bougainvilleaperuviana;transcriptome;real-time PCR;betalain;flavonol
Bougainvillea, belonging to Nyctaginaceae family, one member in Caryophyllales, of which nine families are unique for accumulating betalains instead of anthocyanins[1]. So far, no plant has been found that produces both betalain and anthocyanin pigments[1~2]. The mutually exclusive nature of the betalain and anthocyanin production in the plant kingdom is a curious phenomenon[2~3].Bougainvilleasshow the specific characteristic in pigment metabolisms with rich flavonoids and betalains together. It is well known that a long common biosynthesis pathway(from CHS to DFR) was shared between anthocyanin- and flavonol-synthesis. Recently, transcriptome analysis ofB.spectabiliswas performed and genes involved in Betalains-/flavonoid- production were identified[4]. It was the first report of full genes data inBougainvillea. Xu et al.[5]validated expression of betalain-/anthocyanidins-biosynthesis genes detected(CYP76AD1, DODA, cyclo-DOPA-5GT, DFR, LDOX) in fourBougainvilleas. The correlation between betalain-accumulation and expression of betalain-production genes was not well observed. Betalain-accumulation was complex[5~6].
Given the complexity of pigments metabolisms in bougainvilleas, choosing idea materials will help better unclose the function of these genes inBougainvillea.B.peruviana, one of species inBougainvillea, is characterized with tricolor(white, magenta or white-magenta) inflorescence in the same plant contrasting with usual monocolor inflorescence, which is intriguing in horticulture. The variegation is not fixed-pattern, random in white zones or magenta zones as major color. In our study, we usedB.peruvianato validate the function of betalain-/anthocyanin-biosynthesis genes. Three materials ofB.peruvianain the same plant including magenta(Yp), white-magenta(Ym) and white(Yw), are characterized with more similar genetic background. The species is a good system to explore pigment metabolisms.
We aimed at further exploring the correlation between flavonoid-production and betalains-production. The results will be help for better understanding the function of these genes and profile of flavonoid-/anthocyanins-production pathway in betalain-production plants.
1 Materials and Methods
1.1 Plant Materials and Growth Conditions
In the study, the cultivar,B.peruviana‘Thimma’, was planted inBougainvileaGarden in Fujian Institute of Subtropical Botany, China(altitude 24°26′46″, latitude 118°04′04″, annual average temperature 22℃). White-, magenta- and bicolor-bracted samples ofB.peruviana‘Thimma’ were named as Yp, Ym and Yw, respectively(see Fig.1). White-, magenta-, or bicolor inflorescences from stage Ⅱ & Ⅲ[5]were chosen for transcriptome sequencing, measurement of betalain and flavonol, and expression validation of candidate genes.

Fig.1 Materials in the study Y. Whole plant; Yp. Magenta inflorescence; Ym. Variegated inflorescence; Yw. White inflorescence
1.2 cDNA library construction and sequencing
Total RNA of each sample was isolated using a Quick RNA isolation kit(Bioteke Corporation, Beijing, China) and then characterized on a 1% agarose gel and examined with a NanoDrop ND1000 spectrophotometer(NanoDrop Technologies, Wilmington, DE, USA). The RIN(RNA integrity number) values(>8.0) of these samples were assessed using an Agilent 2100 Bioanalyzer(Santa Clara, CA, USA). The construction of the libraries and the RNA-Seq were performed by the Biomarker Biotechnology Corporation(Beijing, China). The purified mRNA was concentrated using Reasy RNA cleaning kit(QIAGEN, Germany) according to the manufacturer’s protocol, and then broken into short fragments. RNA Fragment reagent Kit(Autolab Co., China) and Reasy RNA cleaning kit(QIAGEN, Germany) were used to fragment and retrieve the mRNA that has been condensed for 30 s. Taking these cleaved mRNA fragments as templates, a random primer(N6) and AMV were used to synthesize the first chain, and Secondary Strand Synthesis Enzyme Mix(Autolab Co., China) was used to synthesize the second chain. The resulting cDNAs were then subjected to end-repair and phosphorylation using T4 DNA polymerase and Klenow DNA polymerase. After that, an ‘A’ base was inserted as an overhang at the 3′ ends of the repaired cDNA fragments and adaptors were subsequently ligated to these cDNA fragments to distinguish the different sequencing samples. To select a size range of templates for downstream enrichment, the products of the ligation reaction were purified and selected on a 2% agarose gel. Next, PCR amplification was performed to enrich the purified cDNA template. MinElute DNA Cleaning Kit extracted the synthesized products(QIAGEN, Germany) according to the protocol. Ampure beads(AgencourtAMPure PCR purification Kit, USA) was used for retrieving adapted products. The quality of retrieved cDNA was checked using the Agilent 2100 Bioanalyzer(RNA Nano Chip, Agilent). Finally, the two libraries were sequenced using an Illumina HiSeqTM2500.
1.3Denovoassemblyandunigenefunctionalannotation
Raw reads were extracted from standard flowgram format(SFF) files using base calling and stored as fastq file. Prior to assembly, the raw reads were subjected for the following processing into clean reads, namely, the adaptor sequence was trimmed using In-house developed program, the low quality, contaminated and vector sequence was trimmed using Bcltofastq 1.8.4(http://www.illumina.com.cn/support/sequencing/downloads.aspx), and sequences shorter than 50 bp were also removed from raw reads. The remaining sequences were then assembled using Trinity (http://TrinityRNASeq.sourceforge.net Released on 2013-11-10) and recognized as unigenes(contigs and singletons) with default parameter.
For each library, short reads were first assembled into longer contigs based on their overlap regions. Then, different contigs from another transcript and their distance were further recognized by mapping clean reads back to the corresponding contigs based on their paired-end information, and thus the sequence of the transcripts was produced. Finally, the potential transcript sequences were clustered using the TGI Clustering tool to obtain uni-transcripts[7].
For the functional annotation of unigenes, the remaining sequences that could putatively encode proteins were searched against the following protein databases: NCBI non-redundant protein database(NR), Swiss-Prot protein database, Kyoto 6 Encyclopedia of Genes and Genomes(KEGG) database, Gene ontology(GO) database using BLASTX search, and nucleotide database(NT) by BLASTN with e-value less than 0.00001. Based on the NR annotation, the Blast2GO program was used to obtain GO annotation of unigenes[8]. The GO functional classification was analyzed using WEGO software.
1.4 Quantification of betalains
According to Kugler et al.[9~10], the aqueous pigment extracts of inflorescence from three blooming stages were diluted with McIlvaine buffer(pH 6.0, citrate-phosphate). The betalain content(BC) was calculated by applying the equation proposed by Cai and Corke[11]: BC(mg·L-1)=[(A×DF×MW×1 000/ε×L)], where, A is the absorption value at the absorption maximum, DF the dilution factor, and l the path length(1 cm) of the cuvette. For quantification of betacyanins, the molecular weight(MW) and molar extinction coefficient(ε) of betanin[MW) 550 g·mol-1; ε=60 000 L/(mol cm) in H2O; λ=538 nm;[12]were applied. Quantitative equivalents of the major betaxanthins(bx) were determined by applying the mean molar extinction coefficient {ε=48 000 L/(mol cm) in H2O; λ=480 nm;[13]} and the molecular weight of Dopa-bx(dopaxanthin; MW=390 g·mol-1) for extracts from inflorescences of bougainvilleas[10]. All measurements were performed in triplicates using a UV-vis spectromer(Perkin-Elmer,Überlingen, Germany) equipped with UVWinLab V 2.85.04 software(Perkin-Elmer Instruments, Norwalk, CT).
1.5 Quantification of flavonols
Fresh bracts with early blooming flowers were collected in May 2015 and rinsed in ddH2O. Hydrolysis of the samples followed the method as described by Hertog et al.[14]. Briefly, each sample was repeated three times. First, 0.5 g of ground freeze-dried sample in a flask was mixed with ascorbic acid solution(80 mg in 7.5 mL water). Then the sample was imbibed with the liquid, and 12.5 mL of methanol and 5 mL of 6 mol·L-1HCl were added. Samples were boiled in a water bath under reflux for 2 h at 90℃. Finally, the samples were cooled at room temperature, and then filtered through a 0.22 μm RC filter(SRI, Roth, Germany) prior to injection of 10 μL to HPLC. Triplicates were performed for each sample.
All chemicals were of chromatographic grade in the study. Quercetin and kaempferol(Sigma-Aldrich, Buchs, Germany); isorhamnetin(Sigma-Aldrich, Buchs, Germany). All solution was prepared using ddH2O(18.2 MΩ). HPLC-grade water was obtained from a Milli-Q System(Millipore, Billerica, MA, USA). All standards were dissolved into methanol to a final concentration of 1.0 mg·mL-1and stored at 4℃.
All measurements were performed on a Waters AcquityTMUltra Performance LC Spectrometry(Quattro Premier XE) equipped with an autosampler, and a Masslynx4.1 data handling system. The analytical column was BEH C18(50 mm×2.1 mm I.D., 1.7 μm, Waters, USA). An aliquot of 10 μL solution was injected for HPLC analysis. Chromatograms were acquired at 350 nm for flavonols. Formic acid-acetonitrile(0.1∶99.9, v/v/v) was selected as phase A and TFA-formic acid-acetonitrile-water(0.1∶2∶35∶62.9, v/v/v/v) as phase B[15]. Additional chromatographic conditions: 0 min, B 30%; 12 min, B 48%; 22 min, B 55%; 35 min, B 60%; 40 min, B 30%; the flow rate, 0.3 mL·min-1; the injection volume, 10 μL; temperature, 35℃.
1.6 Quantification by real-time RT-PCR
Eight candidate genes, responsible for betalains, flavonols and anthocyanins were tested by Real-time PCR(Table 1). These candidate genes were polyphenol oxidase(PPO), cytochrome P 450 76AD1(CYP76AD1), cyclo DOPA-glycosyltransferase(cyclo DOPA-5-GT), Dihydroxy-phenylalanine(DOPA)-4,5-dioxygenase(DODA), flavonol synthase(FLS), dihydroflavonol 4-reductase(DFR), Leucoanthocyanidin dioxygenase(LDOX) and anthocyanidin 3-O-glycosyltransferase(3-O-GT) in the study. Actin 1, which showed no significant change in expression throughout the inflorescence development, was used as control gene to normalize the RT-PCR results. The primers were designed using Primer Premier 5.0 with default parameters(see Table 1). Inflorescences were harvested and immediately frozen in liquid nitrogen and stored at -80℃ for RNA extraction.
First strand cDNA was synthesized from 2 μg of total RNA using PrimerScriptTMRT reagent Kit(TaKaRa), according to the manufacturer’s instructions. The PCR reaction was performed in 25 μL, containing 12.5 μL 26SYBR Green Master mix(TaKaRa), 300 nmol·L-1each primer and 2 μL 10-fold diluted cDNA template. The PCR reactions were run in Chromo 4(BioRad, USA) using the following program: 95℃ for 30 s and 45 cycles of 95℃ 5 s, 57℃ 30 s and 72℃ 30 s. The real time RT-PCR reaction was performed in three biological replicates, and three technical repetitions were performed for each biological replicate.
2 Results and Analysis
2.1 Betalain & Flavonol quantification
In this study, we performed betalain quantification of extracts from freeze-dried inflorescences of samples, Yp, Ym and Yw. Among three samples, content of total betalain ranged from 3 085.6(Yp), 841.7(Ym) to 192.2 mg·kg-1fresh weight(Yw)(Table 2). Betaxanthins are major betalains in Yw whereas betacyanins are major betalains in Yp(the ratio of betacyanins to betaxanthins >1). Content of betaxanthins was similar to that of betacyanins in Ym(the ratio of betacyanins to betaxanthins <1)(Table 2).

Table 2 Betalain contents of three samples(Yp,Ym and Yw)
Note:Contents(mg·kg-1fresh weight is means of triplicate determinations(±standard deviation);Means of triplicate determinations with standard deviations at a constant absorbance of 1.00±0.05 atλmax.The same as below.
Table3Flavonols contents of three samples(Yp, Ym and Yw)

MaterialsQuercetinKaempferolIsorhamnetin∑-threeflavonolsRatioofbetalainstoflavonolsYp398.673.9400.72873.223.53∶1Ym726.94121.06335.241183.240.71∶1Yw436.1647.68177.92661.760.29∶1Retentiontime5.43/8.216.41/9.326.60/9.53——
Also, contents of three flavonols(quercetin, kaempferol and Isorhamnetin) were analyzed in the study. The results showed that the highest contents of total flavonols was detected in Ym(1 183.24 mg·kg-1) and the lowest was detected in Yw(661.76 mg·kg-1)(Table 3). In Ym and Yw, the ratio of betalains and flavonols was less than one(0.29∶1 and 0.71∶1) whereas the ratio was more than one(3.53∶1). Of three falvonlos, quercetin is the richest flavonol detected and kaempferol is lest detected among three samples.
2.2Overviewofsequencing,assemblingandfunctionalannotation
To obtain a global view of the inflorescence transcriptome from three samples of Yp, Ym and Yw, we used the open inflorescence for sequencing. For paired-end reads with a single read length of 100 bp and GC percentages of 44.09%(Yp), 43.31(Ym) and 43.81(Yw), respectively, we obtained 19 530 221(Yp), 26 235 946(Ym) and 22 161 608(Yw) mapped reads, respectively. 127 306 genes with average length of 616 bp and N50 length 953 bp(Table 4). 3 199, 3 168 and 6 091 significantly differentially-expressed genes(DEGs) from the groups of Yp vs Yw, Yw vs Ym and Yp vs Ym, respectively. Out of them, 2 425, 2 641 and 4 490 DEGs matched in the public database, respectively(Table 5).
To identify the putative functions of the assembled unigenes in Yp, Ym and Yw, we searched for sequence similarities in five public databases: NR, Swiss-Prot, KEGG, GO database using the BLASTX search and NT by BLASTN. After Blasting, most of the DEGs(75.3% in the group of Yp vs Yw and 83.4% in the group of Yw vs Ym, 73.7% in the group of Yp vs Ym) were annotated by the NR database; the annotated genes are listed in supplementary Table 1 & Fig.2. In three groups, Yp-specific, Yw-specific and Ym-specific genes were involved in secondary metabolisms.
Table4OverviewofthereadsfromtheinflorescencecDNAlibraryofYw,YmandYp

YpYmYwReadsnumber252090853325938327948031Mappedreads195302212623594622161608G+Ccontent(%)44.0943.3143.81Contignumber(intotal)9676661singletonnumber(intotal)257856ORF(intotal)126442Meanlength(bp)616UnigeneN50length(bp)953DEGsnumber5759COGofDEGs870GOofDEGs1773KEGGofDEGs524SWISSofDEGs1890nrofDEGs2419
Table5Differentiallyexpressedgenes(DEGs)amongcomparisongroupsbasedonthethreesamples(Yp,YmandYw)

ComparisongroupYpvsYwYwvsYmYpvsYmDEGs321931656090AnnotatedDEGs242526404489Yp-specificgenes92—735Ym-specificgenes—190184Yw-specificgenes95303—

Fig.2 Venn of DEGs among Yp, Ym and Yw
For the differences in the pigment metabolisms between the three samples, we compared their transcriptome data. By the GO analysis, the distribution of gene functions for the cDNA sequences among three samples were similar in three main categories: biological processes, cellular components, and molecular functions(Fig.3). Among these categories, the cell, cell junction, binding, catalytic activity, organelle, metabolic process and cell process were the groups with the highest representation of genes from three samples. Functional classification and pathway assignments for the three samples were performed through KEGG analyses. Out of 5759 DEGs, 524 DEGs were assigned to KEGG pathways(Table 3). Out of them, 870 DEGs(15%) could be matched in the public database using COG(31%), GO(31%), Kegg(10%), Swiss analysis(33%) and Nr annotation(42%)(Table 4).
Gene Ontology(GO) was used to gain a global overview of putative gene function. Most of the identified transcripts appeared to be genes involved in biological process(57%, 58%, 59%), cellular component(8%, 12%, 11%) and molecular function(35%, 30%, 31%, in Yp vs Yw, Yw vs Ym, Yp vs Ym, respectively)(Supplementary Table 1). Biological processes involved in secondary metabolisms were further analyzed and the results were shown in Table 6. Pigments metabolites play vital roles in bougainvilleas growth, development and resistance to various biotic and abiotic stresses. Based on identified transcripts, 3106 DEGs involved in betalain biosynthesis(327, GO:0005737, GO:0006725, GO:0016705, GO:0019825, GO:0008152), flavonoid biosynthetic process(308, GO:0045431, GO:0009812, GO:0051555, GO:0009963, GO:0009813), anthocyanin accumulation process(466, GO:0009718 GO:0031540, GO:0043481, GO:0046283), glycosylation(742, GO:0006493, GO:0006487, GO:0006486), pigment biosynthetic process(17, GO:0046148) and pigmentation(17, GO:0043473)(supplementary Table 1 and Table 6).

Fig.3 Functional category distribution of transcriptional changed genes using Gene Ontology(GO) (DFR<0.01 & fold change≥2)

PathwayGOIDDEGsBetalainGO:0006725,0016705,0019825,0008152327flavonoidbiosyntheticprocessGO:0045431,0009813,0009812,0051555,0009963308pigmentbiosyntheticprocessGO:004614817glycosylationGO:0006493,0006487,0006486742anthocyaninaccumulationprocessGO:0031540,0043481,0046283,0009718466pigmentationGO:004347316
2.3ExpressionanalysisofcandidategenesinYp,YmandYw
To verify the accurateness of the RNA-Seq data, we validated the candidate genes involving in pigment biosynthesis through real time RT-PCR. Eight candidate genes were verified. Of all, four genes were responsible for betalain production(PPO, CYP76AD1, cDOPA-5-GT, DODA), four genes responsible for flavonoids production(FLS, DFR, LDOX, 3-GT). The leaves was chosen as the control standardizing other three samples.
The expression of four genes(PPO, cDOPA 5GT, DODA, CYP76AD1) was higher in abundant betalain-accumulation samples from Yp, Ym to Yw. Expression strength of four genes in Ym was between that of Yp and Yw. Similar expression strength was discovered in Yw and leaves(Fig.4).
The performance of four genes involving in flavonoids production varied much more. Expression of FLS in Yw was higher than that in Ym, which was higher than that in Yp(Fig.5). Expression of FLS in leaves was significantly less than that in inflorescence. Expression of DFR was higher in Ym than in Yp, which was higher than that in Yw(Fig.5). Similar to FLS, Expression of DFR in leaves was significantly less than that in inflorescence. Expression of LDOX in Yw was higher than that in Yp, which was higher than that in Ym. Expression of LDOX in leaves was similar to that in Ym(Fig.5). Expression of 3-GT was significantly higher in Ym than that in Yp, Yw and leaves and similar among Yp, Yw and leaves(Fig.5).

Fig.4 Expression patterns of candidate genes involving in betalain-production in Yp, Ym and Yw A.PPO; B.CYP76AD1; C.DODA; D.cycloDOPA-5-GT
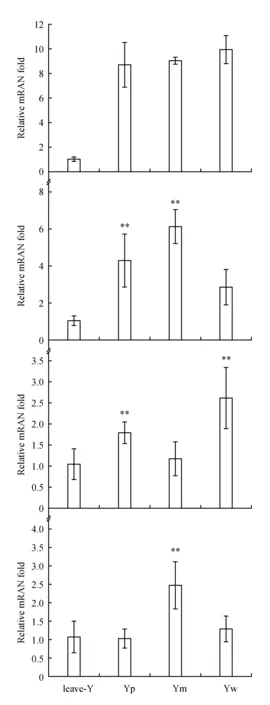
Fig.5 Expression patterns of candidate genes involving in flavonol /anthocyanindin-production in Yp, Ym and Yw A.FLS/F3H; B.DFR; C.LDOX; D.3-O-GT
3 Discussion
3.1 Quantification of betalains and flavonols
Based on betalains quantification, higher betalains accumulation was detected in Yp than that in Ym. Richer betalain accumulation was in Ym than in Yw. The quantification clearly suggests that the different color nuances of inflorescences from bougainvilleas are the result of variations in betaxanthin and betacyanin contents. The results were in accordance with the perceived visual appearance. Betacyanins are major betalains in Yp(ratio of betacyanins to betaxanthins, 2∶1) and betaxanthins are major betalains in Ym and Yw(ratio of betacyanins to betaxanthins, 0.75∶1 and 0.84∶1, respectively)(Table 2). These results were similar to other color cultivars[5].
The three kinds of flavonols are richest in Betalain-production plants[6,16]. Thus, we chose quercetin, kaempferol and isorhamnetin to evaluate contents of flavonols. The quantification results of three flavonols showed that the highest contents of total flavonols was detected in Ym and the lowest was detected in Yw(Table 3). It was observed that the lighter the color was and the smaller the ratio of betalains to flavonols was(Table 3). It seemed that the less betalain accumulated the more flavonols accumulated.
3.2 Expression profile of pigment genes
In the study, all four betalain-producing genes(PPO, CYP76AD1, cyclo-DOPA-5GT and DODA) were verified. The expression level of four genes was well consistent with trends of betalain accumulation(Yp>Ym>Yw). Expression trends of four genes were well consistent with the result of transcriptome sequencing inB.peruviana(supplementary table 1). Our previous study showed no well consistence between betalain accumulation and the expression level of three genes(CYP76AD1, cyclo-DOPA-5GT and DODA) in white, purple, orange and red cultivars[5]. It is possible that not only the expression of these genes, but also the supplementation of DOPA(the substrate of DOD[17]), ascorbic acid(the presence of ascorbic acid reducing agent is necessary to transform o-DOPA-quinone back to L-DOPA[18]) might be a regulatory step for betalain biosynthesis in these plants. Thus, in the study, use ofB.peruviana, Yp, Ym and Yw, eliminate the interference of these factors and verified function of these candidate genes(PPO, CYP76AD1, cyclo-DOPA-5GT and DODA).
Four flavonoids-producing genes(FLS(flavonol-producing), DFR, LDOX, 3-GT(anthocyanin-producing)) were also verified in the study. Flavonol-producing genes(FLS) was first verified inBougainvillea. The expression trends of FLS was well consistent with the result of transcriptome sequencing inB.peruviana(supplementary Table 1). Accumulation of flavonols was Yw>Ym>Yp and betalain accumulation was Yw In our previous study, only two anthocyanin-producing candidate genes(DFR, LDOX) were tested and first validated in floral organs and leaves by Real-time PCR[5]. In the study, we verified full later three anthocyanidin-producing genes(DFR, LDOX, 3-GT) inB.peruviana. Expression of three genes varied in three samples and no obvious correlation was observed. It was speculated that inhibition of anthocyanin production is not caused by anthocyanidin 3-O-GT inBougainvillea. These findings suggest a fully functioning anthocyanin pathway at least to the stage of anthocyanidin-3-O-GT in bougainvilleas. In conclusion, although accumulation of betalains and anthocyanins was excluded, both metabolism pathways were not excluded in the betalain-production plants[2~5,20]. Also even regulating factors could be common, i.e. MYB, in regulating betalain-/anthocyanin-production[21]. Further study maybe report betalain-synthesis pathway exists in anthocyanin-production plants. AcknowledgementsWe appreciated the technical help by Dr. Dahai Wang from Autolab. LTD. Co.,(Beijing,China).SX Xu contributed to the design of the study, data analysis and manuscript writing. QY Huang participated in the cDNA sample preparation, the design of the study and the data analysis. CS Lin helped with RNA Extraction and the data analysis. Yijin Huang contributed to the bioinformatic analysis and helped with the RNA extraction. Ou Hu helped with RNA extraction and discussed the results. 1.Mabry T J.The betacyanins,a new class of red-violet pigments and their phylogenetic significance[M].//Leone C A.Taxonomic biochemistry,physiology,and serology.New York:Ronald Press,1964:239-254. 2.Brockington S F,Walker R H,Glover B J,et al.Complex pigment evolution in theCaryophyllales[J].New Phytologist,2011,190(4):854-864. 3.Harris N N,Javellana J,Davies K M,et al.Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA[J].BMC Plant Biology,2012,12:34. 4.Xu S X,Huang Q Y,Lin C S,et al.Rapid generation and analysis of expressed sequence tags to uncovering inflorescence secondary metabolism ofBougainvilleaspectabilis‘Speciosas’ by pyrosequencing[J].Euphytica,2015,205(3):747-759. 5.Xu S X,Huang Q Y,Lin C S,et al.Transcriptome comparison reveals candidate genes responsible for the betalain-/anthocyanidin-production in bougainvilleas[J].Functional Plant Biology,2016,43(3):278-286. 6.Heuer S,Richter S,Metzger J W,et al.Betacyanins from bracts ofBougainvilleaglabra[J].Phytochemistry,1994,37(3):761-767. 7.Pertea G,Huang X Q,Liang F,et al.TIGR gene indices clustering tools(TGICL):a software system for fast clustering of large EST datasets[J].Bioinformatics,2003,19(5):651-652. 8.Conesa A,Götz S.Blast2GO:a comprehensive suite for functional analysis in plant genomics[J].International Journal of Plant Genomics,2008,2008:619832. 9.Kugler F,Stintzing F C,Carle R.Identification of betalains from petioles of differently colored Swiss chard(BetavulgarisL.ssp.cicla(L.) Alef.Cv.Bright Lights) by high-performance liquid chromatography-electrospray ionization mass spectrometry[J].Journal of Agricultural and Food Chemistry,2004,52(10):2975-2981. 10.Kugler F,Stintzing F C,Carle R.Characterisation of betalain patterns of differently coloured inflorescences fromGomphrenaglobosaL. andBougainvilleasp. by HPLC-DAD-ESI-MSn[J].Analytical and Bioanalytical Chemistry,2007,387(2):637-648. 11.Cai Y,Corke H.Amaranthus betacyanin pigments applied in model food systems[J].Journal of Food Science,1999,64(5):869-873. 12.Wyler H,Meuer U.Zur biogenese der betacyane:versuche mit[2-14C]-dopaxanthin[J].Helvetica Chimica Acta,1979,62(4):1330-1339. 13.Girod P A,Zryd J P.Biogenesis of betalains:purification and partial characterization of DOPA 4,5-dioxygenase fromAmanitamuscaria[J].Phytochemistry,1991,30(1):169-174. 14.Hertog M G L,Hollman P C H,Venema D P.Optimization of a quantitative HPLC determination of potentially anticarcinogenic flavonoids in vegetables and fruits[J].Journal of Agricultural and Food Chemistry,1992,40(9):1591-1598. 15.Yang R Z,Wei X L,Gao F F,et al.Simultaneous analysis of anthocyanins and flavonols in petals of lotus(Nelumbo) cultivars by high-performance liquid chromatography-photodiode array detection/electrospray ionization mass spectrometry[J].Journal of Chromatography A,2009,1216(1):106-112. 16.Xu S X,Lin C S,Huang Q Y,et al.Analysis and comparison of flavonoids in three bougainvilleas by micellar electrokinetic chromatography[J].Bulletin of Botanical Research,2010,30(6):718-724. 17.Takahashi K,Takamura E,Sakuta M.Isolation and expression analysis of two DOPA dioxygenases inPhytolaccaamericana[J].Zeitschrift fur Naturforschung.C,Journal of Biosciences,2009,64(7/8):564-573. 18.Gandía-herrero F,García-carmona F.Biosynthesis of betalains:yellow and violet plant pigments[J].Trends in Plant Science,2013,18(6):334-343. 19.Tsuda S,Fukui Y,Nakamura N,et al.Flower color modification ofPetuniahybridacommercial varieties by metabolic engineering[J].Plant Biotechnology,2004,21(5):377-386. 20.Shimada S,Inoue Y T,Sakuta M.Anthocyanidin synthase in non-anthocyanin-producingCaryophyllalesspecies[J].The Plant Journal,2005,44(6):950-959. 21.Hatlestad G J,Akhavan N A,Sunnadeniya R M,et al.The beet Y locus encodes an anthocyanin MYB-like protein that activates the betalain red pigment pathway[J].Nature Genetics,2015,47(1):92-96. XU Su-Xia1HUANG Qing-Yun1LIN Chun-Song1HUANG Yi-Jin2HU Ou1 三色(Bougainvilleaperuviana‘Thimma’)在转录水平的甜菜色素和类黄酮积累比较 徐夙侠1黄青云1林春松1黄一锦2胡 欧1 (1.福建省亚热带植物研究所植物生理与生化重点实验室,厦门 361006; 2.厦门大学附属医院,厦门 361003) Bougainvilleaperuviana‘Thimma’属于三角梅属,该属植物积累甜菜色素而不是像绝大多数高等植物一样积累花青素。该材料特征同株出现3种颜色:白色、洋红色和白/洋红相间。本研究首次使用3种花色特征的花序(红色Yp、混合色的Ym、白色Yw)作为研究材料进行高通量测序。并通过real-time PCR方法对探测到的花色代谢基因进行验证。共获得平均长度为616 bp的73 325条基因。3种材料的差异显示基因(DEGs)中有327个被注释到甜菜色素合成基因,308个被注释到类黄酮合成基因,466个被注释到花青素合成基因。我们选出8个基因:4个甜菜色素合成基因(PPO,CYP76AD1,cDOPA-5-GT,DODA)和4个花青素合成基因(FLS,DFR,LDOX,3-GT)进行验证。其中,4个甜菜色素合成基因在3种花色材料中的表达较好的正相关于甜菜色素含量。花青素合成途径末端的3个基因(DFR,LDOX,3-GT)在B.peruviana中首次被验证。real-time PCR的验证结果很好的吻合转录组测序的结果。同时,B.peruviana也提供了一个很好的三角梅属植物的生理、生化和分子生物学研究的工具,有效的摒除其他生物学干扰。 Bougainvilleaperuviana;转录组;Real-Time PCR;甜菜色素;类黄酮 S68 A 10.7525/j.issn.1673-5102.2017.02.013 The National Natural Science Foundation of China(Grant No.31071814;31372093);Xiamen Creation Plantform of Science&Technology(3502Z20131004) introduction:XU Su-Xia(1973—),female,Ph.D.,Associate Professor,Major in Plant secondary motablisms. date:2016-07-11
