家兔大型艾美尔球虫四川株的致病性和免疫原性研究
2017-11-09石团员郝力力
王 敏,石团员,李 锐,郝力力*
(1.西南民族大学 生命科学与技术学院 预防兽医系,四川 成都 610041;2.浙江省农业科学院畜牧兽医研究所,浙江 杭州 310021;3.浙江省农业科学院 农产品质量标准研究所,浙江 杭州 310021)
家兔大型艾美尔球虫四川株的致病性和免疫原性研究
王 敏1,石团员2,李 锐3,郝力力1*
(1.西南民族大学 生命科学与技术学院 预防兽医系,四川 成都 610041;2.浙江省农业科学院畜牧兽医研究所,浙江 杭州 310021;3.浙江省农业科学院 农产品质量标准研究所,浙江 杭州 310021)
【目的】为调查兔大型艾美尔球虫四川株的致病性和免疫原性。【方法】本研究利用实验室分离到的大型艾美尔球虫四川株进行了动物感染试验。【结果】5×103和5×104个卵囊组的SPF兔在接种后第4~9天,出现轻微的食欲减少症状;5×105组在接种后第6~8天持续中度腹泻,第8天体重与对照组有明显差异(P≤0.05),但均无死亡现象。感染剂量为5×103时卵囊产量最多,为2.31×108;免疫原性实验中,7×105个卵囊攻毒后,5×103卵囊免疫组卵囊排出最少,为6.28×104,各组体重与对照组相比无明显差异。【结论】大型艾美尔球虫四川株致病性较弱,但免疫原性良好,可作为兔球虫疫苗的备选组分。
兔;大型艾美尔球虫;致病性;免疫原性
【研究意义】我国是家兔生产和贸易大国。2008年,我国家兔全年屠宰量为4.88 亿只,兔肉产量66.0万吨,在世界上占比各为43.8 %、42.1 %,两项指标都是世界第一。除此之外,我国兔毛产量与贸易量均居世界首位[1]。兔球虫病是艾美尔属的球虫寄生在兔肠道或肝脏而导致的一种原虫病,对兔养殖业危害巨大[2]。尽管化学药物是目前控制兔球虫病的常用手段,但随着耐药虫株的不断出现和化学药物残留问题的日益凸现,越来越多的学者寻求用免疫学方法来防控该病[3],使得活疫苗接种在不久的将来成为控制球虫病的主要措施。【前人研究进展】浙江、山东和江苏等省份均是养兔大省,近年来四川省养兔产业也有长足发展,兔球虫病在四川省各地兔场都能检测到,其中尤其以Eimeriamagna较为常见,形态上也最易与其它兔艾美耳球虫相区别。【本研究切入点】本实验在对大型艾美尔球虫成功分离培养的基础上,对其致病性和免疫原性展开了研究。【拟解决的关键问题】以确定能否作为疫苗候选虫株。
1 材料与方法
1.1 供试材料
主要仪器设备和试剂:台式高速离心机(eppendorf 5402型)、Bio-Rad伯乐梯度PCR仪(PTC240型)、2×EasyTaqPCR SuperMix(北京全式金公司)、显微镜(Nikon E200,Japan)、SPF饲养笼(SS3型)、100 和200目不锈钢标准筛、全血DNA小量试剂盒(天根)饱和食盐水、琼脂、麦克马斯特计数板和2.5 %重铬酸钾溶液(自制)。实验动物:母新西兰种兔购买于四川大学实验动物中心,配种新西兰公兔由四川大学实验动物中心提供。E.magna卵囊:含有E.magna卵囊的粪便样本采集于成都旭平兔场。
1.2 试验方法
1.2.1 无球虫SPF兔的培育 配种15只母新西兰种兔,配种后15 d后,检查配种情况。所用不锈钢饲养笼用酒精火焰喷灯喷烧消毒,整个动物房用浓度为28 %的氨水熏蒸48 h。饲料烘烤处理,现用现处理。饮水中按每kg兔体重加入100 μl球虫净,稀释2000倍使用,现配现用。停药1 w后,再用粪便直接涂片法检查球虫卵囊,连查3 d,检查待产母兔有无卵囊排出。仔兔18日龄时断奶,饲喂仔兔颗粒料,并定期补喂婴幼儿奶粉至28日龄。每天取仔兔粪便镜检,连续10 d未检出兔球虫者,即定义为无球虫兔。整个饲养过程中所用的饲料和饮水经无球虫化处理,动物房严格隔离和人员进出,防止外界球虫传入。
1.2.2E.magna单卵囊分离和纯株扩增 在显微镜下对卵囊进行形态学观察,用移液器将含有大型艾美耳球虫的液滴移到另一块载玻片上,继续观察大型艾美耳球虫是否存在,如有其他虫种,用生理盐水稀释,同样用移液器分成数滴,直到一滴生理盐水中只含一个大型艾美耳球虫卵囊时,用移液器移到琼脂块上,在低倍镜下观察,当确定琼脂块内只有一个孢子化的大型艾美耳球虫卵囊时,把此块琼脂装入胶囊。用消毒过的开口器将接种兔的嘴打开,用灭菌镊子夹住胶囊送入口中,灌服少量蒸馏水,确定其咽下,则接种完成。
1.2.3 样品OPG 及每天排卵囊量的计算 每天收取粪便称重,同时称取2 g粪便,采用麦克马斯特法计算每克粪便中的虫卵数(OPG),用OPG乘以粪重即为每天排出的卵囊数。
1.2.4 大型艾美尔球虫18S rRNA 序列的PCR扩增、进化树分析 根据报道的大型艾美尔球虫18S rRNA基因序列,设计上游引物5'-GAAACTGCGAATGG CTCATT-3';下游引物5'-CTTGCGCCTACTAGGCATTC-3',PCR产物克隆后送华大测序。核苷酸序列应用Clustal X 1.81进行比对,分子进化树用MEGA 7构建。
1.2.5 致病性实验 30日龄试验兔共20只,分为A1~A5共5组,每组4只,每笼2只,A2~A5分别按5×102、5×103、5×104和5×105个/只经口感染E.magna孢子化卵囊,A1组设为不感染对照组。分别感染后第2、4、6、8、10、12、14和16天对各组兔称量体重;每天观察并记录各组兔的精神、食欲和排粪便情况,并分别收集感染后第7~15天的兔粪便,用McMaster′s计数法计算OPG,结合排粪量,计算出每组兔的排卵囊总量。综合临床症状、体重和排卵囊量3项指标对E.magna的致病性作出判断。
1.2.6 免疫原性实验 34日龄试验兔共20只,分为B1~B5共5组,每组4只,每笼2只,B1组不免疫不攻毒,B2组不免疫攻毒,B3~B5组免疫剂量分别为5×101, 5×102和 5×103个/只。于免疫后15 d进行攻毒,攻毒剂量为7×105。分别于攻毒后第2、4、6、8、10、12天对各组兔称量体重,计算每2日增重;每天观察并记录各组兔的精神、食欲和排粪便情况,并收集感染后第7~13天的粪便,用McMaster′s计数法计算OPG,结合排粪量,计算出每组兔的排卵囊总量并记录临床症状。
1.2.7 统计学分析 利用统计学软件SPSS 16.0(Tukey test)对各组兔体重进行分析,当组间P≤0.05时,即认为组间存在显著差异,并利用SPSS16.0对分析结果进行作图。
2 结果与分析
2.1 大型艾美尔球虫成都株的生物学特性
单卵囊扩增后的第3代E.magna接种实验兔后,接种后第7天后开始排卵囊,潜伏期是7 d,排卵囊量高峰期是感染后的第9天,持续到16 d排卵结束。如图1所示,E.magna卵囊较大,呈卵圆形,卵壁为黄色或褐色。卵膜孔十分明显,呈堤状分布于卵囊壁之外,有斯氏体和孢子囊余体。卵囊的大小为24.6~39.8 μm×16.4~27.9 μm,平均为33.6 μm×22.5 μm,形态指数为1.49。在恒温28 ℃下,完成孢子化发育的最早时间为28 h,大部分是在48 h。
2.2 18s rRNA PCR检测
从大型艾美尔球虫基因组 DNA中成功扩增出18s rRNA目的条带,片段大小约在1500 bp ,见图2A。应用Mega 6软件对克隆出的18S rRNA的E.magna基因序列与其他11种兔球虫18S rRNA的序列进行同源性分析,并绘制系统发育进化树。由图2B可见,E.magnaSC和E.magnaCzech为同一分支,两者亲缘关系最近。
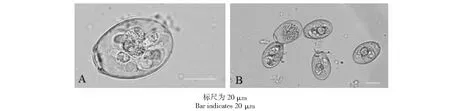
图1 孢子化的第3代大型艾美尔球虫卵囊Fig.1 Sporulated oocyst of third selection of Eimeria magna

M:DL2000 marker;1:阴性对照;2:DNA样本; M: DL2000 marker; 1: Negative control; 2: DNA sample;A:E. magna 18S rRNA基因PCR反应后电泳图;B:E. magna 四川株在基于18S rRNA 基因构建的兔球虫进化树上的分布A:The PCR products of E. magna 18S rRNA; B: Distribution of E. magna SC in the phylogenetic tree of rabbit coccidia based on the 18S rRNA 图2 18S rRNA PCR检测和进化树分析 Fig.2 Detection of 18S rRNA by PCR and phylogenetic tree of the 18S rRNA gene
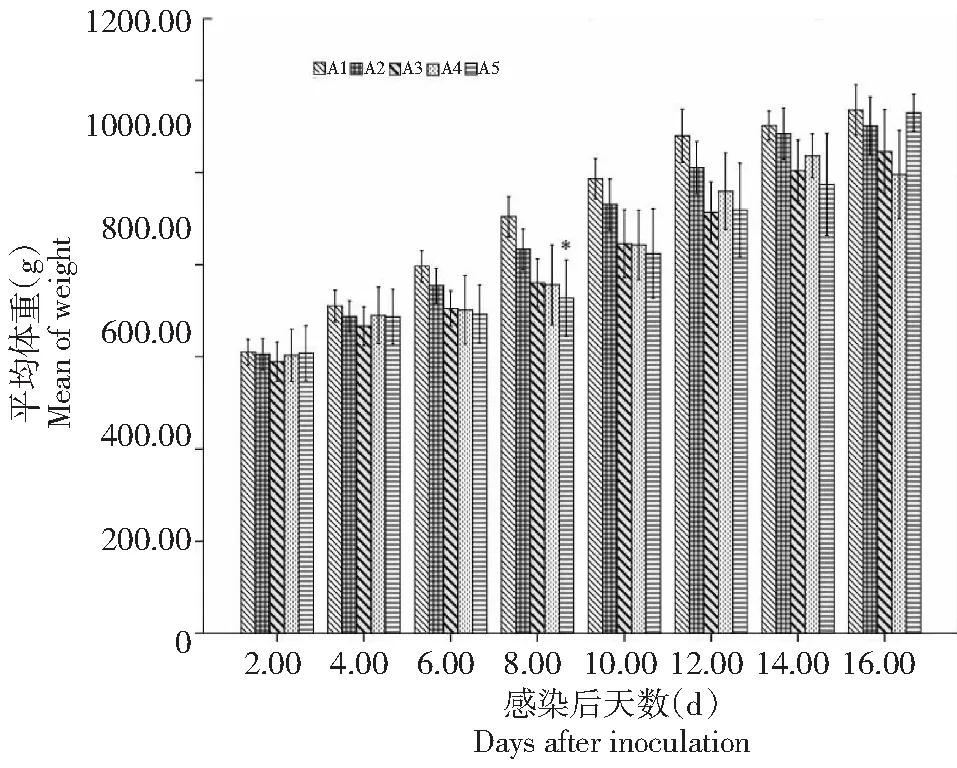
星号表示和对照组相比差异显著(P≤0.05)Asterisks, statistically significant difference between infected and control animals (P≤0.05)图3 不同剂量卵囊感染后每组兔平均体重Fig.3 Mean body weight per rabbit for A1-A5 groups after the infection with different doses of E. magna oocysts
2.3 大型艾美耳球虫的致病性
A3和A4组在感染后4~9 d有轻微的食欲减退现象,A5组在感染后第6天出现便秘、腹泻和食欲减退,其中2只兔子出现中度腹泻,持续3 d(6~8 d),所有组均无死亡现象。所有组体重均持续增长,只有A5组在感染后第8天和对照组相比,体重差异显著(P≤0.05),如图3所示。各组在感染期间总的卵囊产量如表1所示。
2.4 大型艾美耳球虫的免疫原性
B3组在攻毒后4~9 d有轻微的食欲减退现象,B2组中度腹泻持续了4 d(攻毒后6~9 d)。如图4所示,和B1组相比,其余组每隔2 d的体增重无显著差异。攻毒后7~13 d内各组卵囊产量如表2所示,B3和B4组产生了部分保护力, B5组则保护力最好,粪便中未检出卵囊。
表1感染不同剂量后7~16d每组排出的卵囊总量
Table 1 Total oocyst output 7-16 days after infection with different doses ofE.magnaoocysts

组别Group 日龄(d)Age感染剂量(个)Inoculationdoses排出卵囊(个)OocystoutputA1300ndA2305×1021.22×108A3305×1032.31×108A4305×1047.19×107A5305×1057.48×107
注:nd:未检出。
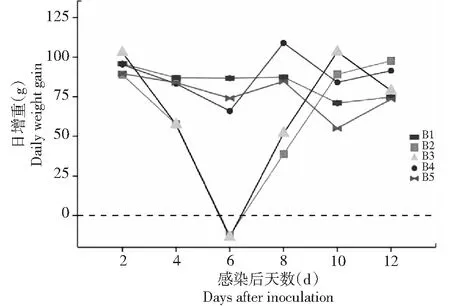
图4 7×105个卵囊攻毒后每组组日增重Fig.4 Daily weight gain after challenge inoculation with 7×105 oocysts of E. magna
3 讨 论
本实验中成功分离到大型艾美尔球虫四川株,PCR得到的18s rRNA序列为1528 bp,经NCBI blast比对后,与捷克株和法国株同源性最高,均为99 %,但捷克株与其序列覆盖度最高,为100 %;其次为法国株,覆盖度为94 %,进化树上和捷克株为同一分支,说明两者亲缘关系最近。构建进化树的意义在于建立分类系统和确定亲缘关系[4],但只根据其18S r RNA也很难鉴别兔球虫种类。如梨形艾美耳球虫和黄艾美耳球虫 2 个种,18S rRNA同源性达99.6%,具有几乎一致的序列,但不能由此确定它们是一个种, 应该和传统分类方法(卵囊形态、生活史、致病性等生物学特征)相结合方能确定。
兔球虫中不同种具有不同的致病力,如E.perforans只有2代裂殖生殖,致病力较弱[5],而E.magna具有4代裂殖生殖,致病力强或者中等,不同的E.magna地理株具有不同的毒力。如Licois D等[6]分别用2.5×103,1×104和 4×104剂量感染42日龄新西兰大白兔, 1×104和4×1042 个剂量组在感染后7 d 出现增重迟缓现象,和空白对照组相比,差异极显著(P≤0.01),但均无死亡现象。林昆华等[7]用分离到的E.magna虫株感染35日龄新西兰大白兔,1×105和1×106剂量组出现厌食、体重下降、腹泻,甚至有血便等临床症状,感染后4~8 d体重下降很明显, 并有死亡现象,死亡率分别为12.5 % and 25 %。本实验中,只有5×105剂量组感染后第8天和空白对照组相比,差异显著(P≤0.05),其余均差异不明显。E.magna四川株感染剂量为5×103时排出的卵囊最多,为2.31×108;Licois D等[6]感染剂量为81个卵囊时,排出卵囊即可达1.7×108。相比之下,四川株致病力弱,产卵囊量少,在感染后第7天可在粪便中检测到卵囊,第9天为排卵高峰,与Licois D报道一致。
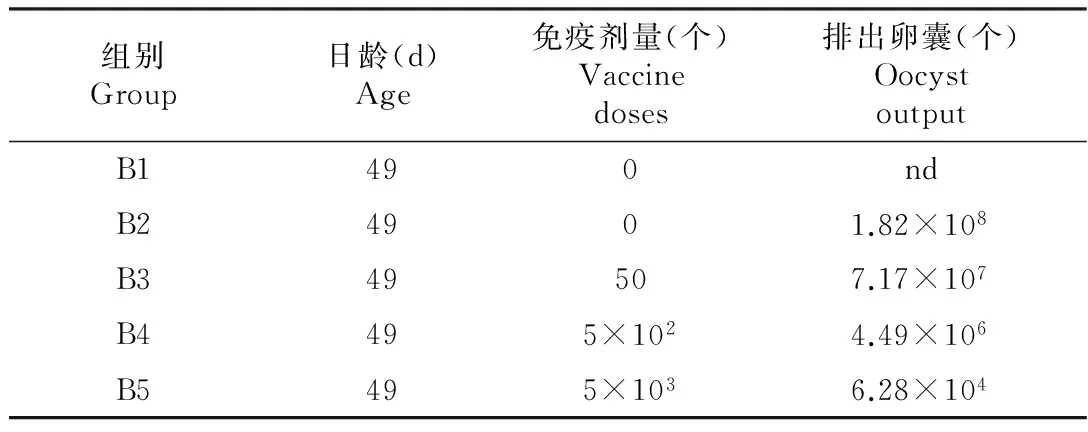
表2 攻毒后不同免疫剂量组7~16 d每组排出的卵囊总量
注:nd:未检出。
本实验中,7×105个卵囊攻毒后,免疫剂量为5×103时可产生较好的保护力,粪便中只检出6.28×104卵囊;Drouet-Viard[8]等用3.5×103早熟株对25~29日龄的新西兰大白兔进行免疫,均可产生较好的保护力,粪便中未能检出卵囊;Akpob Y等[9]采用2.5×103早熟株卵囊免疫后攻毒(攻毒剂量为5000个卵囊),和空白对照组相比,日增重无显著差异,但仍然有较多卵囊排出(48.3×105)。由此可见,E.magna四川株具有良好的免疫原性,第三代卵囊即可对被免疫动物产生较好的保护力。
综上所述,本实验分离到的大型艾美尔球虫四川株致病力弱、免疫原性良好,无需进行早熟株的进一步选育,可作为兔球虫疫苗的候选组分之一。
[1]沈景林. 我国兔产业现状、发展前景及效益分析[J]. 吉林畜牧兽医,2008,29(3):43-45.
[2]闫文朝,王天奇,索 勋,等. 家兔球虫病的研究进展[J]. 中国草食动物科学,2010,40(11):1200-1205.
[3]BHAT T K, JITHENDRAN K P, Kurade N P. Rabbit coccidiosis and its control:a review[J]. World Rabbit Science, 1996, 4(1): 37-41.
[4]KVICEROVA J, PAKANDL M, HYPSA V, et al. Phylogenetic relationships amongEimeriaspp.(Apicomplexa,Eimeriidae)infecting rabbits:evolutionary significance of biological and morphological feature[J]. Parasitology, 2008, 135(4): 443-452.
[5]石团员,魏 巍,付 媛,等. 穿孔艾美耳球虫、点滴复膜酵母与兔腹泻的关系[J]. 浙江农业学报,2015,27(2):160-164.
[6]LICOIS D, COUDERT P, BOIVIN M.Eimeriamagna:pathogenicity, immunogenicity and selection of a precocious line[J]. Veterinary Parasitology, 1995, 60(1-2): 27-35.
[7]史晓春,林昆华. 兔大型艾美耳球虫的致病性、临床及病理组织学研究[J]. 北京农业大学学报,1995, 21(3):341-348.
[8]DROUET V F, LICOIS D, COUDERT P, et al. Acquired protection of the rabbit(Oryctolaguscuniculus) against coccidiosis using a precocious line ofEimeriamagna:effect of vaccine dose and age at vaccination[J]. Veterinary Parasitology, 1997, 69(3-4): 197-201.
[9]AKPO Y, KPODEKON M T, DJAGO Y, et al. Vaccination of rabbits against coccidiosis using precocious lines ofEimeriamagnaandEimeriamediain Benin[J]. Veterinary Parasitology, 2012, 184(1): 73.
Eimeriamagna:PathogenicityandImmunogenicityofStrainIsolatedfromRabbit(Oryctolaguscuniculus)inSichuanProvince
WANG Min1, SHI Tuan-yuan2, LI Rui3, HAO Li-li1*
(1.College of Life Science & Technology, Southwest University for Nationalities, Sichuan Chengdu 610041, China; 2.Institute of Animal Husbandry and Veterinary Medicine, Zhejiang Academy of Agricultural Science, Zhejiang Hangzhou 310021, China; 3.Institute of Quality and Standard for Agro-products, Zhejiang Academy of Agricultural Sciences, Zhejiang Hangzhou 310021, China)
【Objective】The present paper aims to explore the pathogenicity and immunogenicity ofEimeriamagnaisolate in Sichuan province. 【Method】Eimeriamagnafrom our laboratory were used to infect experimental rabbits. 【Result】In the pathogenicity assay, the slight decreases of appetite in SPF rabbits infected with 5×103and 5×104sporulated oocysts during 4-9 days after the infection were observed, respectively; Mild diarrhea of rabbits infected with 5×105oocysts during 6-8 days after the infection lasted for 3 days, and the significant difference of mean body weight per rabbit were observed compared with control group (P≤0.05), however, no death in all groups was found. 5×103oocysts were necessary to attain the maximum oocyst output (2.31×108). In the immunogenicity assay, only 6.28×104oocysts in rabbits immunised with 5×103sporulated oocysts after challenge with 7×105oocysts were detected. Compared with control group, no significant difference in body weight gain in other groups was found. 【Conclusion】TheE.magnaisolate had low virulence but immunogenicity in rabbit, indicating its feasibility as a vaccine component.
Rabbit;Eimeriamagna; Pathogenicity; Immunogenicity
1001-4829(2017)5-1229-04
10.16213/j.cnki.scjas.2017.5.042
2016-05-20
西南民族大学中央高校基本科研业务费青年教师基金项目(2016NZYQN34);四川省教改项目“兽医专业学位研究生教育实践基地建设”
王 敏(1991-),女, 四川泸州人,硕士,研究方向为分子寄生虫学,E-mail: 18683563476@126.com,手机:18683563476,*为通讯作者:郝力力, E-mail: leelee_hao@ 126.com。
S855.9
A
(责任编辑 陈 虹)
