2015年夏季长江口及其邻近海域渔业生物群落结构分析
2017-11-07汤昌盛戴芳群王彦涛
汤昌盛,张 芳,冯 颂,戴芳群,王彦涛
2015年夏季长江口及其邻近海域渔业生物群落结构分析
汤昌盛1,2,3,张 芳1,3,冯 颂1,3,戴芳群4,王彦涛1,3
(1.中国科学院海洋研究所,海洋生态与环境科学重点实验室,青岛 260071;2.中国科学院大学,北京100049;3.青岛海洋科学与技术国家实验室,海洋生态与环境科学功能实验室,青岛 266071;4.中国水产科学研究院黄海水产科学研究所,渔业资源与生态系统研究室,青岛 266071)
根据2015年6月(夏季)长江口及其邻近海域的底拖网调查数据,分析了该海域渔业生物种类组成、优势种、群落结构及其多样性特征。结果表明,调查共捕获渔业生物93种,其中底层鱼类56种、中上层鱼类7种、头足类10种和甲壳类20种,以底层鱼类和甲壳类为主。优势种包括小黄鱼(Larimichthy-spolyactis)、鳀(Engraulis japonicus)和带鱼(Trichiurus haumela)。鱼山渔场及舟山渔场近岸区和长江口渔场离岸区群落多样性较高,舟山渔场离岸区和大沙渔场南部多样性较低。渔业生物群落可分为3组:20 m等深线附近的近岸区、20~50 m水深的长江口渔场外部和大沙渔场南部、以及50~75 m水深的舟山渔场和鱼山渔场中外部。渔业生物群落结构不同组间差异主要由小黄鱼、鳀、带鱼和刺鲳(Psenopsis anomala)引起。
长江口海域;夏季;渔业生物;群落结构
长江口是我国最大的河口,受长江径流、台湾暖流、浙江沿岸流、苏北沿岸流和黄海冷水团等水系相互消长的影响,生态环境复杂多变,营养物质丰富,饵料生物丰盛,是众多渔业生物产卵、育幼和索饵的重要场所,形成舟山渔场、长江口渔场和吕四渔场等优良渔场,是我国重要的渔业基地[1],在我国近海生态系统研究中也占有重要地位[2]。因此,我国学者对长江口渔业生物开展了广泛研究,其焦点主要在于鱼卵、仔稚鱼分布及其数量变动[3-7],渔业生物群落结构[2,8-12]和高营养层次功能群[13]等方面。每年6~9月为该海域的伏季休渔期[14],夏季尤其是6月的调查资料更能代表休渔期早期的资源量水平,对于评估伏季休渔制度施行的效果具有重要的基础意义[15-16],但近十余年来对该海域夏季渔业生物群落结构的研究较少。本文基于2015年6月(夏季)对长江口及其邻近海域渔业生物的底拖网调查,对该海域渔业种类组成、优势种、群落多样性特征、群落结构等进行了分析,旨在为该海域渔业生物群落结构和生物多样性研究提供数据基础。
1 材料与方法
1.1 材料来源与处理
材料取自2015年6月(夏季)对长江口及其邻近海域进行的渔业生物底拖网调查,调查区域为 122°01′E~124°58′E、28°33′N~33°00′N,以与等深线近乎垂直的方式布设了9条大致等间隔的断面,共计34个站位(图1)。调查船为黄海水产研究所“北斗”号,其网具参数为:网目836目×20 cm,网口周长167.2 m,网具总长度83.2 m,囊网网目10/2.4 cm,拖速为3 kn。每站拖网0.5~1 h,在实验室内对渔获生物进行种类鉴定和生物学测定,记录每一种的重量和尾数,并换算为单位时间的生物量(kg·h-1)和单位时间尾数(ind·h-1)。

图1 长江口及其邻近海域调查站位图Fig.1 Sampling station map in the Yangtze River Estuary and adjacent sea areas in this study
1.2 相对重要性指数
采用 PIANKA[17]的相对重要性指数(Index of relative importance,IRI)作为优势度的指标:

式中,W%和N%分别是某种渔业生物占所捕总量的个体重量百分比和个体数量百分比,F为某一种出现的站数占所有采样站站数的百分比。当IRI≥1 000时,该种为优势种;当IRI为100~1 000时,该种为常见种,优势种和常见种一起合称为重要种成分[12]。
1.3 群落多样性指数
在数据分析前,对数据进行标准化处理,即选择在各站位出现频率高于5%,且至少在某一站位中尾数含量1%以上种类用于数据分析[18],从而避免稀有种对数据分析结果的影响。WILHM[19]认为不同种类及同种类个体大小差异很大,用生物量表示的多样性更接近种类间能量分布,因此本文根据生物量计算渔业生物群落种类多样性[11,20-21]。
采用了以下指标对渔业生物群落多样性进行分析:
Shannon-Weaver多样性指标[22]

Margalef种类丰富度指数[23]
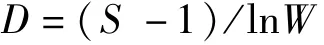
Pielou均匀度指数[24]
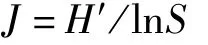
式中,S为渔业生物种类数,W为渔获总重量,Pi为第i种渔获物重量占总渔获物重量的比例。
根据标准化后得到的渔获生物重量数据,利用Shannon-Weaver多样性指数(H′)、Margalef种类丰富度指数(D)和Pielou均匀度指数(J)对长江口及其邻近海域渔业生物多样性进行分析,其综合了渔业生物的丰富度和均匀度,可度量渔业生物群落的生态多样性和异质性[25]。
1.4 群落结构分析
根据相对生物量(kg·h-1)平方根转换后计算Bray-Curtis相似性系数矩阵,应用未加权的组平均方法(UPGMA)进行聚类(CLUSTER)对渔业生物群落进行结构分析。使用单因子相似性分析(ANOSIM)对不同组群落结构差异进行显著性检验,并利用相似性百分比分析(SIMPER)分析造成各组内群落结构相似的典型种以及造成不同组之间群落结构差异的分歧种[26]。
1.5 数理统计
以上数据统计分析和作图采用软件Microsoft Excel 2010、Surfer 12和PRIMER 5完成。
2 结果与分析
2.1 渔业生物组成
2015年夏季长江口及其邻近海域调查共捕获渔业生物93种,其中底层鱼类56种、中上层鱼类7种、头足类10种和甲壳类20种,分别占总种数的 60.2%、7.5%、10.8%和 21.5%。
底层鱼类主要包括小黄鱼(Larimichthys polyactis)、带 鱼 (Trichiurus haumela)、刺 鲳(Psenopsis anomala)、龙 头 鱼 (Harpadon nehereus)、细纹狮子鱼(Liparis tanakae)和绿鳍鱼(Chelidonichthys kumu)等。
中上层鱼类主要包括鳀(Engraulis japonicus)、银 鲳 (Pampus argenteus)、竹 筴 鱼(Trachurus japonicus)和 圆 腹 鲱 (Dussumieria elopsoides)等。
头足类主要包括太平洋褶柔鱼(Todarodes pacificus)、日本枪乌贼(Loligo japonica)、长蛸(Octopus variabilis)和剑尖枪乌贼(Uroteuthis edulis)等。
甲壳类则主要包括细点圆趾蟹(Ovalipes punctatus)、三疣梭子蟹(Portunus trituberculatus)、双斑蟳(Charybdis bimaculata)、葛氏长臂虾(Palaemon gravieri)、须 赤 虾 (Metapenaeopsis barbata)和口虾蛄(Oratosquilla oratoria)等。
鱼类平均渔获重量和平均尾数都相对最高,达 99.410 kg·h-1和 14 672 ind·h-1,分别占总平均重量和总平均尾数的93.5%和91.1%(表1)。
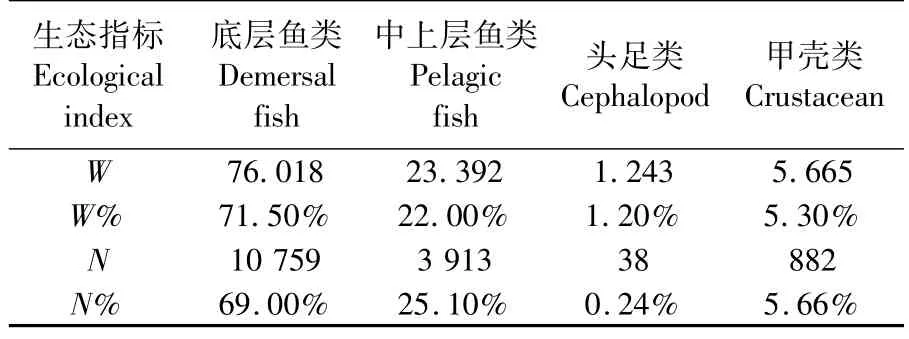
表1 长江口及其邻近海域渔业生物组成Tab.1 Composition of fishery species in the Yangtze River Estuary and adjacent sea areas
2.2 优势种组成
利用相对重要性指数(IRI),对渔获物群落优势种成分进行分析。重要种成分重量占总渔获重量的 88.00%,其尾数占总渔获尾数的91.15%。本次调查的优势种(IRI≥1 000)包括3种,分别为小黄鱼、鳀和带鱼,占总渔获重量的83.84%,占总渔获尾数的88.89%。常见种包括细点圆趾蟹和刺鲳(表2)。
2.3 群落多样性特征及其分布
长江口及其邻近海域渔业生物H′、D和J平均值分别为 1.100、3.100和 0.449。Shannon-Weaver多样性指数H′高值区主要位于鱼山渔场及舟山渔场的近岸区和长江口渔场的东部离岸区,低值区主要位于舟山渔场的西南部及大沙渔场南部(图 2)。H′最高值出现在 S32站,为2.369;其次为 S20站,为 2.334;最低为 S10站,仅为0.045。丰富度指数D高值区分布与H′相同,位于鱼山渔场近岸区和长江口渔场离岸区,低值区则位于鱼山渔场中部、舟山渔场西部近岸区和大沙渔场南部。最高值出现在S32站,达6.823;其次为 S12站,为 5.887;最低值出现在S10站,为0.827。均匀度指数 J分布与H′分布几乎相同,J最高值出现在S31站,为0.825;其次为 S32站,为0.820;最低值为 S10站,仅为0.025。
2.4 群落结构分析
通过CLUSTER聚类分析,将2015年夏季长江口及其邻近海域渔业生物群落分为3组(图3)。其中,S27站捕捞个体过少,不予分组[20];S16站由于76.6%为单一种细点圆趾蟹并且其占该种总捕捞量的90.5%,不宜加入群落聚类分析,亦不予分组。将S2、S8等16个站归为组1,S12、S4等8个站归为组2,其余 S33、S34等8个站归为组3。ANOSIM分析结果显示,不同站位组间群落结构种类组成差异极显著(R=0.659,P<0.001),站位组两两之间差异也极显著(P<0.001)。组1主要分布于20 m等深线附近的近岸区域,组2主要位于20~50 m等深线之间31°N以北的长江口渔场外部和大沙渔场南部,组3则分布于50~75 m等深线之间舟山渔场及鱼山渔场的中外部(图4)。
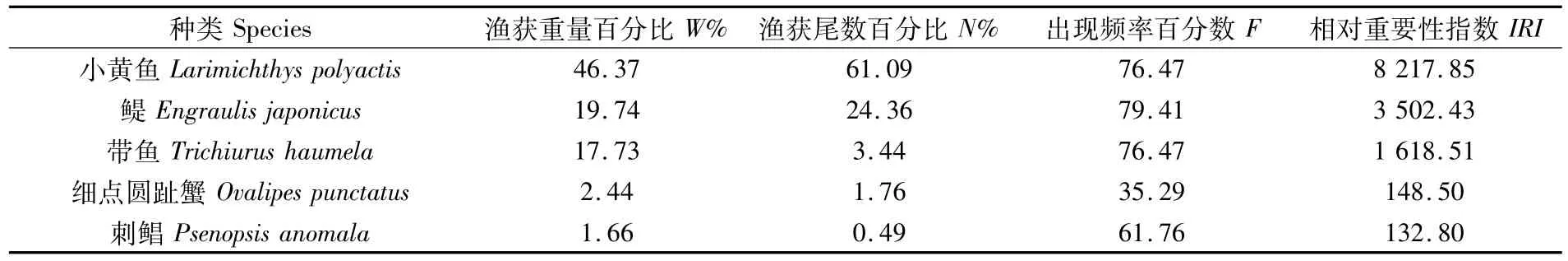
表2 长江口及其邻近海域重要种组成Tab.2 Dominant species in the Yangtze River Estuary and adjacent sea areas

图2 渔业生物多样性指数H′、D和J的空间分布Fig.2 Spatial distribution of H′,D and J of fishery resources
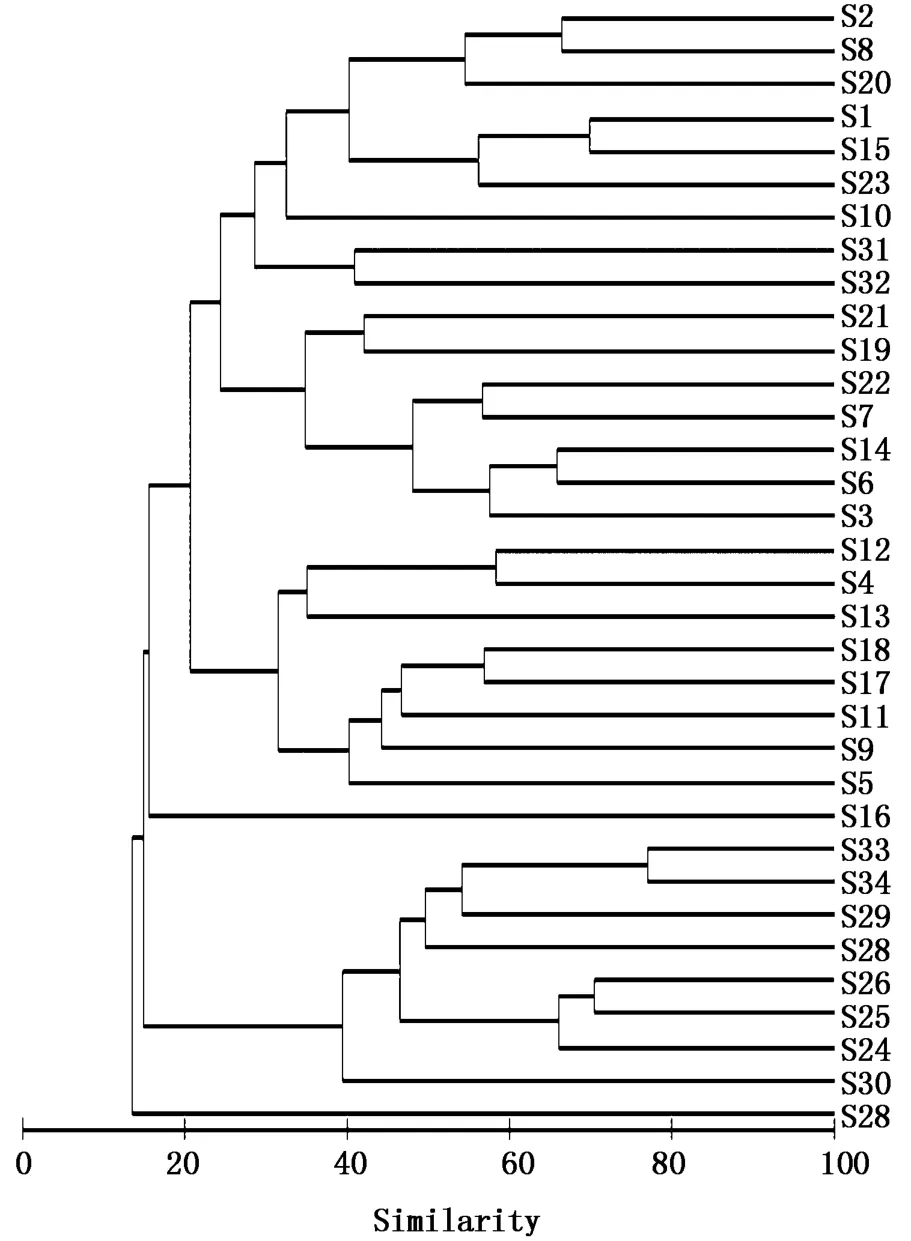
图3 长江口及其邻近海域夏季渔业生物群落聚类分析Fig.3 Cluster analysis for the fishery species communities based on Bray-Curtis similarity matrices in the Yangtze River Estuary and adjacent sea areas in summer
SIMPER分析结果(表 3)显示,组 1、2、3各站位组的组内平均相似性分别为18.89%,22.14%和41.83%。组1的典型种主要包括小黄鱼和鳀,这2种对组内平均相似性贡献达83.76%;组2的典型种包括三疣梭子蟹、小黄鱼、黄鮟鱇(Lophius lityylon)、太平洋褶柔鱼、细点圆趾蟹和鳀,以上6种对组内平均相似性贡献达92.88%;组3的典型种主要包括带鱼和刺鲳,这两种对组内平均相似性贡献达97.32%。
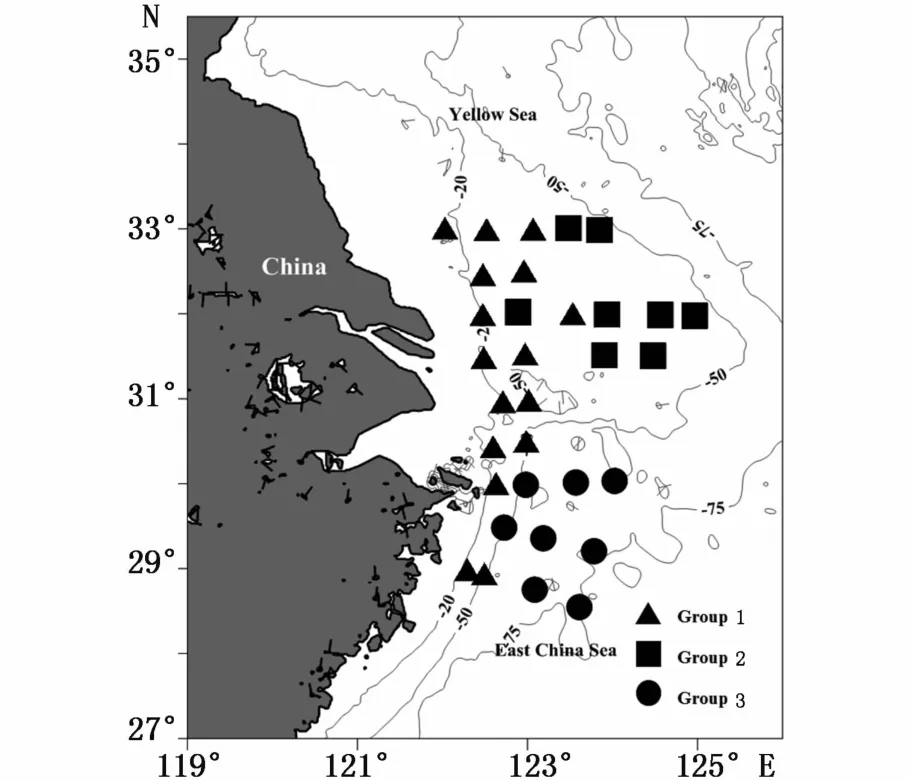
图4 长江口及其邻近海域夏季渔业生物群落的空间分布Fig.4 Spatial distribution of fishery species communities in the Yangtze River Estuary and adjacent sea areas in summer
组1和组2,组1和组3,组2和组3之间的平均非相似性分别为 93.25%,96.05%和97.24%。不同组间形成差异的主要分歧种包括小黄鱼、鳀、带鱼和刺鲳。对组1和组2间平均非相似性贡献较高的分歧种主要为小黄鱼和鳀,组1和组3的分歧种主要包括带鱼、小黄鱼和鳀,组2和组3的分歧种主要为带鱼以及刺鲳。
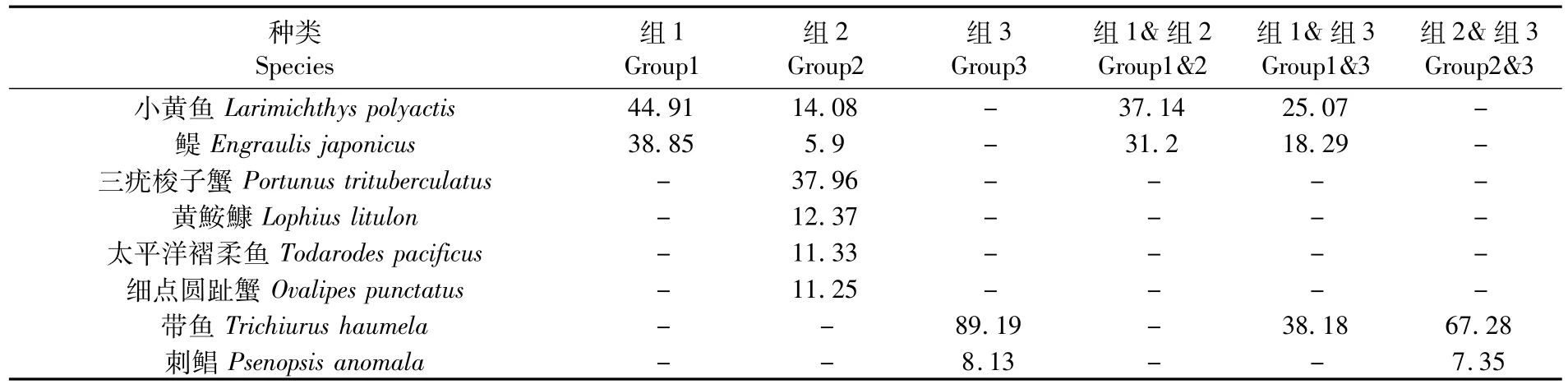
表3 夏季组内典型种对组内平均相似性和组间分歧种对组间平均非相似性的贡献百分比(≥5%)Tab.3 Typical species and their contributions to the average within-group similarity,discriminating species and their contributions to the average between-group dissimilarity in the Yangtze River Estuary and adjacent sea areas in summer(≥5%)
3 讨论
3.1 渔业生物种类和优势种组成
从种类数来看,本次调查渔业生物以底层鱼类和甲壳类为主,这和21世纪以来资料记载的长江口及其毗邻海域各季节以底层鱼类和甲壳类为优势类群[2,8]的结论相符,在黄海南部也有同样的发现[27]。然而从20世纪90年代初期至末期,甲壳类在渔获量中所占的比例均较低且明显下降[10],说明21世纪以来渔业生物种类组成发生了变化,甲壳类比例增加。这可能主要是由于近十余年以来,高强度的捕捞致使传统经济鱼类资源衰退,甲壳类被捕食的压力和饵料竞争的压力下降而得以繁衍和生长,导致渔业生物中甲壳类种类数及资源量相对增加[20,28-29]。
本次调查渔业生物种类数略高于历史同期调查,但平均渔获量较同期要高得多(表4);3个优势种在以往同期调查的优势种中均有出现,但每年优势种的组成有所差异,表明近年来长江口及其邻近海域优势种发生了变化。
本次出现的主要渔业生物小黄鱼、带鱼、刺鲳、鳀、银鲳、细点圆趾蟹、三疣梭子蟹、双斑蟳、长蛸、日本枪乌贼等均是以往调查[2,8,12]出现的主要常规种,渔业生物种类数虽然较高但在正常范围内。本航次调查时间为该海域休渔期开始的时期,渔业生物得以自然生长和繁殖,可能是平均渔获量较高的主要原因之一。其中贡献最大的为小黄鱼,占总平均渔获量的46.37%;其平均渔获量达 49.30 kg·h-1,比以往高得多,如2006年 6月为5.25 kg·h-1[2],2008年6月仅为4.55 kg·h-1[12]。
从优势种的变化可以看出,带鱼一直处于很高的优势地位,表明带鱼是该群落最主要的优势种,生物量和数量都较丰富且分布区域较大,这和资料记载的6~10月长江口及其邻近海域是带鱼繁衍、索饵的重要场所和传统的作业渔场[14,33]结论相符。竹筴鱼、鳄齿鱼(Champsodon capensis)、发光鲷(Acropoma japonicum)等优势地位明显下降,不再成为优势种;刺鲳的优势地位也有所下降,在本次调查中归为常见种(未列出)。小黄鱼和鳀优势度则增加并成为6月份的优势种。这和资料记载的鳀5月下旬到6月中旬随沿岸水温升高至长江口毗邻海域产卵[33],小黄鱼的重要产卵场和幼鱼索饵场位于长江口邻近海域[14,33]等结论相符合。
3.2 群落多样性特征及其分布
通过与历史同期调查资料的比较(表5),可以看出,本次调查渔业生物Shannon-Weaver多样性指数H′与历史同期调查较为一致,但其最小值低于历史平均水平,也低于马克平等[34]提出的H′范围1.5~3.5;Pielou均匀度指数 J也略低于历史同期调查,其主要原因可能是生物量较高。有研究表明,鱼类生物群落的多样性与生物量呈显著负相关关系[12,30],而本次调查生物量显著高于历史同期水平,故而H′与J相对较低。但本次Margalef种类丰富度指数D则例外,平均值和最大值都比同期调查平均水平高出1倍以上。主要原因可能有两点:一是前文提及的种类数较多,而丰富度指数D反映的是生物群落中物种数目的多寡,故而D也较大;二是群落中渔业生物多以幼体为主,如平均体质量小黄鱼仅为5.18 g、银鲳14.9 g,比2001年6月的小黄鱼22.6 g、银鲳71.5 g[30]低,在相同种类数的站位,其个体越小,那么相对生物量一般会越低,丰富度指数D则越大。

表4 长江口及其邻近海域渔业生物种类和优势种组成变化Tab.4 Species component and dominant species component in the Yangtze River Estuary and adjacent sea areas in different years
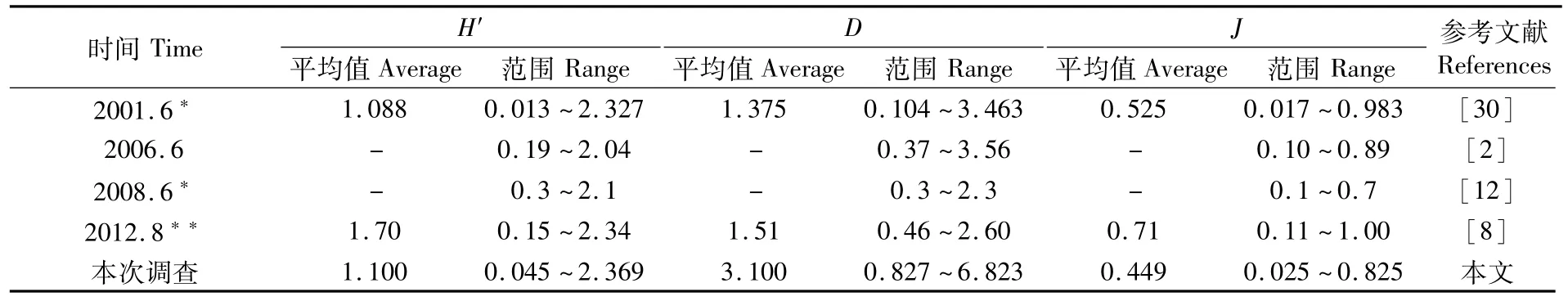
表5 长江口及其邻近海域渔业生物多样性指数变化Tab.5 Variations of diversity indexes of fishery species in the Yangtze River Estuary and adjacent sea areas
2015年夏季多样性指数呈斑块状分布。多样性较高的区域主要有两个,123°E以西浙江近岸的舟山渔场及鱼山渔场近岸区,以及31°N~32°N之间长江口渔场的离岸区;而多样性较低的区域也主要有两个,舟山渔场123°E以东的西南海域,以及32°N以北的大沙渔场的南部。在多样性指数低值区域,相对生物量都很高,如舟山渔场西南部S25站、S24站分别为180.54 kg·h-1、237.75 kg·h-1,大沙渔场南部 S10站、S7站分别达 422.59 kg·h-1、1 050.66 kg·h-1;但这些生物量高的站位单种比例都过高,优势种地位过于突出,如S25站和S24站带鱼分别占88.6%和84.12%,S10站和S7站小黄鱼分别占99.4%和97.7%,因而多样性指数较低,进一步解释和印证了多样性指数和相对生物量的负相关关系。
3.3 群落结构
聚类分析结果表明,夏季长江口及其邻近海域渔业生物群落划分为3组,聚类分组可能最主要由水深决定(图4):组1为20 m等深线附近的近岸区域,组2为20~50 m水深的长江口渔场外部和大沙渔场南部,组3为50~75 m水深的舟山渔场和鱼山渔场中外部。
小黄鱼和鳀为组1近岸区群聚的主要典型种,说明夏季小黄鱼和鳀主要分布于近岸区。组2的主要特点是三疣梭子蟹和细点圆趾蟹等甲壳类比重较大,典型种较多,这与本次调查组2所在海区为多样性指数高值区之一的结论一致。带鱼和刺鲳为组3群聚的主要典型种,表明带鱼和刺鲳主要分布区位于舟山渔场和鱼山渔场中外部,这与资料记载的带鱼6月广泛分布于鱼山渔场和舟山渔场[35],刺鲳以舟山渔场分布密度最高,其次为鱼山、舟外和长江口渔场[36]等结论相符。
小黄鱼、鳀、带鱼和刺鲳是不同组间群落结构形成差异的主要分歧种,不同组间形成差异的主要原因可能有两个:
其一为小黄鱼和鳀季节性洄游。有研究指出,小黄鱼5~6月集中在舟山渔场、长江口渔场、吕四渔场禁渔线外侧,7~9月进入大沙渔场索饵[37];鳀夏季初期主要活动于长江口邻近海域,秋季离开近岸水域游向黄海中南部和东海北部[31,33]。一般而言,温度是影响鱼类洄游分布最重要的因素之一[38],温度控制下的季节性洄游决定了其空间分布,从而成为近岸区与其它两个海区的主要分歧种。
其二为最主要的水深。组1的主要典型种鳀为中上层鱼类,而组2的典型种除小黄鱼和鳀外,主要为三疣梭子蟹、细点圆趾蟹、黄鮟鱇和太平洋褶柔鱼等底层深水种,组3典型种带鱼和刺鲳[36]也都是底层深水种,渔业生物的水深栖息特性决定了其群落结构的差异。在斜坡和大陆架海区鱼类群落的研究中,也有学者认为水深是影响鱼类群落结构变化最主要的环境因子,鱼类的区系组成随着水深梯度变化[39],东海大陆架鱼类群落种类组成沿水深梯度的变化比较明显[40]。
有研究认为,渔业生物群落的空间分布规律与离岸距离、河流入海和地理位置等一系列自然生态环境的变化密切相关,它们通过影响温度、盐度等海洋水文条件来影响渔业生物群落结构[20];也有研究指出,三峡工程[41]、长江口深水航道工程[9]等大型水利工程,过度捕捞[25]和环境污染[1]等人类活动也会对渔业生物群落产生影响。因此,渔业生物群落结构受自然因素和人为因素的综合控制,而后者的评估还有待与相同海区同步的渔业捕捞数据和环境调查数据等历史资料进行对应对比分析。
致谢:样品采集由“北斗”号全体船员协助完成,谨致谢忱。
[1] 罗秉征.河口及近海的生态特点与渔业资源[J].长江流域资源与环境,1992,1(1):24-30.
LUO B Z.Ecological characteristics and fishery resources of the Yangtze River estuary and adjcent sea[J].Resources and Environment in the Yangtze Vally,1992,1(1):24-30.
[2] 金显仕,单秀娟,郭学武,等.长江口及其邻近海域渔业生物的群落结构特征[J].生态学报,2009,29(9):4761-4772.
JIN X S,SHAN X J,GUO X W,et al.Community structure of fishery biology in the Yangtze River estuary and its adjacent waters[J].Acta Ecologica Sinica,2009,29(9):4761-4772.
[3] 蒋 玫,沈新强.长江口及邻近水域夏季鱼卵、仔鱼数量分布特征[J].海洋科学,2006,30(6):92-97.
JIANG M,SHEN X Q.Abundance distributions of pelagic fish eggs and larval in the Changjiang River estuary and vicinity waters in summer[J].Marine Sciences,2006,30(6):92-97.
[4] 蒋 玫,沈新强,陈莲芳.长江口及邻近水域春季鱼卵仔鱼分布与环境因子的关系[J].海洋环境科学,2006,25(2):37-44.
JIANG M,SHEN X Q,CHEN L F.Relationship between with abundance distribution of fish eggs,larvae and environmental factors in the Changjiang estuary and vicinity waters in spring[J].Marine Environmental Science,2006,25(2):37-44.
[5] 刘守海,王金辉,刘材材,等.长江口水域夏季鱼卵和仔稚鱼年间变化[J].生态学报,2015,35(21):7190-7197.
LIU SH,WANG J H,LIU C C,et al.Inter-annual variation in pelagic fish egg,larval and juvenile assemblages during summer in the Yangtze River estuary,China[J].Acta Ecologica Sinica,2015,35(21):7190-7197.
[6] 刘淑德,线薇薇,刘 栋.春季长江口及其邻近海域鱼类浮游生物群落特征[J].应用生态学报,2008,19(10):2284-2292.
LIU S D,XIAN W W,LIU D.Characteristics of ichthyoplankton assemblages in Yangtze estuary and adjacent waters in spring[J].Chinese Journal of Applied Ecology,2008,19(10):2284-2292.
[7] 徐兆礼,袁骐,蒋 玫,等.长江口鱼卵和仔、稚鱼的初步调查[J].中国水产科学,1999,6(5):63-64.
XU Z L,YUAN Q,JIANG M,et al.An investigation of fish eggs,larvae and juveniles in the Changjiang estuary[J].Journal of Fishery Sciences of China,1999,6(5):63-64.
[8] 孙鹏飞,戴芳群,陈云龙,等.长江口及其邻近海域渔业资源结构的季节变化[J].渔业科学进展,2015,36(6):8-16.
SUN P F,DAI F Q,CHEN Y L,et al.Seasonal variations in structure of fishery resource in the Yangtze River estuary and its adjacent waters[J].Progress in Fishery Sciences,2015,36(6):8-16.
[9] 史赟荣,晁 敏,全为民,等.2010年春季长江口鱼类群落空间分布特征[J].中国水产科学,2011,18(5):1141-1151.
SHI Y L,CHAO M,QUAN W M,et al.Spatial variation in fish community of Yangtze River estuary in spring[J].Journal of Fishery Sciences of China,2011,18(5):1141-1151.
[10] 李建生,程家骅.长江口水域主要渔业生物资源状况的分析[J].南方水产科学,2005,1(2):21-25.
LI J S,CHENG J H.Dynamic analysis of fishery biology resources of trawl in waters of the Yangtze River estuary[J].South China Fisheries Science,2005,1(2):21-25.
[11] 李建生,李圣法,程家骅.长江口近海秋季鱼类资源的年变化分析[J].海洋渔业,2008,30(2):120-125.
LI JS,LI SF,CHENG J H.Analysis on the annual variations of fish resources in the offshore water of Yangtze estuary in autumn[J].Marine Fisheries,2008,30(2):120-125.
[12] 单秀娟,金显仕.长江口近海春季鱼类群落结构的多样性研究[J].海洋与湖沼,2011,42(1):32-40.
SHAN X J,JIN X S.Diversity of fish community structure in the spring-time offshore waters at the Yangtze River estuary[J]. Oceanologia Et Limnologia Sinica,2011,42(1):32-40.
[13] 张 波,金显仕,唐启升.长江口及邻近海域高营养层次生物群落功能群及其变化[J].应用生态学报,2009,20(2):344-351.
ZHANG B,JIN X S,TANGQ S.Functional groups of high trophic level communities in adjacent waters of Changjiang estuary[J].Chinese Journal of Applied Ecology,2009,20(2):344-351.
[14] 郑元甲,陈雪忠,程家骅,等.东海大陆架生物资源与环境[M].上海科学技术出版社,2003.
ZHENG Y J,CHEN X Z,CHENG J H,et al.Biological resources and environment of continental shelf of the East China Sea[M].Shanghai Scientific&Technical Publishers,2003.
[15] 程家骅.伏季休渔制度实践的回顾之二:伏季休渔制度效果的实践验证[J].中国水产,2008(7):18-21.
CHENG J H.Review on the implement of summer moratorium of marine fishing II:Verification of the implement effect of summer moratorium of marine fishing[J].China Fisheries,2008(7):18-21.
[16] 程家骅,严利平,林龙山,等.东海区伏季休渔渔业生态效果的分析研究[J].中国水产科学,1999(4):81-85.
CHENG J H,YAN L P,LIN L S,et al.Analyses on the fishery ecological effect of summer close season in the East China Sea region[J].Journal of Fishery Sciences of China,1999(4):81-85.
[17] PIANKA E R. Ecology of the agamid lizard Amphibolurus isolepis in Western Australia[J].Copeia,1971:527-536.
[18] HU H C,XIAN WW.The environment effect on fish assemblage structure in waters adjacent to the Changjiang(Yangtze)River estuary(1998-2001)[J].Chinese Journal of Oceanology and Limnology,2009(27):443-456.
[19] WILHM J L.Use of biomass units in Shannon's formula[J].Ecology,1968:153-156.
[20] 李 涛,张秀梅,张沛东,等.山东半岛南部近岸海域渔业资源群落结构的季节变化[J].中国海洋大学学报:自然科学版,2011,41(1-2):41-50.
LI T,ZHANG X M,ZHANG P D,et al.Seasonal variation on community structure of fishery resources in the coastal waters of southern Shandong Peninsular[J]. PeriodicaL of Ocean University of China(Natural science edition),2011,41(1-2):41-50.
[21] 李因强,陈国宝.我国海洋鱼类群落研究现状[J].现代渔业信息,2008,23(1):16-21.LI Y Q,CHEN G B.Research status of marine fish communities in Chinese Sea area[J].Modern Fisheries Information,2008,23(1):16-21.
[22] SHANNON C E,WEAVER W.The mathematical theory of communication[M].Illinois:University of Illinois Press,1959.
[23] MARGALEF D R.Information theory in ecology[M].Society for General Systems Research,1958.
[24] PIELOU E C.Ecological Diversity[M].New York:Wiley,1975.
[25] 金显仕,邓景耀.莱州湾渔业资源群落结构和生物多样性的变化[J].生物多样性,2000,8(1):65-72.
JIN X S,DENG J Y.Variations in community structure of fishery resources and biodiversity in the Laizhou Bay,Shandong[J].Chinese Biodiversity,2000,8(1):65-72.
[26] CLARKE K,Ainsworth M.A method of linking multivariate community structure to environmental variables[J].Marine Ecology-Progress Series,1993(92):205-205.
[27] 单秀娟,孙鹏飞,金显仕,等.黄海典型断面渔业资源结构的季节变化[J].水产学报,2013,37(3):427-435.
SHAN X J,SUN P F,JIN X S,et al.Seasonal variations of fishery resource structure in the sections of the southern Yellow Sea[J].Journal of Fisheries of China,2013,37(3):427-435.
[28] JIN X,TANG Q.Changes in fish species diversity and dominant species composition in the Yellow Sea[J].Fisheries Research,1996,26(3):337-352.
[29] 唐启升,贾晓平,郑元甲.中国专属经济区海洋生物资源与栖息环境[M].北京:科学出版社,2006.
TANG Q S,JIA X P,ZHENG Y J.Marine living resources and habitats in the exclusive economic zone of China[M].Beijing:Science Press,2006.
[30] 李圣法,程家骅,李长松,等.东海中部鱼类群落多样性的季节变化[J].海洋渔业,2005,27(2):113-119.
LI SF,CHENG J H,LI C S,et al.Seasonal changes on fish community diversity in the middle part of the East China Sea[J].Marine Fisheries,2005,27(2):113-119.
[31] 李显森,于振海,孙 珊,等.长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠[J].应用生态学报,2013,24(8):2353-2359.
LI X S,YU Z H,SUN S,et al.Ecological niche breadth and niche overlap of dominant species of fish assemblage in Yangtze River estuary and its adjacent waters[J].Chinese Journal of Applied Ecology,2013,24(8):2353-2359.
[32] 单秀娟,庄志猛,金显仕,等.长江口及其邻近水域大型水母资源量动态变化对渔业资源结构的影响[J].应用生态学报,2011,22(12):3321-3328.
SHAN X J,ZHUANG Z M,JIN X S,et al.Effects of macro-jellyfish abundance dynamics on fishery resource structure in the Yangtze River estuary and its adjacent waters[J].Chinese Journal of Applied Ecology,2011,22(12):3321-3328.
[33] 邓景耀,赵传絪.海洋渔业生物学[M].北京:中国农业出版社,1991.
DENG J Y,ZHAO C Y.Marine Fisheries Biology[M].Beijing:China Agricultural Press,1991.
[34] 马克平,刘玉明.生物群落多样性的测度方法[J].生物多样性,1994,2(4):231-239.
MA K P,LIU Y M.A method for measuring the diversity of biological communities[J].Chinese Biodiversity,1994,2(4):231-239.
[35] 陈云龙,单秀娟,戴芳群,等.东海近海带鱼群体相对资源密度,空间分布及其产卵群体的结构特征[J].渔业科学进展,2013,34(4):8-15.
CHEN Y L,SHAN X J,DAI F Q,et al.Relative stock density and distribution of hairtail Trichiurus lepturus and its spawning stock structure in coastal waters of the East China Sea[J].Progress in Fishery Sciences,2013,34(4):8-15.
[36] 胡 芬.东海刺鲳资源状况及数量分布[J].海洋渔业,2006,28(2):99-104.
HU F.The resource status and biomass distribution of wart perch Psenopsis anomala in the East China Sea[J].Marine Fisheries,2006,28(2):99-104.
[37] 徐兆礼,陈佳杰.小黄鱼洄游路线分析[J].中国水产科学,2009,16(6):931-940.
XU Z L,CHEN J J.Analysis on migratory routine of Larimichthy polyactis[J].Journal of Fishery Sciences of China,2009,16(6):931-940.
[38] 陈大刚.渔业资源生物学[M].北京:中国农业出版社,1997.
CHEN D G.Fishery Resources Ecology[M].Beijing:China Agricultural Press,1997.
[39] COLVOCORESSES J,MUSICK J.Species associations and community composition of middle Atlantic Bight continental-shelf demersal fishes[J]. Fishery Bulletin,1984,82(2):295-313.
[40] 李圣法,程家骅,严利平.东海大陆架鱼类群落的空间结构[J].生态学报,2007,27(11):4377-4386.
LI SF,CHENG JH,YAN L P.Spatial structures of fish communities on the continental shelf of the East China Sea[J].Acta Ecologica Sinica,2007,27(11):4377-4386.
[41] 于海成.长江口及邻近海域鱼类群落结构分析[D].青岛:中国科学院海洋研究所,2008.
YU H C.Fish community structure in the Changjiang estuary and adjacent waters[D].Qingdao:Institute of Oceanology,Chinese Academy of Sciences,2008.
Biological community of fishery resources in the Yangtze River Estuary and adjacent sea areas in the summer of 2015
TANG Chang-sheng1,2,3,ZHANG Fang1,3,*,FENG Song1,3,DAI Fang-qun4,WANG Yan-tao1,3
(1.Key Laboratory of Marine Ecology and Environmental Sciences,Institute of Oceanology,Chinese Academy of Sciences,Qingdao 266071,China;2.University of Chinese Academy of Sciences,Beijing 100049,China;3.Functional Laboratory of Marine Ecology and Environmental Sciences,Qingdao National Laboratory for Marine Science and Technology,Qingdao 266071,China;4.Laboratory of Fishery Resources and Ecosystems,Yellow Sea Fishery Research Institute,Chinese Academy of Fishery Sciences,Qingdao 266071,China)
Community structure analysis is one of the research focus topics in the Yangtze River Estuary and adjacent sea areas.The survey data in June can well valuate the fishery resources in the early stage of fishing moratorium,which is significant to assess the implementation effect from the summer moratorium for marine fishing.So the species composition,dominant species,community diversity characteristics and community structure of fishery species in the Yangtze River Estuary and adjacent sea areas were analyzed based on the data collected from the bottom trawl surveys in June,2015.The results showed that there were 93 fishery species,which were mainly demersal fish and crustaceans.The average catch weight was relatively high,with fish species occupying the largest proportion.These were dominant species:yellow croaker(Larimichthys polyactis),anchovy(Engraulis japonicus)and largehead hairtail(Trichiurus lepturus),indicating that the dominant species replacement had occurred compared with history data in the same period.The relatively high diversity of the community was found in the nearshore zone of Zhoushan and Yushan fishing ground,as well as the offshore zone of the Yangtze River Estuary fishing ground.While relatively low diversity of the community was found in the offshore zone of Zhoushan fishing ground and the southern part of Dasha fishing ground.The fishery biological communities could be divided into groups by the cluster analysis and the single factor similarity analysis:nearshore zone around the 20 m depth contour,offshore zone of the Yangtze River Estuary fishing ground and the southern part of Dasha fishing ground between 20 m and 50 m depth contour and the central and marginal parts of Zhoushan fishing ground and Yushan fishing ground between 50 m and 75 m depth contour.Similarity percentage analysis showed that the discriminating species among different groups were yellow croaker,anchovy,largehead hairtail and Japanese butterfish(Psenopsis anomala).Seasonal migration of the dominant species driven by temperature and water depth might be the major reason for changes of community structure.
Yangtze River Estuary;summer;fishery biology;community structure
S 931
A
1004-2490(2017)05-0490-10
2016-09-08
国家基金委 -山东省联合基金项目(U1406403);中国科学院战略性先导科技专项(A类)(XDA11020305);国家自然科学基金重点项目(41230963)
汤昌盛(1989-),男,博士研究生,从事海洋生态学研究。E-mail:t-ch-sh@163.com
张 芳,副研究员。E-mail:zhangfang@qdio.ac.cn
