干扰及林龄影响下迪庆州云杉老龄林生态系统碳储量动态
2017-10-16冯源朱建华肖文发皇宝林毕艳玲温庆忠邓喜庆
冯源,朱建华*,肖文发,皇宝林,毕艳玲,温庆忠,邓喜庆
干扰及林龄影响下迪庆州云杉老龄林生态系统碳储量动态
冯源1,2,朱建华1,2*,肖文发1,2,皇宝林3,毕艳玲3,温庆忠3,邓喜庆3
1. 中国林业科学研究院森林生态环境与保护研究所//国家林业局森林生态环境重点实验室,北京 100091;
2. 南京林业大学南方现代林业协同创新中心,江苏 南京 2100373;3. 云南省林业调查规划院,云南 昆明 650051
老龄林是重要的森林碳库,研究老龄林碳储量长期变化对评价老龄林碳源和碳汇功能和量化区域尺度森林生态系统碳循环具有重要的意义。基于云南省迪庆自治州森林资源规划设计调查数据、样地数据和迪庆州造林、采伐、灾害等统计数据,运用林业碳收支模型(CBM-CFS3)模拟并预测了2005—2020年云南省迪庆州区域尺度云杉(Picea likiangensis)老龄林(过熟林)的生物量、死亡有机质(包括枯落物、枯死木和土壤有机碳)以及生态系统碳储量及其动态变化。结果表明,干扰情景下,2005—2020年迪庆州云杉老龄林的生物量、死亡有机质和生态系统碳储量范围分别为3.98~4.73 Tg、5.41~7.28 Tg和9.44~12.01 Tg,且均呈逐渐增长趋势。模拟期间,云杉老龄林的生物量碳密度和生态系统碳密度均呈减少趋势,其中生物量碳密度平均值为106.40 Mg∙hm-2,生态系统碳密度平均值为255.56 Mg∙hm-2;死亡有机质碳库碳密度呈增加趋势,平均值为149.16 Mg∙hm-2。研究结果显示,迪庆州云杉老龄林生态系统碳储量动态受林分生长、成熟林为过熟林和干扰三方面影响;其中自然生长导致生态系统碳储量增加0.51 Tg,成熟林进阶导致生态系统碳储量增加2.75 Tg,而采伐干扰造成生态系统碳储量损失1.14 Tg。建议未来森林经营中将老龄林每年采伐总面积控制在1.9×103hm2∙a-1以内,以保证老龄林生态系统碳储量趋于稳定,避免老龄林转变为碳源。
林龄;人为干扰;固碳;森林经营
老龄林地上生物量与地下生物量碳储量极高(Suchanek et al.,2004),是森林生态系统重要的碳库组分,也是木材产品的主要来源(Carey et al.,2001)。近期研究表明,老龄林是全球生态系统中重要的碳汇(Luyssaert et al.,2008),其生物量和土壤有机质碳储量都可能增加(Tan et al.,2011;Wharton et al.,2016;Zhou et al.,2006),而另外一些研究则表明老龄林固碳能力下降(Taylor et al.,2014)。因此,老龄林生态系统碳储量动态仍然具有不确定性,研究老龄林生态系统长期碳储量动态对于估算区域尺度森林生态系统碳收支、评价森林清除温室气体和减缓气候变化的作用具有极重要的意义(Dai et al.,2013;Ford et al.,2017)。
目前在区域尺度开展老龄林长期碳储量变化研究存在两个难点:第一,已有研究通常将老龄林视为静止对象,并未考虑中长期研究过程中老龄林的位置和面积均可能发生变化;第二,由于区域尺度上森林生态系统碳储量动态受到干扰影响的情况非常普遍(Pregitzer et al.,2004;Seedre et al.,2015),因此研究长期老龄林生态系统碳动态时,不仅需要考虑老龄林位置和面积的变化,还需要考虑干扰对老龄林碳储量的影响。已有研究中通量观测技术多用于评价老龄林一年或多年间生态系统净碳交换量,却不能将干扰和林分生长对老龄林生态系统碳动态的影响区分开来,更不能预测未来老龄林碳储量变化趋势。本文以云南省迪庆州云杉林(Picea likiangensis)老龄林作为研究对象,基于森林资源规划设计调查数据、森林干扰数据和加拿大林业碳收支模型(CBM-CFS3)模拟和预估2005—2020年老龄林碳动态。研究目的在于探索老龄林生态系统碳储量长期变化规律以及区分生长和干扰两个因素分别对老龄林碳储量的影响,对评价老龄林碳源和碳汇功能和区域尺度森林碳循环具有重要意义,有助于提出合理的生态系统碳管理决策并为森林可持续发展提供参考。
1 研究地区与研究方法
1.1 研究区概况
云南省迪庆藏族自治州(98°35′~100°19′E,26°52′~29°16′N)位于滇、川、藏三省区交汇处的横断山脉腹地,总面积为2.39×104km2。境内地势北高南低,垂直气候和立体生态环境特征明显。年平均气温4.7~16.5 ℃,年极端最高气温25.1 ℃,最低气温-27.4 ℃;年日照平均时数为 1740~2190 h;年降雨量600~1000 mm。境内水热差异显著,根据热量的水平分布,迪庆州大部分地区位于温带—寒温带气候区,少部区域处于北亚热带气候区。云南省森林资源规划设计调查数据显示,2005年迪庆州林业用地面积为1.62×106hm2,森林覆盖率高达73.9%,是云南省森林植被类型最丰富的区域之一;其中云杉林面积为1.55×105hm2,占迪庆州森林总面积10.4%,是迪庆州主要森林类型之一(图1)。迪庆州云杉林的特点是以成、过熟林为主,且二者起源均为天然林。其中过熟林面积为3.66×104hm2,占云杉林总面积的 23.6%,它对云杉林生态系统碳动态甚至是区域植被生态系统碳收支都有重要影响。本研究将迪庆州云杉过熟林(林龄>120 a)视为老龄林(Seedre et al.,2015;Soloway et al.,2017),林分特征如表1所示。
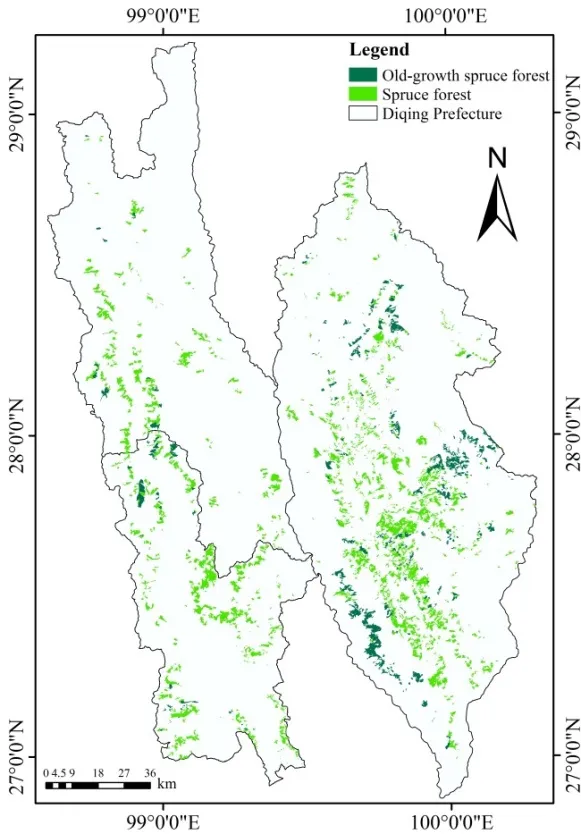
图1 2005年迪庆州云杉及其老龄林分布图Fig. 1 Distribution of spruce forests and its old-growth forest of Diqing in 2005
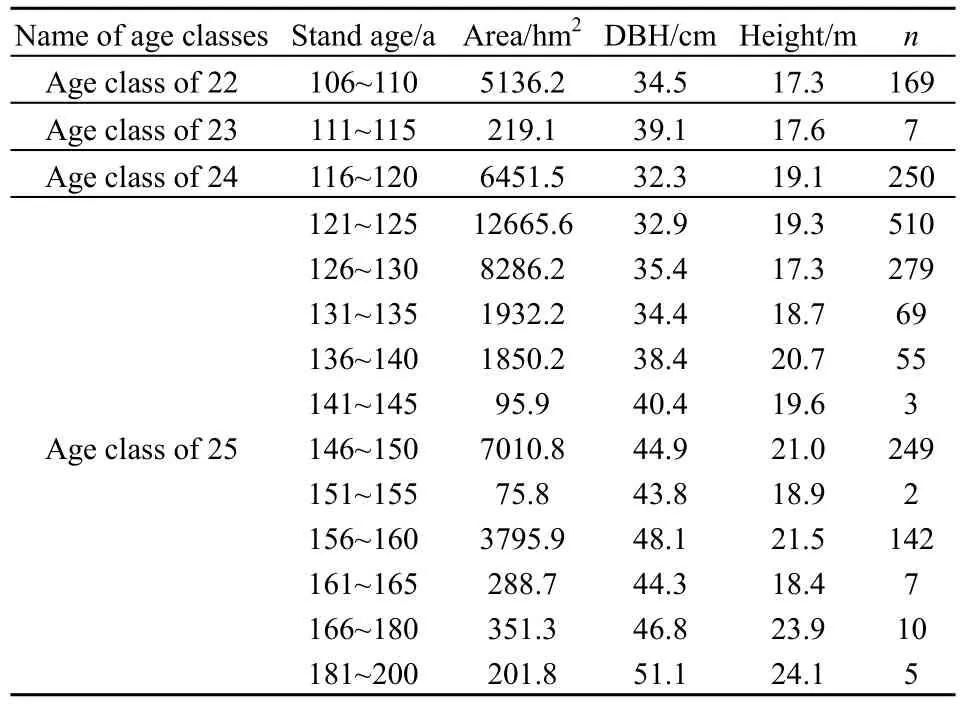
表1 迪庆州云杉老龄林特征Table 1 Description of old-growth spruce forest in Diqing
1.2 CBM-CFS3模型介绍
加拿大林务局开发的碳收支模型(Operationalscale carbon budget model of the Canadian Forest Sector,CBM-CFS3)是加拿大国家森林碳监测、计量和报告系统(NFCMARS)的核心模块(Kurz et al.,2009;Kull et al.,20142-3)。CBM-CFS3 模型采用“清单+变化法”估算森林生态系统碳储量动态,在考虑自然和人为干扰影响下评估林分尺度到国家尺度的森林生态系统碳储量变化。该模型是政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)推荐使用的碳计量模型,已被意大利(Pilli et al.,2013)、韩国(Kim et al.,2016)和墨西哥(Mascorro et al.,2016)等多个国家广泛应用于森林碳储量动态研究和森林管理决策,同时在中国云南(冯源,2014)、辽宁(甄伟等,2014)和河北(张滨等,2016)等地区也得到了初步应用。
CBM-CFS3模型的输入数据包括森林面积、优势树种、林龄、土壤类型、蓄积生长曲线以及干扰数据等;输出数据包含各碳库储量年变化、生态系统总体碳储量动态等结果。CBM-CFS3把生态系统分为生物量碳库及死亡有机质(DOM)碳库两大系统,生物量碳库包含地上生物量和地下生物量碳库;DOM 碳库包含枯落物、死木和土壤有机质 3个碳库(Kull et al.,2014)2-3。
1.3 输入数据及参数本地化
森林资源规划设计调查(简称“二类调查”)是驱动 CBM-CFS3运行的主要数据源。云南省每10年开展1次二类调查,最近一次于2017年初结束,因目前未能获得最新成果,故仍以 2005年二类调查数据为主要数据源。
将云杉老龄林的小班面积、林龄、蓄积、林木株数、起源、所处气候带以及云杉蓄积生长方程作为CBM-CFS3模型输入数据。CBM-CFS3模型提供约200个树种(或森林类型)的生物量估算参数,同时允许用户结合研究区域修改默认参数。本研究根据迪庆地区云杉老龄林特征对模型参数进行适当调整,其中蓄积生长方程来自冯源(2014)对云南省云杉长期林分蓄积生长的拟合结果;云杉生物量参数主要来源于中华人民共和国林业行业标准“立木生物量模型及碳计量参数——云杉(LY/T 2655—2016)”(国家林业局,2017)。根据模型的内置算法估算出模拟中使用的参数,其过程参照Boudewyn et al.(2007)2-37算法,估算出树干和地上生物量,再通过树枝、树叶等组分所占比例估算出各组分生物量参数(表2)。根据 Li et al.(2003)和 Smyth et al.(2010)3-7所提供的算法结合周涛等(2010)、黄锦学等(2010)对中国针叶林的评估(表 3),分别估算出地下生物量碳储量以及生物量周转比例,模型估算土壤碳库深度为1 m。由于云杉天然林灌木层及草木层(或下木层)生物量之和仅占云杉天然林生态系统的0.77%~2.5%(王金叶等,2000;马明东等,2007),因此不考虑灌木和草本碳储量动态。
1.4 干扰情景设置及研究对象的动态性
CBM-CFS3的优点在于可清楚地模拟干扰影响下森林生态系统碳储量的年际动态。此处干扰指自然及人为干预对森林固碳能力的扰动,包括火灾、病虫害等自然干扰,以及造林、采伐和毁林等经营措施。CBM-CFS3模型采用转移矩阵描述不同干扰类型影响下森林生态系统各碳库碳储量的转移方向及转移量(Kull et al.,2014)154。
森林干扰信息通过收集林地征占用项目、人工造林、森林火灾、病虫害与森林采伐等统计资料获得。由于森林干扰统计资料收集难度较大,本文通过迪庆州林业局仅得到 2005—2010年间森林干扰数据。通过提取云杉老龄林所受干扰面积、强度、发生位置及持续时间等信息可知,迪庆州云杉老龄林仅受到采伐干扰,主要为采伐强度为30%以下的疏伐,没有受到火灾、病虫害、毁林及其他人为干扰影响。2005年迪庆云杉老龄林的采伐面积最大(1.27×104hm2),随后采伐面积逐渐下降,2005—2010年累积采伐面积为3.62×104hm2(图2)。
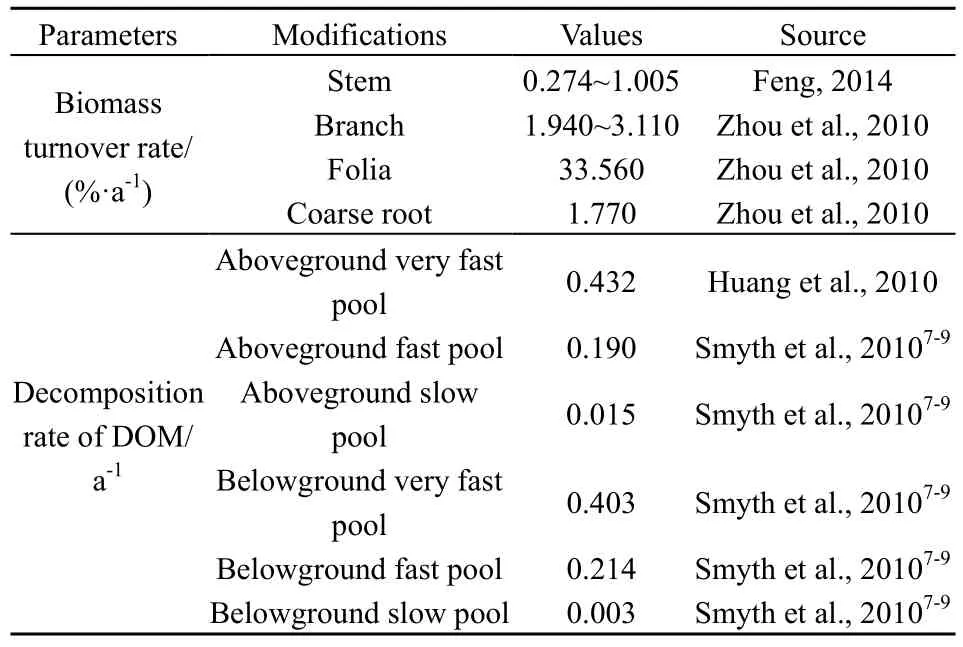
表3 CBM-CFS3模型生物量周转及分解参数Table 3 Parameters of biomass turnover and decomposition of CBM-CFS3 model
1.4.1 干扰情景设置
为量化 2005—2020年林龄效应及干扰对迪庆州云杉老龄林生态系统碳储量的影响,设置两种情景:(1)完全无干扰情景(ND),即假设2005—2020年迪庆州云杉林未受到任何干扰;(2)未来干扰情景(BS),假设2011—2020年间迪庆州云杉林所受干扰类型和强度是 2005—2010年历史干扰水平的循环。本文将BS情景视为未来迪庆州云杉林最可能实现的情景,ND情景则代表云杉老龄林的理想状态。根据两种情景对迪庆州云杉林进行整体设计,并假设干扰后森林类型不变。
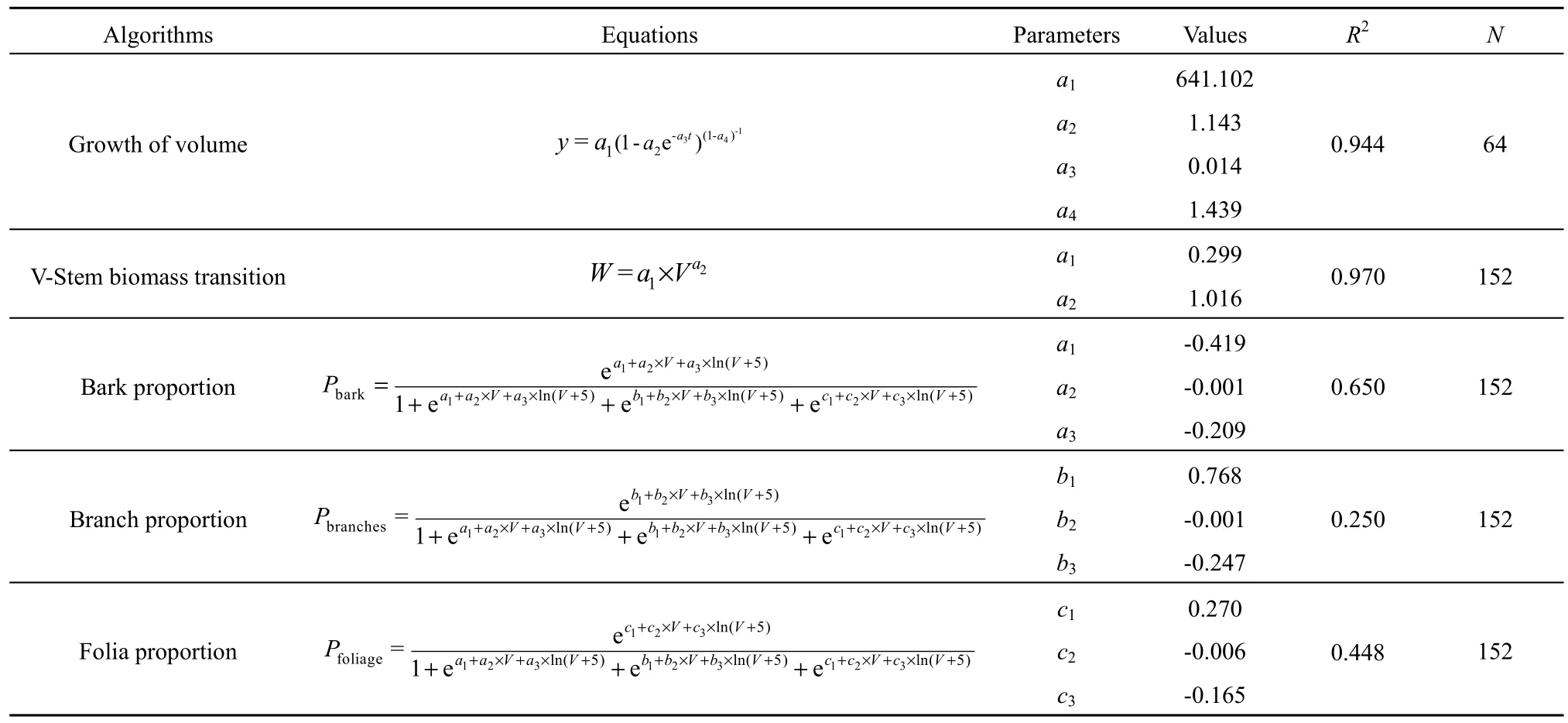
表2 CBM-CFS3模型生物量估算参数修改Table 2 Modified biomass estimating parameters in CBM-CFS3 model
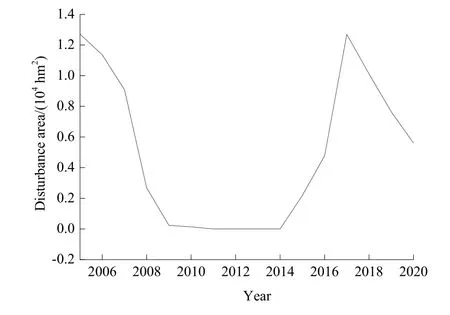
图2 2005—2020年BS情景的干扰面积Fig. 2 Disturbance area of BS scenario from 2005 to 2020
1.4.2 研究对象的动态性
由于模拟时间(共 16年)长于云杉龄级期限(本文设置为5年),随着林龄不断增大,成熟林将不断转化为老龄林,即云杉老龄林的面积和范围随时间动态可变。因此,需要考虑研究期间进阶为老龄林的成熟林。本文将模拟开始时(2005年)龄级在 25及其以上的过熟林视为老龄林(命名为“25龄级”),而在2010年、2015年以及2020年时,龄级为24、23和22的成熟林逐个进阶为老龄林;即2005—2020年间共有3个龄级的成熟林转化为老龄林,分别命名为“24龄级”、“23龄级”和“22龄级”。由于采伐强度小,假设采伐后的林分仍为老龄林(Burton et al.,1999),即模拟期间老龄林面积由于成熟林进阶而持续增加,并且不受干扰影响而减少。迪庆州云杉老龄林面积在 2005年、2010年、2015 年和 2020 年分别为 3.66×104、4.31×104、4.33×104和 4.84×104hm2(图 3)。
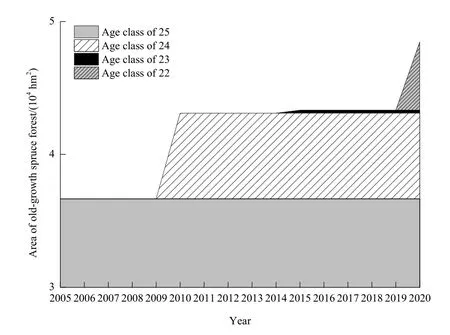
图3 2005—2020年迪庆州云杉老龄林面积Fig. 3 Area of old-growth spruce forest in Diqiang from 2005 to 2020
2 结果与分析
2.1 迪庆州云杉老龄林生态系统碳储量动态
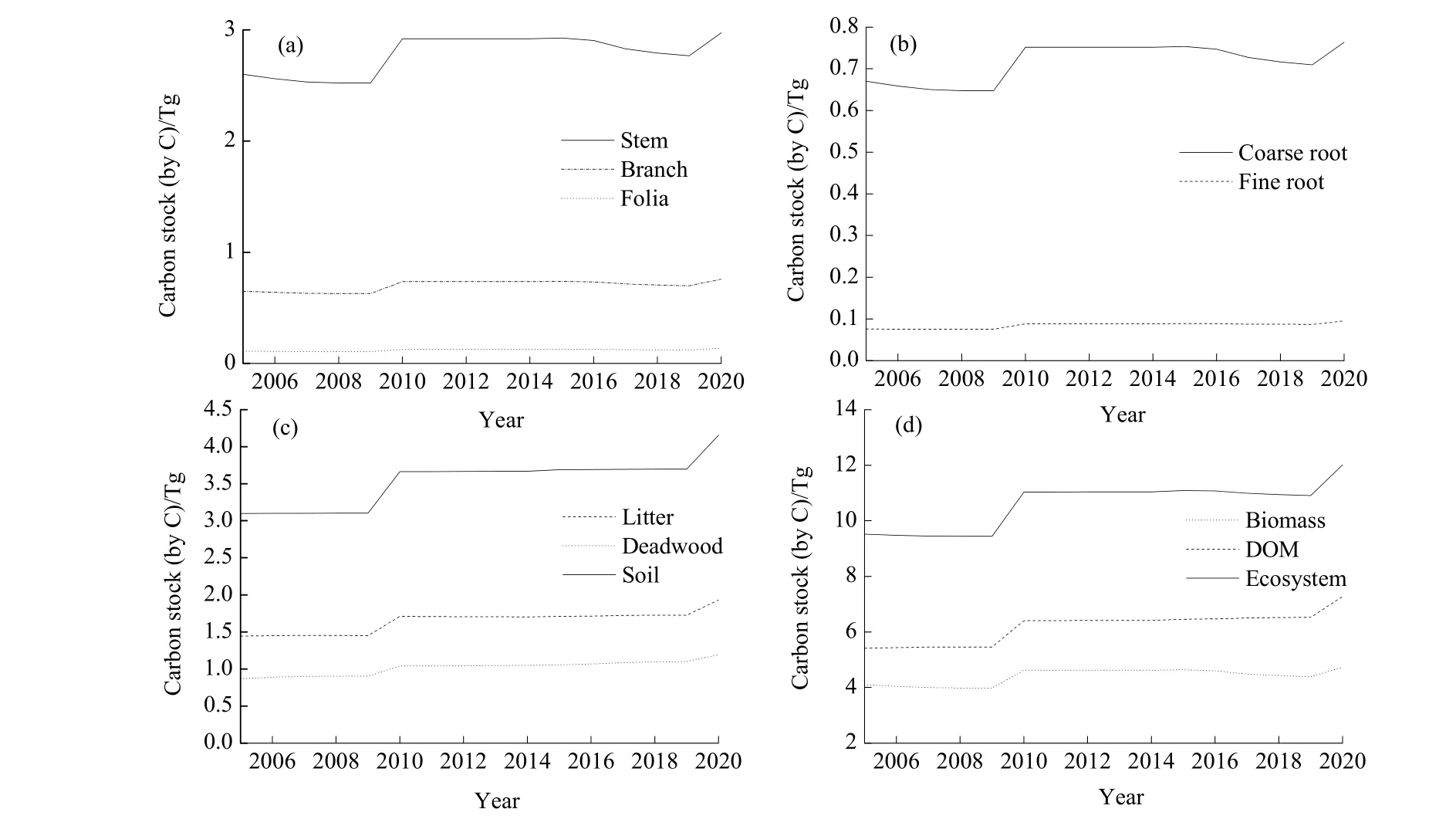
图4 2005—2020年BS情景下迪庆州云杉老龄林碳动态Fig. 4 Carbon dynamics of old-growth spruce forest in Diqing under BS scenario from 2005 to 2020(a)地上生物量 Above-ground biomass;(b)地下生物量 Below-ground biomass;(c)死亡有机质DOM;(d)生态系统 Ecosystem
BS情景下迪庆州云杉老龄林生物量及所有子碳库的碳储量变化趋势相同。连续采伐使得生物量碳储量在2005—2009年下降,2009年降到最低。24龄级在2010年进阶,引起碳储量显著增加并在2015年前保持稳定。而在干扰影响下,生物量碳储量在2016—2019年再次下降,最后在2020年由于22龄级进阶而达到最大值(图 4)。云杉老龄林生物量碳储量(以C计)变化范围为3.98~4.73 Tg,其中树干碳库碳储量最大,其次为树枝碳库,而树叶最小,三者碳储量范围分别为 2.52~2.97、0.63~0.76和0.11~0.14 Tg;地上生物量碳储量范围为 3.26~3.87 Tg;地下生物量碳储量范围为0.72~0.86 Tg。粗根碳储量远大于细根,二者碳储量范围分别为0.65~0.76 Tg和0.08~0.10 Tg。模拟期间,树干和粗根的碳储量波动最大,其他碳库变化相对较小。DOM碳储量范围为5.41~7.28 Tg,其中土壤有机质碳库的碳储量最大,其次为枯落物,而死木碳储量最小,三者碳储量范围分别为3.10~4.16、1.44~1.93 和 0.87~1.19 Tg。模拟期间,DOM及3个子碳库的碳储量均呈逐渐增加趋势,即2005年碳储量最小,2010年显著增加并在2020年达到最大值(图 4)。生态系统由生物量碳库及DOM碳库组成,生态系统碳储量范围为9.44~12.01 Tg,其变化趋势与生物量相同(图5)。ND情景下云杉老龄林生态系统碳储量在模拟期间逐渐增加,范围为9.62~13.15 Tg,并始终大于BS情景下云杉老龄林生态系统碳储量(图6)。
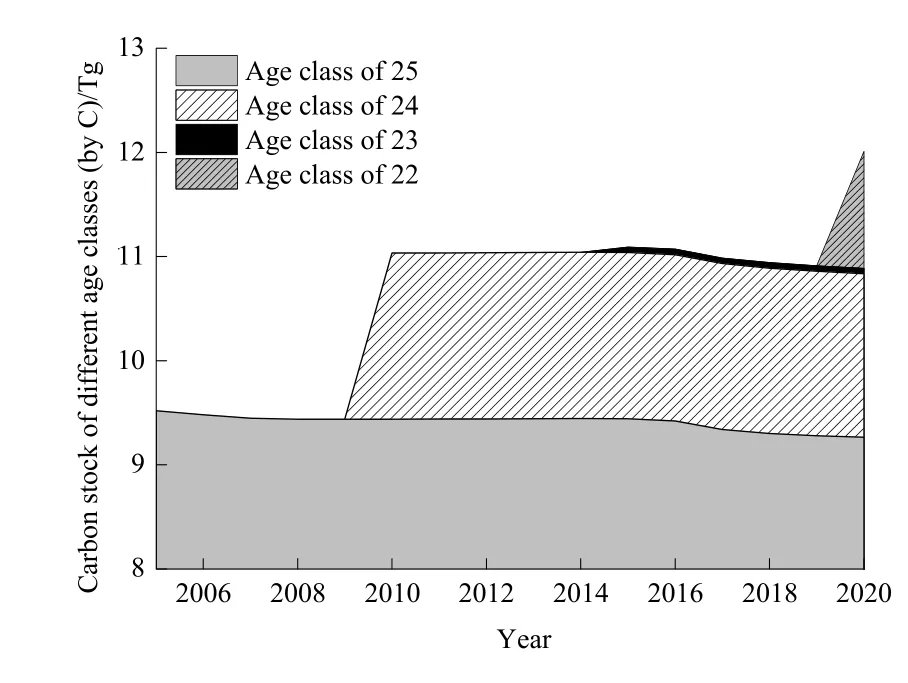
图5 BS情景下云杉老龄林生态系统碳储量Fig. 5 Carbon stock of old-growth spruce forest ecosystem under BS scenario
2.2 生长及采伐干扰对迪庆州云杉老龄林生态系统碳储量的影响
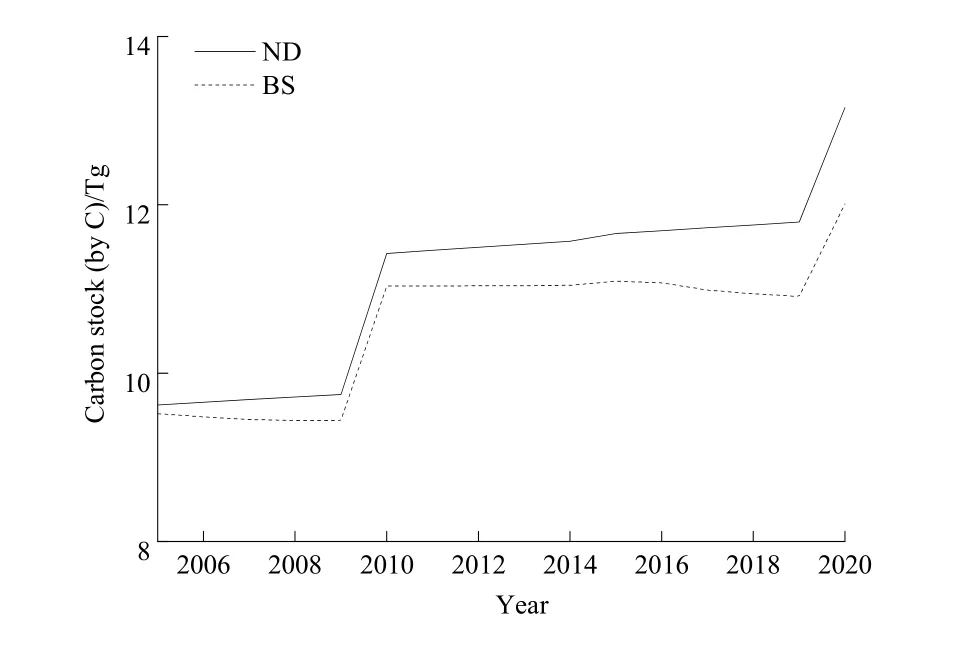
图6 ND及BS情景下云杉老龄林生态系统碳储量Fig. 6 Carbon stock of old-growth spruce forest ecosystem of ND and BS scenarios
迪庆州云杉老龄林的生长包含两方面:(1)成熟林不断进阶引起老龄林碳储量增加;(2)各龄级老龄林自然生长而增加的碳储量。前者由模拟结束时进阶成熟林的碳储量来表示。在2020年,BS情景由于24龄级、23龄级和22龄级进阶共造成老龄林生态系统碳储量净增2.75 Tg,在ND情景中为3.09 Tg(表4)。而老龄林自然生长则体现于模拟期间 ND情景下各龄级的碳储量逐渐增加,其中 25龄级和24龄级的碳储量增加最为明显,分别从9.62 Tg和1.64 Tg增加到10.07 Tg和1.70 Tg,说明在没有干扰影响下,25~23龄级由于生长而增加的碳储量共计0.51 Tg,而22龄级在2020年才进阶,还未有生长。
干扰效应表现为采伐使各龄级碳储量减少。模拟期间,BS情景下所有龄级碳储量均减小,其中25龄级和24龄级的碳储量分别从9.56 Tg和1.60 Tg减小到9.26 Tg和1.57 Tg,说明干扰造成的各龄级碳损失大于生长引起的碳增长量。与ND情景相比,干扰造成BS情景下云杉老龄林生态系统碳储量减少1.14 Tg(表4)。
2.3 迪庆州云杉老龄林生态系统碳密度动态
BS情景下2005—2020年迪庆州云杉老龄林生态系统碳密度平均值为255.56 Mg∙hm-2(表5),生物量平均碳密度为106.40 Mg∙hm-2,DOM平均碳密度为149.16 Mg∙hm-2;其中土壤有机质碳密度最大(85.03 Mg∙hm-2),其次为树干碳库(67.26 Mg∙hm-2)。模拟期间,云杉老龄林生物量所有碳库的碳密度呈减小趋势,其中树干、树枝和粗根碳密度降低幅度较大,而树叶和细根碳密度降低幅度较小;相比之下,DOM 碳库中枯落物和土壤碳密度均逐渐增大;而死木碳密度先增加,随后在 2011—2015年稍有下降,但在整个模拟期呈增大趋势。由于DOM碳密度增加量小于生物量碳密度的减少量,因此老龄林生态系统碳密度逐渐减小。ND情景下,2005—2020年云杉老龄林生态系统平均碳密度为267.46 Mg∙hm-2,所有碳库的碳密度均逐渐增加,并且生物量所有碳库的碳密度均大于BS情景,而所有DOM碳库的碳密度则略小于BS情景。

表4 生长及干扰对云杉老龄林生态系统碳储量的影响Table 4 Impacts of forest growth and disturbance on old-growth spruce forest ecosystem Tg (by C)
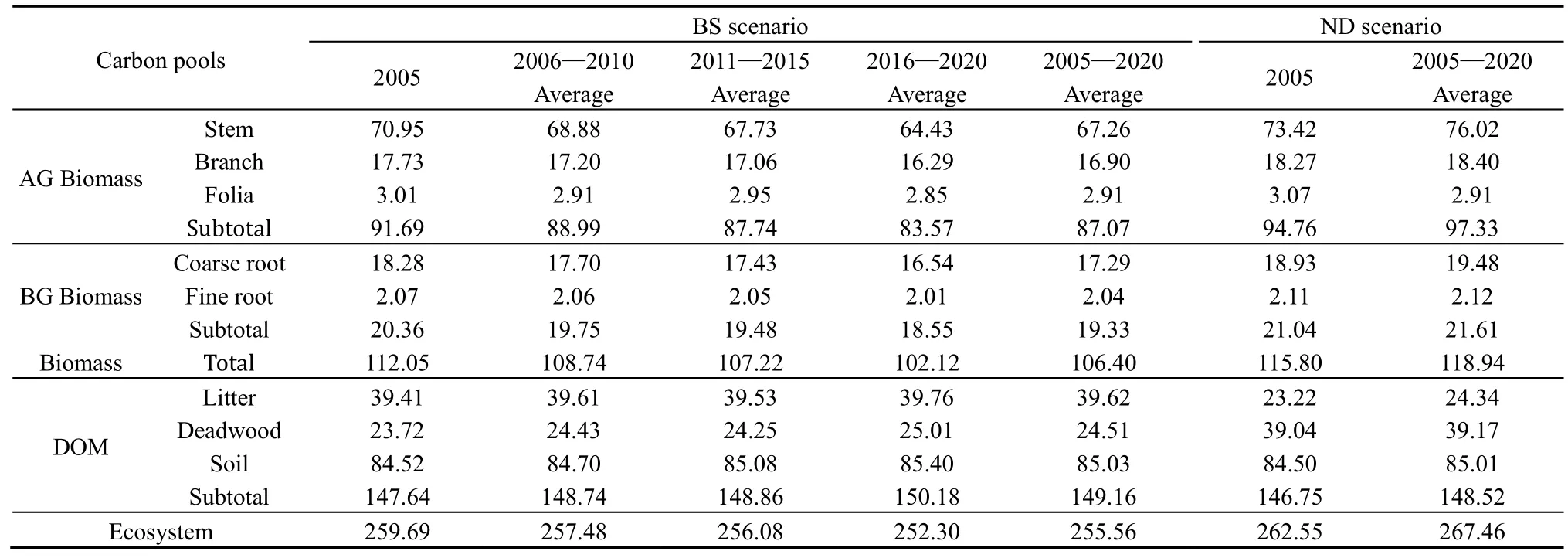
表5 2005—2020年云杉老龄林生态系统碳密度*Table 5 Carbon density of old-growth spruce forest ecosystem from 2005 to 2020 Mg∙hm-2
3 讨论
由于缺乏迪庆地区云杉老龄林碳储量数据,本研究结果只能与周边地区云杉老龄林(过熟林)研究结果进行对比(表 6)。2005年迪庆州云杉老龄林生物量碳密度(106.40 Mg∙hm-2)大于李虎等(2008)、任德智等(2016)和王金叶等(2000)报道的云杉过熟林生物量碳密度(83.93~97.13 Mg∙hm-2,),而小于彭守璋等(2011)和葛立雯(2013)等估算的祁连山北坡和西藏林芝地区过熟林生物量碳密度(117.10~144.52 Mg∙hm-2)。碳密度差异与数据来源、研究方法、生物量估算参数、林分特征、立地环境(如土壤类型)和研究尺度等因素具有密切关系。前人研究多采用样地调查与异速生长方程或生物量转换因子相结合的方法(葛立雯等,2013;任德智等,2016),研究范围为景观尺度,而本文则采用碳收支模型研究行政区内的云杉老龄林总体变化;在生物量估算参数方面,葛立雯等(2013)和任智德等(2016)分别以0.521、0.507作为云杉含碳率,而CBM-CFS3模型则以0.5作为含碳率参数。另外,不同地区对于云杉林龄划分标准不同,如彭守璋(2011)将祁连山地区青海云杉(Picea crassifolia Kom.)过熟林定义为林龄大于181 a的林分,而迪庆州云杉过熟林林龄主要在121~130 a之间,平均林龄远小于180 a。研究对象和研究方法的不同是造成结果差异的主要原因。
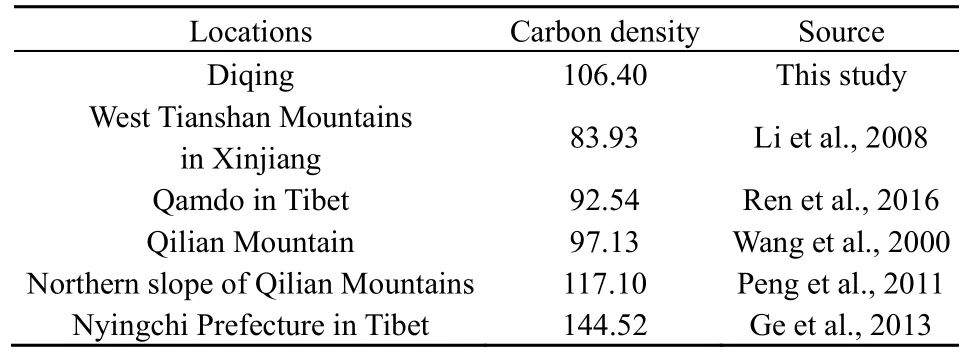
表6 云杉老龄林生物量碳密度对比Table 6 Comparison of biomass carbon density of old-growth spruce Mg∙hm-2
迪庆州云杉老龄林土壤碳密度在BS和ND两种情景下基本相同,而且模拟期间两种情景下土壤碳密度增加幅度相同(0.51 Mg∙hm-2)。据此推断,BS情景下土壤碳密度略高于ND情景是由于历史干扰不同而造成初始化结果稍有差异。研究期内,两种情景的土壤碳密度增加均来源于进阶成熟林,而采伐对老龄林土壤碳密度没有显著影响。这与Nave et al.(2010)得出的土壤总碳储量的变化与采伐没有显著关系的结论相吻合,而与短期内疏伐可增加土壤有机质碳储量(李云红等,2009;Christophel et al.,2013)或采伐后土壤碳储量降低(Kaarakka et al.,2016;Mushinski et al.,2017)的结论不同。采伐对森林土壤碳储量的影响与采伐方式、剩余物处理、立地特征及土壤类型有关,目前仍是研究热点之一,未来仍需对该方面开展深入研究。
2005—2020年云杉老龄林通过自然生长,生态系统碳储量增加0.51 Tg,而且所有碳库的碳储量均逐渐增加。这与 Seedre et al.(2015)和任德智等(2016)结论吻合:即使云杉林龄已经达到过熟林,其碳密度仍随着林龄增大而增大,彭守璋等(2011)进一步指出云杉生物量在 183年时达到最大值,因此,迪庆州云杉老龄林仍具有良好的固碳潜力。迪庆州云杉老龄林生态系统碳动态受林分生长、老龄林面积变化以及采伐干扰三方面影响。尽管林分生长量小于采伐干扰的碳损失(1.14 Tg),但模拟期间成熟林进阶引起的碳储量增加(2.75 Tg)弥补了干扰损失量,使得区域尺度老龄林碳储量有所增加。
如果不考虑模拟期间研究对象的动态性,那么25龄级的碳储量变化即为迪庆州云杉老龄林生态系统的碳储量动态。在2005—2020年云杉老龄林生态系统碳储量将从9.56 Tg减小到9.26 Tg(图4),即历史干扰水平将导致生态系统碳储量持续减少,增加云杉老龄林成为碳源的风险。因此,未来对森林经营进行合理规划、实施科学的碳管理策略是十分必要的。假设采伐仅影响干扰当年的生态系统碳储量,参考2007—2016年25龄级老龄林碳储量近于稳定的情况,在未来干扰类型和干扰强度不变情况下,将老龄林每年采伐面积控制在 1.9×103hm2∙a-1以内,可保证云杉老龄林生态系统碳储量趋于稳定。
随着模拟时间的增长,老龄林分布和面积始终是动态变化的,这是中长期老龄林碳动态模拟的一项挑战。区分林龄生长和采伐干扰对老龄林碳储量动态的影响是本研究的重要创新点。本文通过假设未来迪庆州云杉林整体干扰强度不变,预测出2012—2020年迪庆州云杉老龄林碳储量动态,但该情景假设具有两处不确定性:(1)BS情景是建立在采伐后土地利用类型不变的基础上进行的,因此可能高估了采伐后的林龄,继而高估了老龄林生态系统碳储量;(2)近些年来随着天然林保护力度加大,严格的采伐限额和禁伐措施使得云杉老龄林更少受到采伐影响,因此模拟结果可能高估了采伐对未来云杉老龄林碳储量的影响。
CBM-CFS3模型不能反映短期内气候因子变化对森林生产力的影响,因此该模型尚不能有效反映2010年和2015年迪庆州气象干旱对云杉老龄林生态系统碳动态的影响。气候因子变化可影响森林生态系统生产力(石兆勇等,2017),因此未来需要改进该模型对气候变化影响的估算,并且需围绕不同采伐类型、气候变化或自然灾害等干扰,假设更为合理的情景对老龄林生态系统碳动态进行深入研究。
4 结论
考虑了林龄和干扰对老龄林的影响,利用二类调查数据和 CBM-CFS3模型预估未来迪庆州云杉老龄林生态系统碳动态。结果表明,2005—2020年迪庆州云杉老龄林生态系统与生物量碳库的碳储量变化趋势相同,表现为先降低后增加再降低,而二者碳密度均表现为减小趋势;DOM 碳储量及碳密度均为增加趋势。模拟期间树木生长导致云杉老龄林生态系统碳储量(以C计)增加0.51 Tg,同时成熟林进阶导致碳储量增加2.75 Tg,大于干扰引起的碳储量降低量(1.14 Tg),表明迪庆州云杉老龄林生态系统碳动态受林分生长、老龄林面积变化以及采伐干扰三方面影响。未来将老龄林每年采伐面积控制在1.9×103hm2∙a-1以内,可保证云杉老龄林生态系统碳储量趋于稳定。
BOUDEWYN P, SONG X, MAGNUSSEN S, et al. 2007. Model-based,volume-to-biomass conversion for forested and vegetated land in Canada.Victroia [M]. Victoria, BC: Pacific Forestry Centre: 2-37.
BURTON P J, KNEESHAW D D, COATES K D. 1999. Managing forest harvesting to maintain old growth in boreal and sub-boreal forests [J].Forestry Chronicle, 75(75): 623-631.
CAREY E V, SALA A, KEANE R, et al. 2001. Are old forests underestimated as global carbon sinks? [J]. Global Change Biology, 7(4): 339-344.
CHRISTOPHEL D, SPENGLER S, SCHMIDT B, et al. 2013. Customary selective harvesting has considerably decreased organic carbon and nitrogen stocks in forest soils of the Bavarian Limestone Alps [J].Forest Ecology and Management, 305: 167-176.
DAI L M, JIA J, YU D P, et al. 2013. Effects of climate change on biomass carbon sequestration in old-growth forest ecosystems on Changbai Mountain in Northeast China [J]. Forest Ecology and Management,300(4): 106-116.
FORD S E, KEETON W S. 2017. Enhanced carbon storage through management for old-growth characteristics in northern hardwoodconifer forests [J]. Ecosphere, 8(4): 1-20.
KAARAKKA L, HYVÖNEN R, STRÖMGREN M, et al. 2016. Carbon and nitrogen pools and mineralization rates in boreal forest soil after stump harvesting [J]. Forest Ecology and Management, 377: 61-70.
KIM M, LEE W K, KURZ W A, et al. 2016. Estimating carbon dynamics in forest carbon pools under IPCC standards in South Korea using CBM-CFS3 [J]. iForest-Biogeosciences and Forestry, 10(1): 83-92.
KULL S J, RAMPLEY G J, MORKEN S, et al. 2014. Operational-scale carbon budget model of the Canadian forest sector (CBM-CFS3) version 1.2: user's guide [M]. Victoria, BC: Pacific Forestry Centre: 2-154.
KURZ W A, DYMOND C C, WHITE T M, et al. 2009. CBM-CFS3: a model of carbon-dynamics in forestry and land-use change implementing IPCC standards [J]. Ecological Modelling, 220(4): 480-504.
LI Z, KURZ W A, APPS M J, et al. 2003. Belowground biomass dynamics in the Carbon Budget Model of the Canadian Forest Sector: recent improvements and implications for the estimation of NPP and NEP [J].Canadian Journal of Forest Research, 33(1): 126-136.
LUYSSAERT S, SCHULZE E D, BÖRNER A, et al. 2008. Old-growth forests as global carbon sinks [J]. Nature, 455(7210): 213-215.
MASCORRO V S, COOPS N C, KURZ W A, et al. 2016. Attributing changes in land cover using independent disturbance datasets: a case study of the Yucatan Peninsula, Mexico [J]. Regional Environmental Change, 16(1): 213-228.
MUND M, KUMMETZ E, HEIN M, et al. 2002. Growth and carbon stocks of a spruce forest chronosequence in central Europe [J]. Forest Ecology and Management, 171(3): 275-296.
MUSHINSKI R M, BOUTTON T W, SCOTT D A. 2017. Decadal-scale changes in forest soil carbon and nitrogen storage are influenced by organic matter removal during timber harvest [J]. Journal of Geophysical Research: Biogeosciences, 122(4): 846-862.
NAVE L E, VANCE E D, SWANSTON C W, et al. 2010. Harvest impacts on soil carbon storage in temperate forests [J]. Forest Ecology and Management, 259(5): 857-866.
PILLI R, GRASSI G, KURZ W A, et al. 2013. Application of the CBM-CFS3 model to estimate Italy's forest carbon budget, 1995—2020 [J]. Ecological Modelling, 266(1): 144-171.
PREGITZER KS, EUSKIRCHEN ES. 2004. Carbon cycling and storage in world forests: biome patterns related to forest age [J]. Global Change Biology, 10(12): 2052-2077.
SEEDRE M, KOPÁČEK J, JANDA P, et al. 2015. Carbon pools in a montane old-growth Norway spruce ecosystem in Bohemian Forest:Effects of stand age and elevation [J]. Forest Ecology and Management, 346(2): 106-113.
SMYTH C E, TROFYMOW J A, KURZ W A, et al. 2010. Decreasing uncertainty in CBM-CFS3 estimates of forest soil carbon sources and sinks through use of long-term data from the Canadian Intersite Decomposition Experiment [M]. Victoria, BC: Pacific Forestry Centre: 3-9.
SOLOWAY A D, AMIRO B D, DUNN A L, et al. 2017. Carbon neutral or a sink? Uncertainty caused by gap-filling long-term flux measurements for an old-growth boreal black spruce forest [J]. Agricultural and Forest Meteorology, 233: 110-121.
SUCHANEK T H, MOONEY H A, FRANKLIN J F, et al. 2004. Carbon dynamics of an old-growth forest [J]. Ecosystems, 7(5): 421-426.
TAN Z H, ZHANG Y P, SCHAEFER D, et al. 2011. An old-growth subtropical Asian evergreen forest as a large carbon sink [J].Atmospheric Environment, 45(8): 1548-1554.
TAYLOR A R, SEEDRE M, BRASSARD B W, et al. 2014. Decline in net ecosystem productivity following canopy transition to late-succession forests [J]. Ecosystems, 17(5): 778-791.
WHARTON S, FALK M. 2016. Climate indices strongly influence old-growth forest carbon exchange [J]. Environmental Research Letters, 11(4): 4-16.
ZHOU G, LIU S, LI Z, et al. 2006. Old-growth forests can accumulate carbon in soils [J]. Science, 314(5804): 1417-1417.
冯源. 2014. 基于CBM模型云南普洱地区森林生态系统碳收支研究[D].北京: 中国林业科学研究院.
葛立雯, 潘刚, 任德智, 等. 2013. 西藏林芝地区森林碳储量, 碳密度及其分布[J]. 应用生态学报, 24(2): 319-325.
国家林业局. 2017. LY/T 2655—2016, 立木生物理模型及碳计量参数-云杉[M]. 北京: 中国标准出版社: 1-21.
黄锦学, 黄李梅, 林智超, 等. 2010. 中国森林凋落物分解速率影响因素分析[J]. 亚热带资源与环境学报, 5(3): 56-63.
李虎, 慈龙骏, 方建国, 等. 2008. 新疆西天山云杉林生物量的动态监测[J]. 林业科学, 44(10): 14-19.
李云红, 张彦东, 孙海龙. 2009. 采伐干扰对东北温带次生林土壤碳矿化和活性有机碳的影响[J]. 水土保持学报, 23(6): 139-143.
马明东, 江洪, 罗承德, 等. 2007. 四川西北部亚高山云杉天然林生态系统碳密度, 净生产量和碳贮量的初步研究[J]. 植物生态学报, 31(2):305-312.
彭守璋, 赵传燕, 郑祥霖, 等. 2011. 祁连山青海云杉林生物量和碳储量空间分布特征[J]. 应用生态学报, 22(7): 1689-1694.
任德智, 葛立雯, 王瑞红, 等. 2016. 西藏昌都地区森林植被碳储量及空间分布格局[J]. 生态学杂志, 35(4): 903-908.
石兆勇, 张晓龙, 肖莉, 等. 2017. 菌根组合类型对森林总初级生产力应对温度和降水变化的影响[J]. 生态环境学报, 26(3): 379-385.
王金叶, 车克钧, 蒋志荣. 2000. 祁连山青海云杉林碳平衡研究[J]. 西北林学院学报, 15(1): 9-14.
张滨, 张丽娜, 刘秀萍, 等. 2016. 河北省北部森林植被碳储量和固碳速率研究[J]. 中国生态农业学报, 24(3): 392-402.
甄伟, 黄玫, 翟印礼, 等. 2014. 辽宁省森林植被碳储量和固碳速率变化[J]. 应用生态学报, 25(5): 1259-1265.
周涛, 史培军, 贾根锁, 等. 2010. 中国森林生态系统碳周转时间的空间格局[J]. 中国科学: 地球科学, 40(5): 632-644.
Abstract: Old-growth forest is an essential carbon pool in forest ecosystem. To understand the function of old forest as a carbon sink or source, it is necessary to quantify the long-term carbon dynamics in old-growth forest ecosystem on regional scale. Supported by local forest inventory data, field measurement data, statistics data on afforestation, reforestation, harvesting and disturbances (forest fire and other damages) from Diqing Tibetan Autonomous Prefecture, we used a forest carbon budget model (CBM-CFS3) to simulate and project the carbon dynamics affected by disturbance scenarios in the old-growth spruce (Picea likiangensis) forest in Diqing from 2005 to 2020. The results showed that the carbon stocks increased from 3.98 to 4.73 Tg(by C) in biomass, from 5.41 to 7.28 Tg in dead organic matter (DOM, which include litter, dead wood and soil organic carbon), and from 9.44 to 12.01 Tg in total ecosystem from 2005 to 2020. The average carbon density in biomass and total ecosystem tended to decrease, which averaging as 106.40 Mg∙hm-2and 255.56 Mg∙hm-2during the period, respectively. But the carbon density in DOM, averaged as 149.16 Mg∙hm-2,tended to increase during the simulation period. Our simulation result also showed that carbon dynamics of old-growth spruce forest was obviously influenced by three factors: stand growth, mature-forest moved forward and harvesting disturbance. Positive effects including mature-forest moving forward (totally 2.75 Tg) and forest ageing (totally 0.51 Tg) increased the carbon stock of old-growth forest ecosystem. Harvesting, in contrast, had a negative effect and reduced carbon stock by 1.14 Tg during the period. If the disturbed area could be controlled as smaller than 1.9×103hm2∙a-1in the future forest management practices, it would effectively prevent the old-growth spruce forest ecosystem in Diqing from being a net carbon source.
Key words: forest age; anthropogenic disturbance; carbon sequestration; forest management
Disturbances and Ageing Affected Carbon Dynamics in Old-growth Spruce Forest in Diqing Prefecture
FENG Yuan1,2, ZHU Jianhua1,2*, XIAO Wenfa1,2, HUANG Baolin3, BI Yanling3,WEN Qingzhong3, DENG Xiqing3
1. Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry//State Forestry Administration Key Laboratory of Forest Ecology and Environment, Beijing 100091, China;2. Co-innovation Center for sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China;3. Yunnan Forest Inventory and Planning Institute, Kunming 65051, China
10.16258/j.cnki.1674-5906.2017.09.002
S718.5; X173
A
1674-5906(2017)09-1465-08
冯源, 朱建华, 肖文发, 皇宝林, 毕艳玲, 温庆忠, 邓喜庆. 2017. 干扰及林龄影响下迪庆州云杉老龄林生态系统碳储量动态[J]. 生态环境学报, 26(9): 1465-1472.
FENG Yuan, ZHU Jianhua, XIAO Wenfa, HUANG Baolin, BI Yanling, WEN Qingzhong, DENG Xiqing. 2017. Disturbances and ageing affected carbon dynamics in old-growth spruce forest in Diqing Prefecture [J]. Ecology and Environmental Sciences, 26(9):1465-1472.
国家林业局948项目(2012-4-78);中国清洁发展机制基金赠款项目(20130105)
冯源(1990年生),女,博士研究生,研究方向为生态系统管理及其服务功能变化。E-mail: flyh0901@163.com*通信作者:朱建华(1975年生),男,副研究员,博士,研究方向为气候变化与森林生态系统管理。E-mail: zhucool@caf.ac.cn
2017-07-20
