黄河三角洲养殖塘水-气界面CO2、CH4和N2O通量特征
2017-10-16宋红丽刘兴土文波龙
宋红丽,刘兴土*,文波龙
黄河三角洲养殖塘水-气界面CO2、CH4和N2O通量特征
宋红丽1,2,刘兴土2*,文波龙2
1. 山东省水土保持与环境保育重点实验室//临沂大学资源环境学院,山东 临沂 276005;
2. 中国科学院湿地生态与环境重点实验室//中国科学院东北地理与农业生态研究所,吉林 长春130102
随着经济的增长和人口的增加,沿海地区养殖业迅速发展,故养殖区水-气界面的CO2、CH4和N2O交换通量对大气中温室气体浓度的影响不容忽视。以黄河三角洲神仙沟(桩11)南部的养虾塘为研究对象,于2013年6月—2014年4月,采用漂浮箱-气相色谱法对养殖塘白天水-气界面CO2、CH4和N2O通量进行观测,并同步测定养殖塘部分环境因子。结果表明,黄河三角洲养殖塘 CO2、CH4和 N2O 的通量范围分别为-71.1403~7.2786、-0.0058~0.0165、-0.0020~0.0079 mg∙m-2∙h-1,均值分别为-36.3475、0.0059、0.0027 mg∙m-2∙h-1,整体上表现为从大气吸收CO2的汇和向大气排放CH4和N2O的源。养殖塘温室气体通量存在明显的季节变化特征,在温度较高的夏秋季,CH4和N2O排放通量较大。相关性分析结果表明,温度、盐度、SO42-、Cl-和磷水平对养殖塘温室气体具有一定的影响,且盐度与N2O通量、SO42-浓度与CO2通量之间的相关性达到显著水平。黄河三角洲养殖塘温室气体通量与自然水体和其他地区养殖塘存在较大差异,其中,地理位置的差异、养殖方式、杀菌剂的使用等环境因子和人为因素的作用显著。此外,养殖塘作为受人为干扰严重的水体,表现为多因子共同作用的结果,这一结论还有待于进一步的研究。研究结果对于综合评价水产养殖对区域气候变化的影响,深化人类活动对全球变暖的影响认识具有一定意义。
黄河三角洲;水-气界面;CH4;CO2;N2O
滨海湿地是由陆地、海洋和大气等多系统交互作用形成的动态平衡系统,具有高度的动态特征,是脆弱的生态敏感区,任何对生态系统和自然环境的干扰都会对其稳定性产生影响。随着经济的增长和人口的增加,人类对滨海湿地的围垦和开发强度越来越大。其中,滩涂养殖是滨海湿地开发利用的重要方式之一。近年来,海洋鱼类等资源的严重下降,为水产养殖注入新的动力。自 1970年起,随着世界对鱼类产品需求的剧增,养殖业以每年8.7%的速率高速增长,尤其是中国(Williams et al.,2010)。目前全球沿海地区养殖鱼类和贝类的产量比过去15年增加了1倍(Naylor et al.,2000)。据联合国粮食及农业组织数据显示,目前世界淡水养殖塘和海水养殖塘面积分别高达 87500 km2和23330 km2(Verdegem et al.,2009)。
此外,近年来以CH4、CO2和N2O浓度增加为主题的全球变暖问题是全世界关注的主要环境问题之一。自工业化以来,全球大气CO2、CH4和N2O浓度持续增加,2012年,全球大气 CO2、CH4和N2O 体积分数分别为 393.1 μL·L-1、1819 nL·L-1和325.1 nL·L-1,分别比工业化前增加 41%、160%和20%(IPCC,2013)。因此,减少温室气体的排放已成为当务之急。中国海岸线长达 18000 km,具有众多的河口、海湾、岛屿、浅海和滩涂湿地,为沿海养殖提供了优良的场所,这些区域海洋渔场面积高达 8.2×107hm2,其中浅海和滩涂面积为1.33×107hm2,故养殖区水-气界面的 CH4、CO2和N2O交换通量对于大气中温室气体浓度的影响不容忽视(杨平等,2013)。
黄河三角洲位于渤海湾南岸和莱州湾西岸,由黄河携带大量泥沙填充渤海淤积而成,为中国最大的三角洲,也是中国温带最广阔、最完整和最年轻的湿地。黄河三角洲地区主要的围垦利用类型包括水库港口建设、养殖、盐田以及农业。陈建等(2011)研究发现,1976—2008年,现代黄河三角洲人工湿地总面积增加了70.7倍,增加面积约为4.2×104hm2,与该区域盐田及鱼塘等人工湿地的不断增多有关。目前,关于黄河三角洲温室气体排放的研究仅在自然湿地区中开展(王玲玲等,2011;Sun et al.,2013),而关于黄河三角洲潮滩湿地遭受人类围垦后形成的水产养殖塘温室气体通量的研究还未见报道。为此,于2013年6月—2014年4月对黄河三角洲养殖塘CH4、CO2和 N2O通量特征及影响因素进行了初步研究。研究结果对了解水体温室排放特征,综合评价水产养殖对区域气候变化的影响,深化人类活动对全球变暖的影响认识具有一定意义。
1 材料与方法
1.1 研究区概况
黄河三角洲属暖温带季风性气候,具有明显的大陆性季风气候特点,雨热同期,四季分明,冷热干湿界限极为明显。春季干旱多风,常有春旱;夏季炎热多雨,高温高湿,时有台风侵袭。该区平均气温为12.1 ℃,年平均降水量为551.6 mm,70%的江水集中于7月和8月。试验所在地神仙沟(桩11)位于现黄河口北侧约30 km处,在1953—1960年的黄河入海口附近。1976年黄河由三角洲北部的钓口流路改为目前的清水沟流路入海。废弃神仙沟-钓口三角洲叶瓣位于现代黄河三角洲的北部。1976年黄河尾闾改道后,神仙沟-钓口三角洲叶瓣由原来快速向海口淤进转为向陆蚀退,严重威胁胜利油田的生产(王厚杰等,2010)。因此,目前神仙沟流路区主要为防止蚀退的人工海堤。此区域受人类活动干扰严重,除存在大量的油田外,盐田和养殖塘随处可见。研究所用养殖塘为海水养虾塘,通过水闸与大海连通,便于及时补充水分,调节养殖塘盐度,养殖密度约为7.5 ind·m-2。全池投撒饲料,初期每天投喂2~3次,后期每天投喂不少于3次,饲料日投放量控制在虾体重的3%~5%。
1.2 气体样品采集与分析
分别于2013年6月、8月、10月、12月和次年2月、4月采用浮箱-气相色谱法采集黄河三角洲养殖塘CH4、CO2和N2O气体。采样箱由PVC管制成(直径30 cm,高50 cm的圆筒),箱体四周及顶部用保温层包裹以确保样品采集过程中箱内的温度变化不大。外围装有漂浮装置和平衡装置以使采样箱能稳定地浮在水面上。采样箱内部装有监测箱内温度的传感器、采气用的硅胶管和三通阀及混合箱内气体的小风扇。同时,采样时为了确保箱内空气与外界隔绝,需将采样箱置于水面,使箱口浸入水中,浸入深度需根据采样时的风浪状况来定。采样点设在养殖塘的中间,将采样箱固定于小木船上,3个重复等间距直线排列。选择风速较小的晴天,于上午8:00开始,每隔2 h采集1次样品,即分别在8:00、10:00、12:00、14:00、16:00进行气体采集。样品采集采用50 mL注射器,在60 min时间段内每隔 20 min采集1次样品,所采集的气体样品装于250 mL气袋中(大连德霖气体包装有限公司)。样品采集后36 h内,在实验室用Agilent 7890气象色谱仪分析CH4、CO2和N2O气体浓度。
1.3 水样及环境因子的测定
采集气体样品的同时,监测气温以及水温的变化情况,同时采集采样箱体周围表层(0~10 cm)水样,装入500 mL聚乙烯瓶中带回实验室立即进行测定。水质盐分、总磷、阴离子(SO42-、Cl-)分别采用称重法、钼酸铵分光光度法、离子色谱法测定。
1.4 通量计算
水-气界面 CH4、CO2和 N2O排放通量采用以下计算公式:

式中,J 为气体通量(mg∙m-2∙h-1),dc/dt为采样时气体体积分数随时间变化的回归曲线斜率,M 为被测气体摩尔质量(g∙mol-1),P为采样点气压(Pa),T为采样时绝对温度(K),A为采样箱覆盖面积,V为箱内空气体积,V0、P0、T0分别为标准状态下的气体摩尔体积(mL∙mol-1)、空气气压(1013.25 hPa)和绝对温度(273.15 K)。浮箱内的气体浓度变化率通过对每次采集的3个气样所测的各气体含量进行线性回归分析获得,测量的浓度数据只有在线性回归系数R2>0.90时才视为有效。
1.5 数据分析
采用Origin 8.0和SPSS 18.0软件对数据进行作图、计算、相关分析和线性拟合分析。
2 结果与分析
2.1 CH4、CO2和N2O通量月变化特征
2013年6月—次年4月,黄河三角洲养殖塘CO2通量范围为-71.1403~7.2786 mg∙m-2∙h-1(图 1a),均值为-36.3475 mg∙m-2∙h-1,整体表现为吸收 CO2的汇。其中,2013年6月、8月、10月、12月及次年2月CO2的通量分别为-3.9585、-71.1404、-19.972、-62.8925 和-67.4000 mg∙m-2∙h-1,均表现为从大气中吸收CO2,且吸收峰出现在8月。仅2014年4月表现为排放 CO2的源,月均通量为 7.2786 mg∙m-2∙h-1。

图1 黄河三角洲养殖塘水-气界面CO2通量月变化(a)和季节变化(b)特征Fig. 1 Monthly variation (a) and seasonal variation (b) of CO2fluxes at water-air interface of aquaculture pond in the Yellow River estuary
2013年6月—次年4月,黄河三角洲养殖塘CH4通量范围为-0.0058~0.0165 mg∙m-2∙h-1(图 2a),年均值为 0.0059 mg∙m-2∙h-1,整体上表现为排放 CH4的源。6月和 12月 CH4通量分别为-0.0028 mg∙m-2∙h-1,-0.0058 mg∙m-2∙h-1,表现为从大气中吸收CH4的汇,且吸收峰出现在12月。2013年8月、10月及次年2月和4月CH4通量分别为0.0057、0.165、0.0067 和 0.0150 mg∙m-2∙h-1,表现为排放CH4的源,且排放峰出现在10月。
2013年6月—次年4月,黄河三角洲养殖塘N2O通量范围为-0.0020~0.0079 mg∙m-2∙h-1(图 3a),均值为0.0027 mg∙m-2∙h-1,整体上表现为排放N2O的源。2013年6—12月N2O排放特征呈先增加后减少的趋势,排放峰出现在 8月,通量值分别为 0.0029、0.0079、0.0051 和 0.0018 mg∙m-2∙h-1。次年 2 月和 4月 N2O 通量值分别为-0.0020 mg∙m-2∙h-1和 0.0006 mg∙m-2∙h-1,表现为N2O弱吸收汇和排放源。
2.2 CH4、CO2和N2O通量季节变化特征

图2 黄河三角洲养殖塘水-气界面CH4通量月变化(a)和季节变化(b)特征Fig. 2 Monthly variation (a) and seasonal variation (b) of CH4fluxes at water-air interface of aquaculture pond in the Yellow River estuary

图3 黄河三角洲养殖塘水-气界面N2O通量月变化(a)和季节变化(b)特征Fig. 3 Monthly variation (a) and seasonal variation (b) of N2O fluxes at water-air interface of aquaculture pond in the Yellow River estuary
黄河三角洲养殖塘水-气界面CH4、CO2和N2O具有显著季节变化特征(图1b、图2b、图3b)。具体而言,春、夏、秋、冬4个季节CO2通量的均值分别为-37.55、-19.97、-65.15 和 7.28 mg∙m-2∙h-1,除冬季表现为向大气排放CO2的源,其余3个季节均表现为从大气中吸收CO2的汇,吸收峰出现在冬季。CH44个季节通量的均值分别为0.0015、0.0165、0.0005 和 0.0150 mg∙m-2∙h-1,均表现为向大气中排放CH4的源,排放峰出现在秋季,最小值出现在冬季。冬季黄河三角洲养殖塘表现为从大气中吸收N2O的弱汇,而夏季和秋季则为向大气中排放N2O的强源,春季则为弱源,4个季节N2O通量的均值分别为-0.0001、0.0054、0.0051 和 0.0006 mg∙m-2∙h-1。
2.3 环境因子
观测时间段内气温和水温具有明显的季节变化特征(图4),呈现先减小后增加的趋势。气温的变化范围为(8.67±1.83)~(26.60±0.56) ℃,最低温和最高温分别出现在12月和6月。10 cm和20 cm水温的变化范围分别为(2.42±0.56)~(26.90± 0.35) ℃、(3.34±0.51)~(26.87±0.33) ℃。与气温的变化相似,最低温和最高温也分别出现在12月和6月。养殖塘SO42-质量浓度变化范围为2.95~3.97 g∙L-1(表1),不同月份之间含量存在波动。Cl-的变化趋势与SO42-基本一致,变化范围为 19.84~35.68 g∙L-1。总磷含量呈现“增加—减小—增加”的趋势,最大值出现在 4月(0.452 mg∙L-1)。盐度变化范围为23.10‰~57.56‰,最大值出现在4月。

图4 黄河三角洲养殖塘气温和水温季节变化特征Fig. 4 Seasonal variations of air temperature and water temperature in the Yellow River estuary
相关分析结果表明,养殖塘水-气界面CO2通量与水体 SO42-质量浓度之间存在显著正相关关系(r=0.829,P=0.042)(表2),与气温、水温和总磷则表现为正相关,但均未达到显著水平。N2O通量与气温、水温之间存在正相关性,与盐度、SO42-、Cl-、总磷之间存在负相关性,且与盐度之间的相关性达到了显著水平(r=-0.886,P=0.019)。CH4通量与所检测的环境因子之间均存在正相关,但均未达到显著水平。此外,通过对CO2、CH4和N2O通量与环境因子(气温、水温、盐度、SO42-浓度、Cl-浓度和总磷含量)进行二次线性拟合,结果表明,黄河三角洲养殖塘水-气界面 CO2通量与气温和水体 SO42-浓度、CH4通量与水体盐度存在较好的线性拟合关系,拟合方程分别为:Y=17.379+0.0448X1+4.4560X2( R2=-0.657)、 Y=3.7603+0.02326X1+0.00021X2(R2=0.69759)和 Y=35.78062-0.15368X1+0.02529X2(R2=-0.63091)。
2.4 相关研究结果的对比分析
黄河三角洲养殖塘CO2、CH4、N2O通量均值分别为-36.3475、0.0059、0.0027 mg∙m-2∙h-1(图 1,图2,图3),通过与已有研究的对比可以发现(表3):(1)黄河三角洲养殖塘CO2排放通量小于其他水体,南极莫愁湖(-70.8 mg∙m-2∙h-1)和团结湖(-36.9 mg∙m-2∙h-1)除外;(2)养殖塘水-气界面 CO2和 CH4排放通量高于自然水体,如在同一地区,江苏省中国水产科学研究院大宗淡水鱼养殖试验池塘 CO2(81.21 mg∙m-2∙h-1)和 CH4(1.86 mg∙m-2∙h-1)高于太湖CO2(35.145 mg∙m-2∙h-1)和CH4(0.095 mg∙m-2∙h-1)通量;(3)不同养殖塘水-气界面温室气体排放通量存在差异性。
3 讨论
3.1 CO2、CH4和N2O通量的时空变化特征
黄河三角洲养殖塘温室气体通量存在明显的月变化和季节变化特征(图1,图2,图3),8—12月从大气中吸收CO2,温度相对较高的6—10月出现 N2O排放通量的高峰期,与已有研究结果相似(Duan et al.,2005;Xing et al.,2006;杨平等,2013)。杨平等(2013)对闽江河口鱼虾混养塘温室气体监测结果显示,养殖塘温室气体通量具有明显的季节变化,表现为在温度相对高的月份(9—11月)出现CO2、CH4及N2O排放通量的高峰期,在温度相对低的月份(12月—次年1月)出现3种温室气体排放通量的低谷。吕迎春等(2007)认为叶绿素 a(Chl a)含量可能是富营养湖泊(水库)CO2季节变化的显著特征之一,夏季浮游植物生长旺盛,Chl a含量最高,因此浮游植物通过光合作用吸收大气中CO2,从而使水体中CO2欠饱和,成为大气CO2的汇;而冬季则为向大气中排放CO2的源。吕东珂(2009)对泥河水库水-气界面CO2通量的研究发现,影响CO2季节变化的主要因素有温度、水生植物、水位以及天气情况,从季节尺度上看,温度与通量之间存在一定的正相关性,即温度越高,通量越高,温度越低,通量就越低。CH4排放通量的季节变化除了与湖泊的植被和水深相关外,还与湖底淤泥的温度和光照强度密切相关(Duan et al.,2005)。
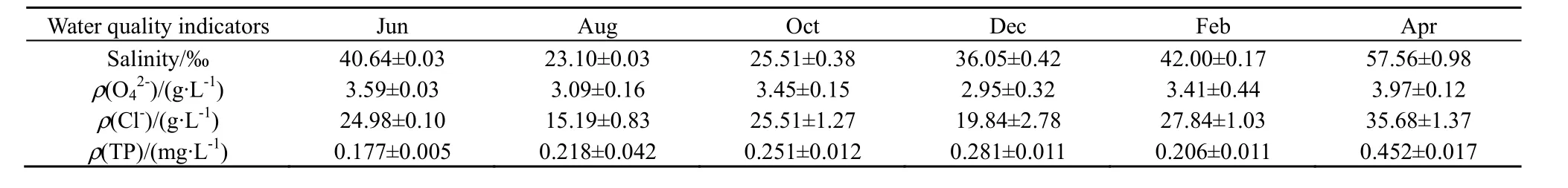
表1 黄河三角洲养殖塘水质变化特征Table 1 Variations of water quality in the aquaculture pond in the Yellow River estuary

表2 黄河三角洲养殖塘水-气界面温室气体通量与环境因子间的Sperman相关性分析Table 2 Sperman's correlation coefficient between greenhouse gases fluxes and environment factors in the aquaculture pond in the Yellow River estuary

表3 已有研究结果的对比分析Table 3 Comparative analysis of existing research results
本研究中养殖塘12月为从大气中吸收CH4的汇,主要原因在温度较低的月份,养殖塘中对虾的生命活动趋缓,对浮游植物的捕食强度降低,从而使得水体浮游植物光合作用释放的氧含量增多,水体变成好氧环境,不利于CH4的产生。值得注意的是,在温度较高的6月,养殖塘也表现为从大气中吸收CH4的汇,究其原因在于黄河三角洲对虾放养的最佳月份为5月中下旬,对虾放养时需在养殖塘灌入大量海水,对养殖塘的扰动较大,加之虾的投入也会对养殖塘造成扰动,从而增大水体氧含量。本研究6月温室气体的采样在上旬,此时水体处于不稳定状态,从而表现为吸收CH4的汇。黄河三角洲养殖塘春夏秋三季为排放N2O的源,仅冬季为从大气中吸收N2O的弱汇,主要与冬季水体盐度较高有关。相关分析结果表明,N2O通量与水体盐分存在极显著负相关关系,冬季高盐分含量抑制了N2O的产生。此外,温度对微生物硝化作用和反硝化作用也具有较大影响,温度相对较低的2月为从大气中吸收N2O的汇,4月则为N2O弱源。养殖塘为人为干扰严重的水体,在养殖过程中会人为添加各种外源物质,如饲料等,此外养殖塘中不存在大型沉水、挺水植被,故养殖塘水-气界面温室气体排放的季节变化与上述自然水体(如水库、湖泊)温室气体排放通量的季节变化存在一定的差异性。与上述研究不同的是,本研究中CO2通量虽与气温和水温均存在正相关性,但均未达到显著水平(表2),究其原因可能在于养殖塘受人为干扰严重,影响温室气体季节变化的因素与自然水体发生较大的变化。
水-气界面温室气体排放通量存在明显的空间变化(表3),主要表现在两个方面,一是不同地点的养殖塘温室气体排放通量存在差异性,其原因在于区域地理位置、采样方式、管理方式、养殖类型等方面具有差异性。地理位置的差异导致养殖塘水热条件不同,尤其是温度影响养殖塘微生物的活性,从而对温室气体排放产生影响。不同实验过程采集温室气体的时间间隔和方式存在一定的差异性。本研究中,黄河三角洲养殖塘水-气界面 CO2和CH4通量明显小于其他养殖塘,可能原因在于受光合作用和呼吸作用的影响,水气界面的交换都有明显的日变化,一般白天是大气的汇,夜晚是大气的源(李香华等,2005;嵇晓燕等,2006),受实验条件的限制,本研究中仅监测了白天水-气界面的温室气体,导致CO2和CH4通量明显小于其他养殖塘。养殖塘管理方式对温室气体通量的影响也较大,特别是饲养方式、杀菌剂的使用,肖启涛(2014)研究杀菌剂(CuSO4和HgCl2)添加对湖泊水体CO2、CH4、N2O浓度的影响发现,杀菌剂的添加能降低水体 pH,增加游离无机碳的浓度,显著增加水体CO2的浓度;Chen et al.(2016)研究发现,喂食的养殖塘从大气中吸收CO2,而不喂食的养殖塘则向大气中排放CO2,可能原因在于高水平的营养物质的输入,使得浮游植物大量繁殖,从而导致从大气中吸收CO2,排出O2。尤其是人工复合饵料进行水产养殖时,1 kg鱼的产出需要消耗70 g氮和800 g有机物,且这些氮和有机物的利用率很低,其中80%会通过鱼类的排泄进入养殖水体及底泥中,造成养殖水体碳、氮负荷的大量增加(刘浩等,2011),从而对养殖塘温室气体的排放产生显著的影响。目前,可以养殖的海鲜类型丰富,包括鱼类、虾类、蟹类、贝类等,不同的养殖类型对水质和养殖方式的要求不同,导致温室气体排放的不同。二是养殖塘和自然水体温室气体排放量不同。综上所示,养殖塘由于需要人为活动的干预,包括饲料的添加、杀菌剂的使用等,使得养殖塘水质条件(溶解氧、可溶性有机物、酸碱度、氧化还原电位等)存在很大差异性,特别是常年进行养殖活动的养殖塘,底泥的理化性质会发生很大的变化,导致养殖塘和自然水体的水-气界面温室气体的排放存在较大的差异性。
3.2 CO2、CH4和N2O通量与环境因子的关系
3.2.1 与温度的关系
总体而言,影响水-气界面温室气体排放的温度因子包括气温和水温两大方面,气温受太阳辐射等因素的影响,昼夜变化幅度较大;而水体由于比热较大,水温24 h内的变化较小,但水温受气温影响比较明显。温度对水-气界面温室气体排放的影响主要体现在以下3个方面:第一,温度是影响气体在水体中的溶解度的重要因素,在大气压一定的情况下,气体在水中的溶解度与温度呈反比,温度升高,溶解度降低,气体从水面逸出;温度降低,溶解度增加,气体从大气中进入水体(赵小杰等,2008),这也是热带地区,如闽江河口区养殖塘温室气体排放高于本研究结果的一个原因(表3)。第二,温度通过影响水体中水生生物的生长、繁殖以及水体中物质的分解、水生植物的光合作用及微生物活性间接影响水体中温室气体产生的过程和速率(兰晶,2015)。水温升高,水生生物活性增强,有利于水生生物的光合作用,加强对CO2的吸收,本研究黄河三角洲养殖塘表现为CO2的汇,部分原因在于样品的采集仅在温度相对较高的白天进行;CH4是在严格厌氧条件下由产甲烷菌作用于产甲烷底物的结果,研究表明产甲烷菌发挥作用的最适宜温度为25~30 ℃(Dunfield et al.,1993),环境中=温度的改变会对产甲烷菌活性产生影响,从而影响水-气界面 CH4的通量。第三,水温影响气泡的形成。水-气界面温室气体自然排放包括扩散和气泡两种方式(赵小杰等,2008)。Louis et al.(2000)认为,水温较高的沉积物中积累的甲烷量大于扩散到水中的量,导致甲烷过饱和而形成气泡,尤其是在浅水区,气泡对气体的传输是不容忽视的。Huttunen et al.(2003)对芬兰北部水库和湖泊温室气体通量特征的研究发现,浅水地区甲烷气泡的排放通量比水面扩散排放量高。
综上所述,温度对水-气界面温室气体的排放通量有较大的影响,在本研究中,气温和水温与CO2、CH4和N2O通量均呈正相关关系,但均未达到显著水平(表2),可能原因在于养殖塘受人为活动干扰较大,与自然水体相比,影响温室气体排放的因素较为复杂,是多因素综合作用的结果。
3.2.2 与盐度、SO42-和 Cl-的关系
本研究中,黄河三角洲养殖塘为海水养殖,其中养殖塘盐度、水体中SO42-和Cl-浓度范围分别为23.10~57.56‰、2.95~3.97 g∙L-1和 19.84~35.68 g∙L-1(表1)。相关分析结果表明,CO2和CH4与盐度、SO42-和 Cl-存在正相关关系,且 CO2通量与 SO42-相关关系达到显著水平(r=0.829,P=0.042);N2O与盐度、SO42-和Cl-均存在负相关关系,且N2O与Cl-浓度相关关系达到显著水平(r=-0.886,P=0.019)(表2)。N2O气体是硝化和反硝化过程作用的结果,盐分会直接影响硝化和反硝化速率,从而减少N2O的产生(Rysgaard et al.,1993),与本研究中N2O与盐度、SO42-和 Cl-均存在负相关关系的结果相一致。Bemhard et al.(2005,2007)的研究发现硝化潜力存在个盐度的最适值,在海水盐度下其速率会显著下降,本研究养殖塘为海水养殖,更加验证了盐度与水-气界面N2O通量存在负相关关系的结论。此外,盐分对水-气界面 CO2和 CH4通量也存在一定的影响,王纯(2015)模拟盐水入侵对闽江口淡水湿地温室气体产生与排放的影响时,发现低浓度的盐度作用下,盐水入侵对二氧化碳产生具有一定的促进作用,但未达到显著水平,与本研究中CO2与盐度之间存在正相关关系,但未达到显著水平的结果相一致。SO42-是一种重要的电子受体,可通过与甲烷产生菌竞争底物、提高甲烷产生环境的氧化还原条件,其还原过程中形成的硫化氨对甲烷产生菌的毒害作用等抑制了甲烷产生过程(王维奇等,2008)。相关研究也验证SO42-与CH4之间存在负相关关系(杨平等,2015),但在本研究中,相关分析结果表明,SO42-与CH4之间存在正相关关系,但未达到显著水平,其可能原因在于黄河三角洲养殖塘底泥中具有丰富的电子受体,在此情况下,SO42-与甲烷产生菌争夺电子受体的竞争关系便不成立。此时,电子受体丰富,CO2气体将成为主要的碳分解产物(Beal et al.,2009);且硫酸盐在土壤(底泥)呼吸中发挥着重要的作用,硫酸盐还原与CO2产生的比率是 1∶2 (Vandernat et al.,1998;Chambers et al.,2011)。3.2.3 磷营养水平
在水产养殖过程中,水体会接纳含有大量碳、氮的饲料及其他物质,这就强烈影响了水体中的碳、氮循环。Huttunen et al.(2002)的研究表明,如果有长期可利用的氮和磷供给,水体中碳循环以及初级生产可保证数十年甲烷和二氧化碳的大量排放。目前针对水体中溶解有机碳和氮含量对水-气界面温室气体影响的研究已有报道,王钰祺等(2011)对泥河水库水-气界面CO2通量特征的研究发现,CO2通量与溶解有机碳存在着正相关性;李红丽等(2012)对玉渡山水库生长季温室气体排放特征的影响研究发现,CO2和N2O通量与水体总氮呈显著正相关,CH4通量与水体总氮和水体铵态氮呈显著负相关。杨平等(2013)对闽江河口养殖塘水-气界面温室气体的研究发现,CO2、CH4和N2O通量与TN均存在显著正相关关系。针对水体磷营养水平对水-气界面温室气体排放通量的研究还鲜有报道,为此本研究分析了水体总磷水平对黄河三角洲养殖塘温室气体的影响,发现总磷与 CO2和CH4通量存在正相关关系,与N2O存在负相关关系,但均未达到显著水平(表2)。磷作为水体中生物的必需营养元素,支持着生物初级生产,从而间接影响温室气体的排放。
此外,水位、含氧量、水体pH、水面风速等因素均会影响水-气界温室气体通量(程炳红等,2012;赵小杰等,2008),尤其是养殖塘作为人为活动干扰较大的水体类型,影响其温室气体排放的影响往往是多因素共同作用的结果。由于野外试验的难操作性及样品的保存问题,本研究未对上述影响因子进行交互作用分析,将在下一步试验中加强。
4 结论
利用静态箱-气象色谱法对黄河三角洲养殖塘水-气界面CO2、CH4和N2O通量特征进行了研究,结果表明:(1)养殖塘水-气界面CO2、CH4和N2O通量具有明显的月变化和季节变化,整体上表现为从大气中吸收CO2的汇和向大气中排放CH4和N2O的源;(2)养殖塘为受人为活动干扰严重的湿地类型,影响其温室气体排放的因子,表现为多因子共同作用的结果,单个因素对温室气体排放的影响并不显著,仅盐度与N2O通量、SO42-与CO2通量之间的相关性达到显著水平;(3)黄河三角洲CO2、CH4和N2O通量与自然水体和其他养殖塘相比存在一定的差异性,可能与地理位置的差异性、养殖方式以及采样方式等因素有关。
BEAL E J, HOUSE C H, ORPHAN V J. 2009. Manganese- and iron-dependent marine methane oxidation [J]. Science, 325(5937):184-187.
BEMHARD A E, DONN T, GIBLIN A E, et al. 2005. Loss of diversity of ammonia-oxidizing bacteria correlates with increasing salinity in an estuary system [J]. Environmental Microbiology, 7(9): 1289-1297.
BEMHARD A E, TUCKER J, GIBLIN A E, et al. 2007. Functionally distinct communities of ammonia-oxidizing bacteria along an estuarine salinity gradient [J]. Environmental Microbiology, 9(6): 1439-1447.
CHAMBERS L G, REDDY K R, OSBORNE T Z. 2011. Short-term response of carbon cycling to salinity pulses in a freshwater wetland[J]. Soil Science Society of America Journal, 75(5): 2000-2007.
CHEN Y, DONG S, WANG Z, et al. 2015. Variations in CO2fluxes from grass carp Ctenopharyngodon idella aquaculture polyculture ponds [J].Aquaculture Environment Interactions, 8: 31-40.
CHEN Y, DONG S, WANG F, et al. 2016. Carbon dioxide and methane fluxes from feeding and no-feeding mariculture ponds [J].Environmental Pollution, 212: 489-497.
DIEM T, KOCH S, SCHWARZENBACH S, et al. 2008. Greenhouse gas emissions (CO2, CH4and N2O) from perialpine and alpine hydropower reservoirs [J]. Biogeosciences Discussions, 5(5): 3699-3736.
DUAN X, WANG X, MU Y, et al. 2005. Seasonal and diurnal variations in methane emissions from Wuliangsu Lake in arid regions of China [J].Atmospheric Environment, 39(25): 4479-4487.
DUNFIELD P, DOMONT R, MOORE T R. 1993. Methane production and consumption in temperate and subarctic peat soils: response to temperature and pH [J]. Soil Biology and Biochemistry, 25(3): 321-326.
HUTTUNEN J T, ALM J, LIIKANEN A, et al. 2003. Fluxes of methane,carbon dioxide and nitrous oxide in boreal lakes and potential anthropogenic effects on the aquatic greenhouse gas emissions [J].Chemosphere, 52(3): 609-621.
HUTTUNEN J T, VAISANEN T S, HELLSTEN S K, et al. 2002. Fluxes of CH4, CO2, and N2O in hydroelectric reservoirs Lokka and Porttipahta in the northern boreal zone in Finland [J]. Global Biogeochemical Cycles, 16(1): 3-17.
IPCC. 2013. Climate change 2013: the physical science basis[R].Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change.
LOUIS V L S, KELLY C A, DUCHEMIN E, et al. 2000. Reservoir surfaces as sources of greenhouse gases to the atmosphere: A global estimate[J]. Bioscience, 50(9): 766-775.
NAYLOR R L, GOLDBURG R J, PRIMAVERA J H, et al. 2000. Effect of aquaculture on world fish supplies [J]. Nature, 405(6790): 1017-1024.
REPO M E, HUTTUNEN J T, NAUMOV A V, et al. 2007. Release of CO2and CH4from small wetland lakes in western Siberia [J]. Tellus B,59(5): 788-796.
RYSGAARD S, RISGAARD-PETERSEN N, NIELSEN L P, et al. 1993.Nitrification and denitrification in lake and estuarine sediments measured by the15N dilution technique and isotope pairing [J]. Applied and Environmental Microbiology, 59(7): 2093-2098.
SUN Z G, WANG L L, TIAN H Q, et al. 2013. Fluxes of nitrous oxide and methane in different coastal Suaeda salsa marshes of the Yellow River estuary, China [J]. Chemosphere, 90(2): 856-865.
VERDEGEM M C J, BOSMA R H. 2009. Water withdrawal for brackish and inland aquaculture, and options to produce more fish in ponds with present water use [J]. Water Policy, 11(S1): 52-68.
VANDERNAT F J, MIDDELBURG J J, 1998. Seasonal variation in methane oxidation by the rhizosphere of Phragmites australis and Scirpus lacustris [J]. Aquatic Botany, 61(2): 95-110.
WILLIAMS J, CRUTZEN P J. 2010. Nitrous oxide from aquaculture [J].Nature Geoscience, 3(3): 143-143.
XING Y P, XIE P, YANG H, et al. 2006. The change of gaseous carbon fluxes following the switch of dominant producers from macrophytes to algae in a shallow subtropical lake of China [J]. Atmospheric Environment, 40(40): 8034-8043.
XING Y, XIE P, YANG H, et al. 2005. Methane and carbon dioxide fluxes from a shallow hypereutrophic subtropical Lake in China [J].Atmospheric Environment, 39(30): 5532-5540.
ZHU R, LIU Y, XU H, et al. 2010. Carbon dioxide and methane fluxes in the littoral zones of two lakes, east Antarctica [J]. Atmospheric Environment, 44(3): 304-311.
陈建, 王世岩, 毛战坡. 2011. 1976—2008年黄河三角洲湿地变化的遥感监测[J]. 地理科学进展, 30(5): 585-592.
程炳红, 郝庆菊, 江长胜. 2012. 水库温室气体排放及其影响因素研究进展[J]. 湿地科学, 10(1): 121-128.
嵇晓燕, 崔广柏, 杨龙元, 等. 2006. 太湖水-气界面 CO2交换通量观测研究[J]. 环境科学, 27(8): 1479-1486.
兰晶. 2015. 养殖塘水体温室气体的溶存与排放及其影响因素研究[D].武汉: 华中农业大学.
李红丽, 杨萌, 张明祥, 等. 2012. 玉渡山水库生长季温室气体排放特征及其影响因素[J]. 生态学杂志, 31(2): 406-412.
李香华, 胡维平, 杨龙元, 等. 2005. 太湖梅梁湾冬季水-气界面二氧化碳通量日变化观测研究[J]. 生态学杂志, 24(12): 1425-1429.
刘浩, 董莹, 田玲玲. 2011. 湖北省湖泊水产养殖对湖区生态系统的影响[J]. 湖北农业科学, 50(9): 1790-1792
吕迎春, 刘丛强, 王仕禄, 等. 2007. 贵州喀斯特水库红枫湖、百花湖p(CO2)季节变化研究[J]. 环境科学, 28(12): 2674-2681.
吕东珂. 2009. 泥河水库水-气界面CO2通量研究[D]. 哈尔滨: 东北林业大学.
王纯, 2015. 模拟盐水入侵对闽江口淡水湿地温室气体产生与排放的影响[D]. 福州: 福建师范大学.
王厚杰, 原晓军, 王燕, 等. 2010. 现代黄河三角洲废弃神仙沟-钓口叶瓣的演化及其动力机制[J]. 泥沙研究, (4): 51-60.
王钰祺, 吕东珂. 2011. 泥河水库秋季水-气界面CO2通量日变化特征及影响因子分析[J]. 森林工程, 27(2): 19-22.
王玲玲, 孙志高, 牟晓杰, 等. 2011. 黄河口滨岸潮滩湿地 CO2、CH4和N2O通量特征初步研究[J]. 草业学报, 20(3): 51-61.
王维奇, 曾从盛, 仝川. 2008. 闽江口芦苇湿地土壤甲烷产生与氧化能力研究[J]. 湿地科学, 6(1):60-68.
肖启涛. 2014. 太湖水-气界面温室气体 (CO2、CH4、N2O) 通量研究[D].南京: 南京信息工程大学.
杨平, 张子川, 杜威宁, 等. 2015. 河口盐地梯度下短叶茳芏沼泽湿地土壤间隙水溶性甲烷时空特征[J]. 环境科学, 36(10): 3633-3640.
杨平, 仝川, 何清华, 等. 2013. 闽江口鱼虾混养塘水-气界面温室气体通量及主要影响因子[J]. 环境科学学报, 33(5): 1493-1503.
赵小杰, 赵同谦, 郑华, 等. 2008. 水库温室气体排放及其影响因素[J].环境科学, 29(8): 2377-2384.
赵炎, 曾源, 吴炳方, 等. 2011. 三峡水库香溪河支流水域温室气体排放通量观测[J]. 水科学进展, 22(4): 546-553.
朱林, 车轩, 刘晃, 等. 2016. 团头鲂池塘养殖生态系统晒塘阶段温室气体排放通量分析[J]. 农业工程学报, 32(3): 210-215.
Abstract: With the growth of economics population, the aquaculture industry in coastal areas developed rapidly. Therefore, the influence of CO2, CH4and N2O fluxes from aquaculture areas can not be ignored. In this study, variations of CO2, CH4and N2O fluxes at water-air interface in the daytime were determined using a floating chamber- gas chromatography method in shrimp pond from June 2013 to April 2014 in the Yellow River estuary. Meanwhile, some of environmental factors were also measured. The results showed that the ranges of CO2, CH4and N2O fluxes were -71.1403~7.2786, -0.0058~0.0165, -0.0020~0.0079 mg∙m-2∙h-1,and the mean values were -36.3475, 0.0059, 0.0027 mg∙m-2∙h-1, respectively, indicating that the culture pond acted as an emission of CH4and N2O and consumption of CO2. Seasonal variations of greenhouse gases from culture pond were observed, the fluxes of CH4and N2O were large at high temperature. Correlation analysis results showed that temperature, salinity, SO42-, Cl-concentration and phosphorus had a certain influence on greenhouse gases fluxes, and the correlation between salinity and N2O flux, and SO42-and CO2flux reached significant level. The greenhouse gas fluxes of the Yellow River delta aquaculture pond were different from that of natural water and other aquaculture ponds, which might due to geographical differences, breeding way, use of fungicides and so on.In addition, culture pond was seriously affected by anthropogenic activities, and the greenhouse gases emission were complicated and might be controlled by the interactions of multiple controlling factors, which needed further research. The results of this study have certain significance for the comprehensive evaluation of the impacts of aquaculture influence on regional climate change, and deepening the impact of human activities on global warming.
Key words: Yellow River estuary; water-air interface; CH4; CO2; N2O
Greenhouse Gases Fluxes at Water-Air Interface of Aquaculture Ponds in the Yellow River Estuary
SONG Hongli1,2, LIU Xingtu1, WEN Bolong1
1. Shandong Provincial Key Laboratory of Water and Soil Conservation and Environmental Protection, Linyi University, College of Resources and Environment, Linyi 276005, China;2. Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences,Changchun 130102, China
10.16258/j.cnki.1674-5906.2017.09.014
X14; X16
A
1674-5906(2017)09-1554-08
宋红丽, 刘兴土, 文波龙. 2017. 黄河三角洲养殖塘水-气界面CO2、CH4和N2O通量特征[J]. 生态环境学报, 26(9):1554-1561.
SONG Hongli, LIU Xingtu, WEN Bolong. 2017. Greenhouse gases fluxes at water-air interface of aquaculture ponds in the Yellow River Estuary [J]. Ecology and Environmental Sciences, 26(9): 1554-1561.
国家重大科学研究计划项目(2012CD956100);国家青年基金项目(41601086);山东省自然科学基金博士项目(ZR2016DB05)
宋红丽(1986年生),女,讲师,主要从事湿地生态方面研究。E-mail: songhongli@lyu.edu.cn*通信作者。E-mail: lxtmxh@163.com
2017-03-27
