2014—2015年珠江春季禁渔前后浮游动物群落变化
2017-10-16高原赖子尼杨婉玲曾艳艺
高原,赖子尼,杨婉玲,曾艳艺
2014—2015年珠江春季禁渔前后浮游动物群落变化
高原,赖子尼*,杨婉玲,曾艳艺
中国水产科学研究院珠江水产研究所,广东 广州 510380
为探究珠江春季禁渔前后浮游动物群落变化特征,深入分析浮游动物群落对珠江春季禁渔的响应,为珠江流域的渔业资源和生物多样性保护积累基础数据,于2014—2015年3—6月,在西江上游至下游河口设置封开、肇庆和磨刀门3个采样点每年开展4次浮游动物及水质调查。分析了浮游动物的种类组成和优势种分布、浮游动物现存量和生物多样性指数的分布,探讨了浮游动物群落结构与环境因子的关系。调查期间共检出浮游动物88种(未包括无节幼体及桡足幼体)。萼花臂尾轮虫(Brachionus calyciflorus)、广布中剑水蚤(Mesocyclops leuckarti)、角突臂尾轮虫(Brachionus angularis)、跨立小剑水蚤(Microcyclops varicans)以及中华窄腹剑水蚤(Limnoithona sinensis)等种类在禁渔前、中、后均占有较大优势。浮游动物平均丰度在2579~2957 ind∙L-1范围内波动,3个采样点平均丰度变化趋势为:5月>4月>3月>6月,平均生物量变化趋势为:3月>5月>4月>6月。浮游动物生物多样性指数和均匀度指数从3—6月均总体呈现上升趋势,生物多样性指数和均匀度指数均值分别为2.85和0.91。PCA分析表明:封开浮游动物优势种类丰度与总氮和pH等环境因子显著相关;肇庆浮游动物优势种类丰度与总氮呈极显著正相关,与高锰酸盐指数和叶绿素a含量呈显著正相关,与溶解氧呈显著负相关;磨刀门浮游动物优势种类丰度与透明度呈显著正相关,与总磷呈显著负相关。可以看出,禁渔前后珠江干流浮游动物群落发生了明显变化,且受到营养盐等诸多因素的影响。
珠江;春季禁渔;浮游动物;种类组成
珠江是中国南方最大的河流,水域面积达2110 m2。西江作为珠江主干流,地处亚热带,气候温暖,自然生态条件独特,栖息有120多种不同类型的水生经济动物;同时其又是许多洄游性鱼类的洄游通道和繁殖场所(谭细畅等,2007)。根据近年来调查资料记载(内部资料),西江干流江段渔获物共采集到鱼类75种,隶属于10目、23科、65属。每年 4—6月,西江均有青、草、鲢、鳙、广东鲂等亲鱼溯江洄游集群产卵。其中,广东鲂是珠江的主要经济鱼类,属半洄游性鱼类,平常普遍分布于珠江及其支流的中、下游生长育肥,性成熟时,集群产卵。封开地处低纬地带,属亚热带气候,西江在此江段形成的青皮塘产卵场是西江广东段最重要的广东鲂产卵场之一(杨婉玲等,2011)。肇庆地处西江“黄金水道”的重要节点,西江肇庆段是珠江干流汇入珠三角河网水域的咽喉通道,是漂流性鱼卵、仔鱼进入珠三角河网的必经之地,历史上曾是采集天然“四大家鱼”鱼苗的重要区域(谭细畅等,2007)。珠江八大入海口门(以磨刀门为代表)地处亚热带,是咸淡水交汇的水域,受珠江径流、广东沿岸流和外海水的综合影响,生态环境独特,生物组成多样化,是洄游性鱼类的必经之地,渔业资源丰富(高原等,2008)。因此,本研究选取珠江干流上的封开、肇庆、磨刀门这3个具有代表性的水域作为研究对象。
浮游动物在水生态系统的结构和功能中起着重要的调控作用,它通过摄食控制浮游植物的数量,以调节水体生态平衡,同时又是许多经济水产动物的饵料,其数量变化可以直接影响渔业资源量(纪焕红等,2006)。而且,浮游动物的生物量也是对水域生态系统的鱼产力进行评估的重要指标(卢敬让等,1995)。
珠江流域自 2011年起开始实行禁渔期制度,至2015年已实施禁渔期制度5年。禁渔期制度的作用是多方面的,包括保护和恢复生物的自然资源及其多样性,保护主要物种的繁殖及子代的孵化生长,改善水生生物的资源结构,修复水域生态环境等(韩耀全等,2015)。以往的研究主要集中在禁渔对渔业资源(鱼、虾、蟹和鱼苗补充群体)群落结构及生物多样性的影响方面(施炜纲等,2005;刘凯等,2006;张敏莹等,2006;徐东坡等,2006;刘凯,2007;严利平等,2010;李跃飞等,2014),以及利用数学模型进行休渔效果的分析(严利平等,2006;刘汉武等,2012),而有关禁渔对浮游生物等渔业生态环境的影响研究未见报道。本研究对珠江春季禁渔前、中、后浮游动物的群落动态进行深入研究,探讨浮游动物群落对珠江春季禁渔的响应,为该水域的渔业资源和生物多样性保护积累基础数据。
1 材料与方法
1.1 研究水域概况
珠江水系是中国南方最大的水系,由西江、北江、东江及珠江三角洲诸河等4个水系所组成。西、北江在广东省三水县思贤滘以下,东江在广东东莞石龙镇以下,汇入珠江三角洲(杨婉玲等,2017),最后由崖门、虎跳门、鸡蹄门、磨刀门、横门、洪沥门、蕉门、虎门等8个口门流入南海,形成放射状的珠江出水口。西江是珠江流域内最大的水系,各水系在流域内的面积及所占比重:西江为353120 m2,占77.83%;北江为46710 m2,占10.3%;东江为27040 m2,占5.96%;三角洲及入注三角洲诸河26820 m2,占5.19%。珠江水系水资源较丰富,年径流量为 3.36×1011m3,出海河川径流量为3.26×1011m3,其中西江 2.38×1011m3,北江 3.85×1010m3,东江 2.29×1010m3,珠江三角洲 2.56×1010m3。珠江流域大部分地区属南亚热带,气候温和,多年平均气温在 20 ℃以上。雨量充沛,多年平均降雨量为1200~2200 mm。
1.2 调查时间及采样点
2014—2015年3—6月,在珠江春季禁渔前、中、后于珠江干流上设置封开(FK)、肇庆(ZQ)、磨刀门(MDM)3个采样点(图 1),进行浮游动物调查及水质监测。
1.3 研究方法
浮游动物定性、定量样品的采集和分析参考《淡水浮游生物研究方法》(章宗涉等,1995)。

图1 采样点位图Fig. 1 Location of sampling site
(1)原生动物和轮虫类:定性样品用25号(64µm)浮游生物网在水面按“∞”形移动拖取3~5 min,实验室内鉴定种类。定量样品于每个采样点用采水器采集表层水样1 L,现场加入福尔马林溶液以固定(4%体积比),带回实验室静置沉淀,浓缩至 10~20 mL,取0.1 mL浓缩水样于计数框内在10×20放大倍数下镜检2片计数原生动物数量,取1 mL浓缩水样于计数框内在10×10放大倍数下镜检2片计数轮虫数量。
(2)枝角类和桡足类:定性样品用13号(113µm)浮游生物网在水面按“∞”形拖取采集,定量样品用5 L采水器采集表层水30 L经25号浮游生物网过滤浓缩,加入福尔马林溶液固定后带回实验室静置浓缩至20 mL,用1 mL浮游生物计数框在10×10放大倍数下全部计数。
采集浮游动物样品的同时使用便携式水质分析仪(YSI6920-0,USA)进行pH和溶解氧等现场水化指标的监测,水体透明度采用塞氏盘法测定。另取适量水样(约250 mL)带回实验室用水质流动分析仪(Skalar-SA1100,Holand)进行总氮、总磷等营养盐指标的检测分析,叶绿素a含量采用N, N-二甲基甲酰胺(DMF)法进行测定。
1.4 数据处理
(1)采用优势度(Y)计算浮游动物的优势种,公式如下:

式中,fi是该种在各站位中出现的频率,mi为第i种的丰度,M为总丰度,优势度Y≥0.02视为优势种(徐兆礼等,1995)。
(2)采用标准回归系数(β)衡量各优势种对浮游动物总丰度变化的影响程度(徐兆礼等,2005)。标准回归系数是相关变量间因果关系的一个统计量,是自变量和因变量间带有方向的相关系数。以浮游动物总丰度为因变量,各优势种丰度为自变量,采用逐步线性回归分析方法,通过t检验筛选出对浮游动物总丰度回归方差贡献显著的优势种,然后计算标准回归系数(陈希孺等,1987)。
(3)采用Shannon-Wiener多样性指数(H)和均匀度指数(J)进行浮游动物多样性特征的研究,公式如下:

式中,S为种数,ni为第i种的个体数,N为总个体数(Shannon et al.,1963;Pielou,1966)。
(4)相关性分析:选取浮游动物的多样性指数及均匀度指数与 pH、透明度、溶解氧、总氮、总磷、高锰酸盐指数、叶绿素a等环境因子,取自然对数后采用SPSS 19.0软件对浮游动物与环境因子进行Pearson相关性分析。
(5)非约束型排序方法中的主成分分析(PCA):运用Canoco 4.5软件完成浮游动物群落与环境因子的PCA分析。在进行PCA分析之前,先进行降趋对应分析(DCA),当DCA分析结果中排序轴梯度最大值小于3时选择PCA分析对浮游动物与环境因子间的关系进行进一步分析。
2 结果与分析
2.1 种类组成
调查期间共检出浮游动物 88种(未包括无节幼体及桡足幼体)。其中,封开、肇庆及磨刀门分别检出80、82、85种。总体上,3个采样点轮虫类种类最多,所占比例最大,桡足类其次,原生动物和枝角类比例相对较小(见表 1)。在封开,2014年禁渔之前(3月)共检出72种,禁渔期间(4月和5月)均检出73种,禁渔之后(6月)检出68种;2015年禁渔之前(3月)共检出71种,禁渔期间(4月和5月)分别检出72和73种,禁渔之后(6月)检出66种。在肇庆,2014年禁渔之前(3月)共检出77种,禁渔期间(4月和5月)分别检出73和71种,禁渔之后(6月)检出68种;2015年禁渔之前(3月)共检出75种,禁渔期间(4月和5月)分别检出72和71种,禁渔之后(6月)检出 67种。在磨刀门,2014年禁渔之前(3月)共检出74种,禁渔期间(4月和5月)分别检出72和76种,禁渔之后(6月)检出74种;2015年禁渔之前(3月)共检出70种,禁渔期间(4月和5月)分别检出71和74种,禁渔之后(6月)检出72种。总体上,2015年3个采样点的浮游动物均比2014年同期种类数少,2年调查禁渔前、中、后的浮游动物种类数无显著性差异。
2.2 优势种
2年调查禁渔前、中、后浮游动物主要优势种及优势度分布见表2。禁渔前(3月)3个采样点共有16种优势种,禁渔期间(4月和5月)3个采样点共有19种优势种,禁渔后(6月)3个采样点共有 17种优势种。萼花臂尾轮虫(Brachionus calyciflorus)、广布中剑水蚤(Mesocyclops leuckarti)、角突臂尾轮虫(Brachionus angularis)、跨立小剑水蚤(Microcyclops varicans)以及中华窄腹剑水蚤(Limnoithona sinensis)等种类在禁渔前、中、后均占有较大优势。
调查发现,中华窄腹剑水蚤在磨刀门占有较大优势,但旋回侠盗虫(Strobilidium gyrans)却在磨刀门无明显优势(表2)。
通过逐步线性回归,进一步从优势种中筛选出对总丰度贡献显著的物种(表3)。其中,萼花臂尾轮虫、广布中剑水蚤、角突臂尾轮虫在禁渔前、中、后3个时期对浮游动物总丰度的贡献均显著;跨立小剑水蚤在4、5、6月对浮游动物总贡献率分别为20%、14%、16%;中华窄腹剑水蚤在禁渔后(6月)对浮游动物总丰度贡献显著,贡献率为14%。
2.3 现存量
2年调查禁渔前、中、后浮游动物丰度分布如图 2a所示。调查期间,封开浮游动物丰度变化总体表现为 5月>4月>3月>6月,丰度波动范围为3217~3628 ind∙L-1,均值为 3456 ind∙L-1;肇庆浮游动物丰度变化趋势与封开相似,丰度波动范围为3128~3757 ind∙L-1,均值为 3530 ind∙L-1;磨刀门浮游动物丰度从 3—6月总体呈下降趋势,丰度波动范围为 1391~1637 ind∙L-1,均值为 1501 ind∙L-1。方差分析结果表明,封开、肇庆、磨刀门浮游动物丰度在不同监测时间差异均不显著。
2年调查禁渔前、中、后浮游动物生物量分布如图2b所示。调查期间,封开浮游动物生物量变化总体表现为5月>4月>3月>6月,生物量波动范围为 0.931~1.162 mg∙L-1,均值为 1.104 mg∙L-1;肇庆浮游动物生物量变化趋势与封开相似,生物量波动范围为 0.903~1.062 mg∙L-1,均值为 0.967 mg∙L-1;磨刀门浮游动物生物量从3月至6月总体呈现上升趋势,生物量波动范围为0.530~0.559 mg∙L-1,均值为 0.541 mg∙L-1。方差分析结果表明,封开和肇庆浮游动物的生物量在不同监测时间差异显著(F>F0.05,P<0.05);磨刀门浮游动物生物量在不同监测时间差异不显著。

表1 浮游动物种类组成Table 1 The species composition of zooplankton

表2 浮游动物主要优势种(优势度)及分布Table 2 Dominance and distribution of the major dominant species
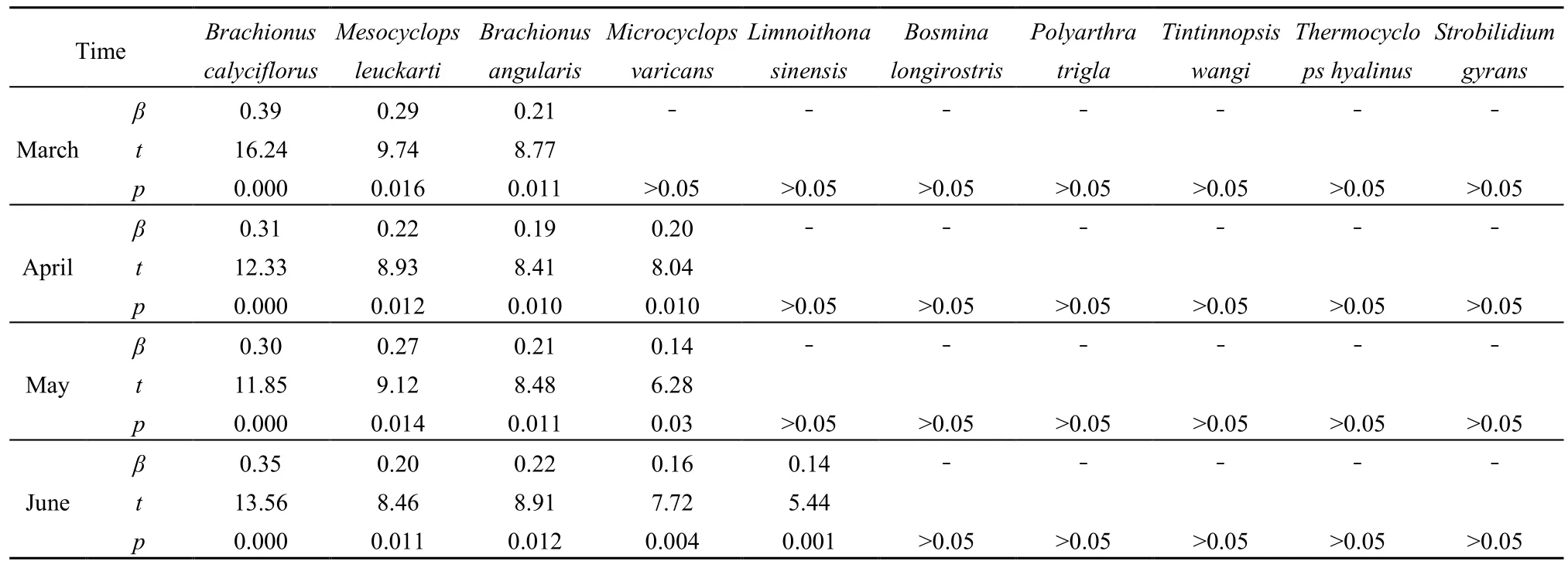
表3 浮游动物优势种对总丰度的贡献Table 3 Contribution of the dominant zooplankton to the total abundance
2.4 物种多样性
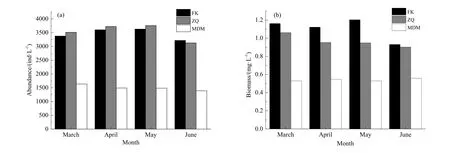
图2 浮游动物现存量分布Fig. 2 The distribution of zooplankton standing crop(a)平均丰度 the average bundance;(b)平均生物量 the average biomass
2年调查禁渔前、中、后浮游动物的平均生物多样性指数(H)和均匀度指数(J)分布如图3所示。总体上,浮游动物生物多样性指数和均匀度指数从 3—6月均呈上升趋势,生物多样性指数和均匀度指数均值分别为2.85和0.91。
方差分析结果表明,封开、肇庆、磨刀门浮游动物的生物多样性指数及均匀度指数在不同监测时间差异不显著。
将浮游动物的多样性指数及均匀度指数与环境因子进行相关性分析,结果显示,浮游动物的生物多样性指数及均匀度指数与叶绿素a含量呈极显著正相关,生物多样性指数与透明度的负相关关系达到显著水平(表4)。
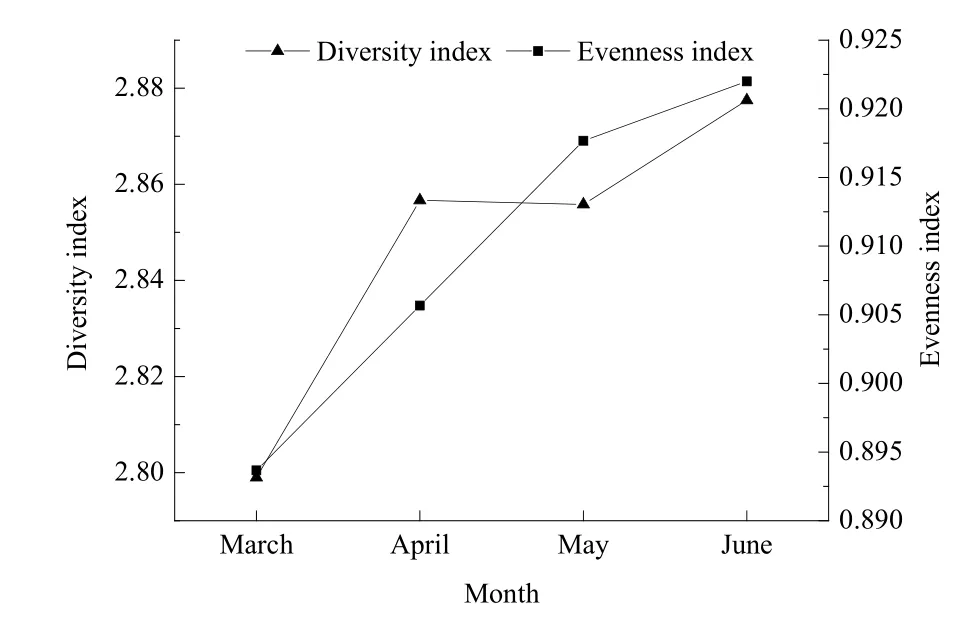
图3 浮游动物多样性指数和均匀度指数分布(2年平均)Fig. 3 The distribution of the average diversity index and the average evenness index of zooplankton

表4 浮游动物多样性与环境因子的Pearson相关性分析Table 4 The Pearson correlation analysis between zooplankton diversity and environmental factors
2.5 与非生物因子的关系
应用Canoco 4.5软件对封开、肇庆和磨刀门3个采样点浮游动物优势种的丰度数据进行降趋对应分析(DCA),在所得的各特征值部分发现 4个排序轴中梯度最大值小于 3,以此为依据选择线性模型中的间接梯度分析PCA模型对3个采样点浮游动物优势种丰度与pH、透明度、溶解氧、总氮、总磷、高锰酸盐指数、叶绿素a等因子进行分析,得到PCA二维排序结果,如图4所示。
结果显示,封开、肇庆、磨刀门3个采样点PCA二维排序的前两个排序轴对物种变量的解释量(累积贡献率)分别达到 83.6%、85%及 84.1%,可以认为前两个排序轴为主成分轴,这也意味着前2个排序轴包含有环境因子的 83.6%、85%及 84.1%的信息,所以可以用 PCA二维降维图来研究环境因子间的相互关系。3个采样点环境变量与主成分轴的相关系数见表5。
从图4a及表5可以看出,封开浮游动物优势种类丰度与总氮和pH等环境因子显著相关;从图4b及表5可以看出,肇庆浮游动物优势种类丰度与总氮呈极显著正相关,与高锰酸盐指数和叶绿素 a含量呈显著正相关,与溶解氧呈显著负相关;从图4c及表5可以看出,磨刀门浮游动物优势种类丰度与透明度呈显著正相关,与总磷呈显著负相关。
3 讨论
3.1 浮游动物群落结构变化
调查期间,禁渔期间3个采样点浮游动物丰度和生物量略大于禁渔前和禁渔后,生物多样性指数及均匀度指数从 3—6月总体呈现上升趋势,但这是否是禁渔这一单一因素对浮游动物群落结构所产生的影响?鉴于禁渔前后浮游动物群落结构还受到气候变化和环境污染等多种因素的共同影响,因此暂时很难定量评估,要准确分析浮游动物群落变化对禁渔的响应,还有待通过长期的监测进行深入研究。
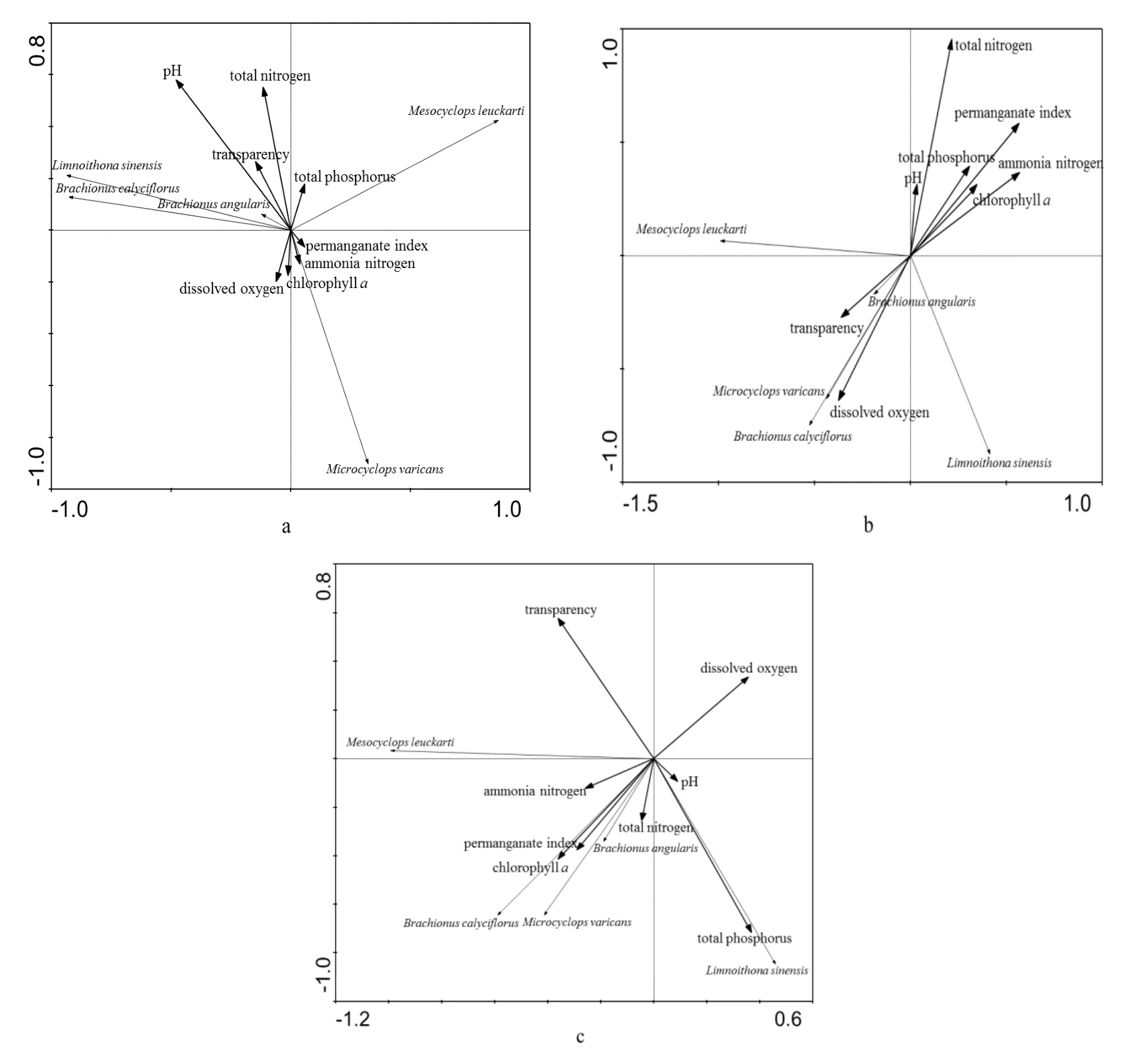
图4 物种—环境因子的PCA双序图Fig. 4 Ordination diagram of the first two axes of principle correspondence analysis of zooplankton abundance and environmental factors(a)封开 FK;(b)肇庆 ZQ;(c)磨刀门 MDM

表5 浮游动物的物种—环境因子的PCA主成分轴与环境因子之间的相关系数Table 5 Correlation coefficient for density of zooplankton axis1 and axis2, environment factors axis1and axis2, and environment factors
3.2 浮游动物群落变化与非生物因子的关系
浮游动物在水体中与多种因素有密切联系,因此浮游动物群落结构受到水生态系统中诸多因素的影响。
在本研究中,3个采样点浮游动物优势种类丰度与pH、溶解氧有明显的相关性(图4)。国内外学者研究发现,水体中的溶解氧含量和pH值的大小会影响浮游动物的分布和生长繁殖。浮游动物的生长和繁殖在溶解氧较低的条件下会受到抑制(Sedlacek et al.,2005)。各种浮游动物的最适酸碱度范围不同,故其在水体中的分布和生长代谢情况也有较大差异。此外,在溶解氧充足和pH值适宜的水体,浮游动物通过无性生殖产生非需精的夏卵,发育成非混交雌体,当水体溶氧含量过低或pH值不利时,则通过有性生殖产生冬卵以应对不良环境条件(赵文,2005)。本研究发现封开、肇庆和磨刀门3个采样点的浮游动物优势种丰度均与总氮和总磷含量密切相关。有专家研究发现,营养盐的种类及其含量是浮游动物分布极为重要的限制因子。在贫营养水体中,营养盐是浮游动物群落结构变化的主要影响因子,而在富营养化程度较高的情况下,营养盐对浮游动物群落结构变化的影响较小,成为次要因子(Reynolds,2006)。总磷含量是判定富营养化程度的主要指标之一,富营养化程度会随着磷含量的升高而加剧,从而影响浮游动物结构组成(Wen et al.,2011)。此外,水体富营养化,水质恶劣导致浮游动物的物种数和密度减少,生物多样性降低(郑金秀等,2014;成芳,2010),富营养化指示种如螺形龟甲轮虫Keratella cochlearis、臂尾轮虫 Brachionus、长三肢轮虫 Filinia longisela等大量出现(郭凯等,2010),改变浮游动物的群落结构。本研究中发现浮游动物群落与叶绿素a含量呈显著相关关系,是该水域浮游植物对浮游动物种群饵料调控的间接反映。
有研究表明,浮游动物群落结构及分布与电导率密切相关(王博等,2013;陈磊等,2016),电导率通过影响浮游植物间接作用于浮游动物(Christine et al.,2006),从而影响浮游动物的群落多样性。本研究暂时缺乏这方面的研究,将在下一步研究计划中补充分析。
3.3 生物因素对浮游动物群落结构的影响
由于生态系统空间具有差异性,浮游动物的群落结构和功能除了受到非生物因子的影响,鱼类的捕食也会对浮游动物的种类组成、多样性及分布产生重要的影响(Bröenmark et al.,2005;吴明姝,2015)。根据体积效率假说,在同等耗能的情况下,鱼类优先选择个体大且逃避能力弱的动物为食(Hall et al.,1976)。本研究中,在3、4月调查中某些体型稍大的甲壳类浮游动物在5、6月调查中优势度有所降低,而一些小型甲壳类浮游动物在禁渔期结束后优势度略有升高。这一结果是不是春季禁渔期间鱼类生殖群体及幼鱼得到有效保护的体现?笔者将在后续调查中通过在珠江河网不同生境布设采样点展开深入研究,通过数据来证明这一结论。
4 结论
(1)从调查结果来看,2年调查禁渔前、中、后的浮游动物种类数无显著性差异。3个采样点浮游动物丰度和生物量虽然在禁渔期间略大于禁渔前和禁渔后,但差异却不显著。3个采样点浮游动物的生物多样性指数及均匀度指数在不同监测时间差异不显著,但3—6月总体呈现上升趋势。
(2)封开浮游动物优势种类丰度与总氮和 pH等环境因子显著相关;肇庆浮游动物优势种类丰度与总氮呈极显著正相关,与高锰酸盐指数和叶绿素a含量呈显著正相关,与溶解氧呈显著负相关;磨刀门浮游动物优势种类丰度与透明度呈显著正相关,与总磷呈显著负相关。
BRÖENMARK C, HANSSON L A. 2005. The Biology of Lakes and Ponds[M]. Oxford, New York: Oxford University Press.
CHRISTINE L W, PAN Y D. 2006. Diatom assemblages and their associations with environmental variables in Oregon Coast Range streams USA [J]. Hydrobiologia, 561(1): 207-219.
HALL J G, THRELKELD S T, BURNS C W. 1976. The size-efficiency hypothesis and the size structure of zooplankton communities [J].Annual Review of Ecology and Systematics, 7: 177-208.
PIELOU E C. 1966. Species-diversity and pattern-diversity in the study of ecological succession [J]. Journal of Theoretical Biologyl, 10(2):370-383.
REYNOLDS C S. 2006. The ecology of freshwater phytoplankton [M].Cambridge: Cambridge University Press.
SHANNON C E, WEAVER W. 1963. The mathematical theory of communication [M]. Urbana: University of Illinois Press.
SEDLACEK C, MARCUS N H. 2005. Egg production of the copepod Acartia tonsa: The influence of hypoxia and food concentration[J].Journal of experimental marine biology and ecology, 318(2): 183-190.
WEN X L, XI Y L, QIAN F P, et al. 2011. Comparative analysis of rotifer community structure in five subtropical shallow lakes in East China:role of physical and chemical conditions. Hydrobiologia, 661(1):303-316.
陈磊, 高东泉, 舒凤月, 等. 2016. 南四湖浮游动物群落结构特征及其与环境因子的关系[J]. 动物学杂志, 51(1): 113-120.
陈希孺, 王松桂. 1987. 近代回归分析[M]. 合肥: 安徽教育出版社.
成芳. 2010. 太湖水体富营养化与水生生物群落结构的研究[D]. 苏州:苏州大学.
高原, 赖子尼, 王超, 等. 2008. 珠江口浮游动物分布特征研究[J]. 中国水产科学, 15(2): 261-269.
郭凯, 赵文, 殷守仁, 等. 2010. 北京官厅水库轮虫群落结构与水体富营养化状况[J]. 湖泊科学, 22(2): 256-264.
韩耀全, 何安尤, 施军, 等. 2015. 珠江水域(广西段)三年禁渔期效果评估[J]. 水产科技情报, 42(3): 135-139.
纪焕红, 叶属峰. 2006. 长江口浮游动物生态分布特征及其与环境的关系[J]. 海洋科学, 30(6): 23-30.
李跃飞, 李新辉, 杨计平, 等. 2014. 珠江禁渔对广东鲂资源补充群体的影响分析[J]. 水产学报, 38(4): 502-508.
刘汉武, 靳祯, 张凤琴. 2012. 休渔的建模与分析[J]. 河南师范大学学报: 自然科学版, 40(5): 8-12.
刘凯, 张敏莹, 徐东坡, 等. 2006. 长江春季禁渔对崇明北滩渔业群落的影响[J]. 中国水产科学, 13(5): 834-840.
刘凯. 2007. 长江下游渔业群落多样性现状及春禁效果初步评价[D]. 南京: 南京农业大学.
卢敬让, 李德尚. 1995. 浮游生物生物量指数作为水库营养状态和鱼产力评估指标的作用[J]. 生态学报, 15(2): 223-228.
施炜纲, 刘凯, 张敏莹, 等. 2005. 春季禁渔期间长江下游鱼虾蟹类物种多样性变动(2001—2004年)[J]. 湖泊科学, 17(2): 169-175.
谭细畅, 李新辉, 陶江平, 等. 2007. 西江肇庆江段鱼类早期资源时空分布特征研究[J]. 淡水渔业, 37(4): 37-40.
王博, 廖剑宇, 刘全儒, 等. 2013. 东江干流浮游动物群落结构特征及与水质的关系[J]. 北京师范大学学报(自然科学版), 49(6): 608-612.
吴明姝. 2015. 安徽省太平湖水库浮游动物群落结构及水质评价[D]. 上海: 上海师范大学.
徐东坡, 张敏莹, 刘凯, 等. 2006. 长江安庆江段春禁前后渔业生物多样性变化[J]. 安徽农业大学学报, 33(1): 76-80.
徐兆礼, 沈新强, 马胜伟. 2005. 春、夏季长江口邻近水域浮游动物优势种的生态特征[J]. 海洋科学, 29(12): 13-19.
徐兆礼, 王云龙, 陈亚瞿, 等. 1995. 长江口最大浑浊带区浮游动物的生态研究[J].中国水产科学, 2(1): 39-48.
严利平, 凌建忠, 李建生, 等. 2006. 应用Ricker动态综合模型模拟解析东海区伏季休渔效果[J]. 中国水产科学, 13(1): 85-91.
严利平, 刘尊雷, 李圣法, 等. 2010. 东海区拖网新伏季休渔渔业生态和资源增殖效果的分析[J]. 海洋渔业, 32(2): 186-191.
杨婉玲, 赖子尼, 曾艳艺, 等. 2017. 珠江中下游表层水体 CODMn时空分布特征及水环境评价[J]. 生态环境学报, 26(4): 643-648.
杨婉玲, 赖子尼, 庞世勋, 等. 2011. 西江广东鲂产卵场水质营养盐现状与评价[J]. 水生态学杂志, 32(5): 55-58.
张敏莹, 刘凯, 徐东坡, 等. 2006. 春季禁渔对常熟江段渔业群落结构及物种多样性影响的初步研究[J]. 长江流域资源与环境, 15(4):442-446.
章宗涉, 黄祥飞. 1995. 淡水浮游生物研究方法[M]. 北京: 科学出版社.
赵文. 2005. 水生生物学[M]. 北京: 中国农业出版社.
郑金秀, 池仕运, 李聃, 等. 2015. 富营养化对浅水湖泊轮虫种群结构影响研究[J]. 生态环境学报, 22(2): 302-310.
Abstract: The present study aimed to investigate the changes of zooplankton community before and after the spring closed season in the Pearl River. Four ecological investigations annually were carried out on zooplankton and water samples at Fengkai, Zhaoqing and Modaomen of Pearl River before (March), during (April and May) and after (June) closed fishing in 2014 and 2015. The community structure, including the distributions of species composition, dominant species, standing crop and biodiversity, was investigated. The correlation between the community structure of zooplankton and the environmental factors was discussed. Records showed that there were 88 species of zooplankton and the dominant species were Brachionus calyciflorus, Mesocyclops leuckarti, Brachionus angularis, Brachionus angularis and Limnoithona sinensis. Zooplankton had an average abundance of three sampling points ranging from 2579 to 2957 ind∙L-1, which was higher in May than April and that of March was followed and that of June was lowest. The average abundance of zooplankton of three sampling points was 0.871 mg∙L-1, which was higher in March than May and that of April was followed and that of June was lowest. In the whole investigation period, the Shannon-Wiener diversity indexes and evenness indexes increased from March to June, and the average values of the Shannon-Wiener diversity indexes and evenness indexes were 2.85, 0.91, respectively. The results of principal component analysis (PCA) showed that the dominant species abundance in Fengkai was closely correlated with the total nitrogen content and pH value; the abundance of dominant zooplankton species inluding Brachionus calyciflorus, Brachionus angularis and Limnoithona sinensis in Zhaoqing showed very significantly positive correlation with the total nitrogen content and was significantly positively correlated with the total phosphorus content, the ammonia nitrogen content, permanganate index and chlorophyll a content, while significantly negatively correlated with water transparency and dissolved oxygen content; the dominant species abundance in Modaomen was closely correlated with environment factors, such as water transparency, dissolved oxygen content, the total phosphorus content, and chlorophyll a content. We concluded that zooplankton community changed obviously before and after the spring closed season in the Pearl River artery, which was influenced by many factors such as nutrients.
Key words: the Pearl River; spring closed season; zooplankton; species composition
Changes of Zooplankton Community before and after the Spring Closed Season in the Pearl River in 2014—2015
GAO Yuan, LAI Zini*, YANG Wanling, ZENG Yanyi
Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510380, China
10.16258/j.cnki.1674-5906.2017.09.015
S931.1; X174
A
1674-5906(2017)09-1562-08
高原, 赖子尼, 杨婉玲, 曾艳艺. 2017. 2014—2015年珠江春季禁渔前后浮游动物群落变化[J]. 生态环境学报, 26(9):1562-1569.
GAO Yuan, LAI Zini, YANG Wanling, ZENG Yanyi. 2017. Changes of zooplankton community before and after the spring closed season in the Pearl River in 2014—2015 [J]. Ecology and Environmental Sciences, 26(9): 1562-1569.
中国水产科学研究院重大项目预研“珠江口水生生物食物网结构与功能研究”(2015A01YY02);农业部财政项目“珠江禁渔对浮游动物生态的影响监测及评价”
高原(1981年生),女,助理研究员,研究方向为渔业环境保护及修复。E-mail: gaoyuan@prfri.ac.cn*通信作者。赖子尼,E-mail: znlai01@prfri.ac.cn
2017-07-12
