向日葵主要农艺与品质性状配合力及杂种优势分析
2017-09-25贾秀苹卯旭辉梁根生赵晓文甘肃省农业科学院作物研究所兰州730070甘肃省农业技术推广总站兰州73000
贾秀苹,卯旭辉,岳 云,梁根生,赵晓文(.甘肃省农业科学院 作物研究所,兰州 730070; .甘肃省农业技术推广总站,兰州 73000)
向日葵主要农艺与品质性状配合力及杂种优势分析
贾秀苹1,卯旭辉1,岳 云2,梁根生1,赵晓文2
(1.甘肃省农业科学院 作物研究所,兰州 730070; 2.甘肃省农业技术推广总站,兰州 730020)
选用6份不育系和6份恢复系材料为亲本,按照NCⅡ不完全双列杂交设计配制36份向日葵杂交组合,对其组合的7个主要农艺性状和2个品质性状的亲本配合力及杂种优势进行分析。结果表明:36份杂交组合的主要农艺和品质性状间存在显著的遗传差异。其中株高、茎粗、叶片数、盘径、百粒质量、粗蛋白各性状同时受基因加性效应和非加性效应的共同作用,但基因非加性效应大于加性效应;单盘实粒数、单株产量、粗脂肪等性状受基因加性效应主导;广义遗传力大小顺序为:百粒质量>单株产量>单盘实粒数。亲本一般配合力(GCA)和特殊配合力(SCA)与向日葵杂交组合竞争优势呈极显著正相关。强优势组合特点是双亲或亲本之一具有较高的GCA效应或SCA效应。12份亲本材料中,恢复系X3639R、318R、LD5009R在单株产量、农艺性状、单盘实粒数均表现为较大的正向GCA效应值;不育系S10-390-3A、S10-336-2A、S10-390-1A在百粒质量、单株产量、粗脂肪、粗蛋白方面均表现为较大的正向GCA效应值,故均为较好的亲本材料。此外,除茎粗外,恢复系的GCA效应对杂种优势的贡献明显高于不育系。向日葵杂交组合主要农艺和品质性状存在广泛的杂种优势。叶片、单盘实粒数、单株产量平均超亲优势分别为66.13%、73.58%、61.17%。粗蛋白平均超亲优势值为68.96%,各性状正向超亲优势组合数超过83%。
向日葵;一般配合力;特殊配合力;杂种优势
向日葵是世界第三大油料作物,中国第四大油料作物。自20世纪60年代首次在向日葵野生种和栽培种的一个杂交后代中选育出细胞质雄性不育系以来,向日葵育种工作在世界范围内得到快速发展[1]。作物杂种优势的研究和利用是现代育种学成就突出的重要领域,也是发掘农作物生产潜力最有效的育种手段[2]。而配合力是决定作物杂种优势的主要因素,也是选配亲本的重要依据[3-6]。只有选育出高配合力的亲本,才有可能组配出强优势的杂交组合[7]。国内外学者在主要农作物农艺、品质、抗性等相关性状的配合力与杂种优势方面已有大量的研究[8-9]。近几年随着向日葵产业的迅速发展,其杂种优势利用研究突飞猛进,加之中国具有丰富的向日葵遗传资源,特别是食用向日葵,数百个地方品种散布在全国各地,这些材料之间具有形态上的差异,但材料间的遗传背景、亲本配合力、杂种优势及亲缘关系等尚不清楚[10]。本试验选取6个自育胞质雄性不育系和6个分枝型恢复系按照NCⅡ不完全双列杂交设计配置36份杂交组合,对亲本的一般配合力、特殊配合力及遗传效应与杂种优势进行深入分析,旨在为向日葵杂交育种提供理论依据。
1 材料与方法
1.1 材 料
选用来源于国内外的6份分枝型恢复系X3639R、FK5036R、318R、9638R、LD5009R、3939R作父本(P1),甘肃省农业科学院作物研究所自育的6份胞质互作不育系S07-153-4-1A、S10-336-2A、S10-390-1A、S10-390-2A、S0920A、S10-390-3A作母本(P2),LD5009为对照品种,各亲本来源及系谱见表1。
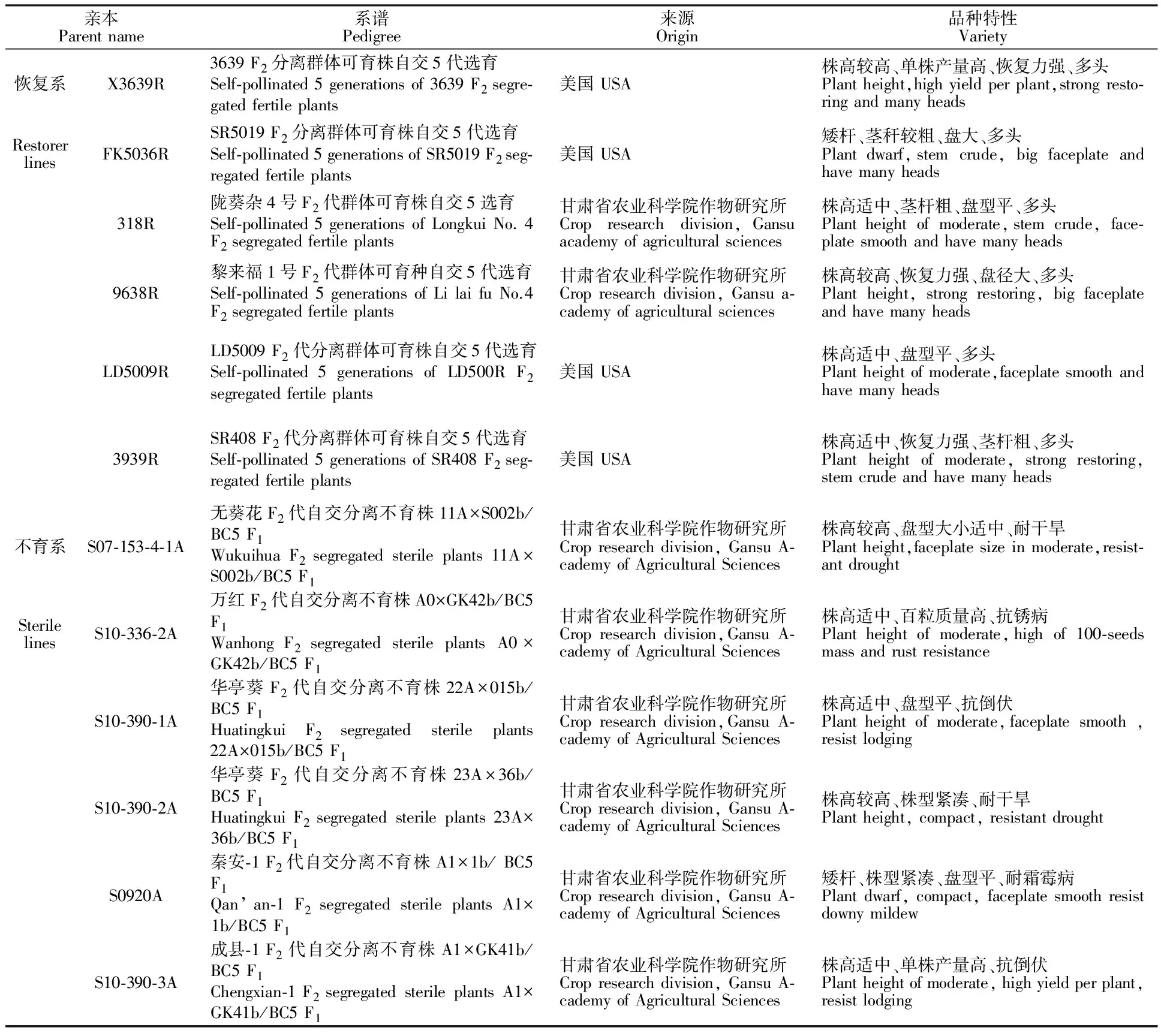
表1 供试材料名称及来源Table 1 Material and origin
1.2 试验设计
试验设在甘肃省景泰县向日葵试验基地,于2014-04-18播种材料,按照6×6不完全双列杂交(NCⅡ)设计,随机区组排列,3次重复,每小区种植4行,每行13株,株距40 cm,行距50 cm,小区面积5.2 m×2 m=10.4 m2。
1.3 测定指标
成熟期取中间2行具有代表性的9株,考察株高(基部至花盘背部)、茎粗(株高一半处茎秆直径)、叶片数(茎基部子叶以上至花盘处全部叶片)、花盘直径(花盘正面直径)、单盘实粒数(粒)、单株产量(g)、百粒质量 (g)7个农艺性状指标。并依据索氏提取法GB/T 14772-2008[11]测定粗脂肪和粗蛋白质量分数。
1.4 数据统计分析
数据处理与遗传交配设计统计分析分别采用Microsoft Excel 2003 及 DPS 7.05软件进行,在性状方差和配合力方法分析的基础上,估算不育系、恢复系的一般配合力(General combining ability,GCA)、各杂交组合的特殊配合力(Specific combining ability,SCA)及其遗传参数。
杂种优势计算:超亲优势(PH)=(F1-P)/P×100%,F1为杂交F1代值,P为优势亲本父本值;竞争优势(CH)=(F1-CK)/CK×100%,CK为对照(LD5009)值。
2 结果与分析
2.1 供试材料农艺性状方差与配合力分析
通过对6份父本、6份母本所组配的36份杂交组合的株高、茎粗、叶片数、盘径等7个主要农艺性状和粗脂肪、粗蛋白等品质性状进行方差分析(表2),结果表明,农艺组除单盘实粒数外,其余性状组合间差异达显著或极显著水平,说明供试各杂交组合间存在真实的遗传差异。以各组合平均值为单位,进一步对恢复系(GCAR)和不育系(GCAS)的一般配合力和特殊配合力进行方差分析,结果表明恢复系除农艺组的茎粗与百粒质量外,其他各性状均达极显著水平,说明受基因加性效应控制,早代加强对这些性状的选择是必要的。不育系除百粒质量外其他各性状一般配合力方差均为不显著。特殊配合力除叶片、单株产量和粗蛋白外,其他各性状达极显著水平,说明一般配合力低的亲本其特殊配合力不一定低,组配出强优势杂交组合有一定的现实性。
农艺组恢复系的株高、盘径、单盘实粒数及品质组粗脂肪的一般配合力和组合的特殊配合力方差均达显著水平,表明受基因加性效应和非加性效应的共同作用,所以在亲本恢复系一般配合力选择的基础上,应注意组合特殊配合力的选择。同时发现不育系农艺组除百粒质量外的其他农艺性状与品质组的粗脂肪和粗蛋白的一般配合力均为不显著,表明在两组亲本中恢复系对主要农艺和品质性状的改良上具有决定性的作用,因此,在向日葵杂交组合选育上要特别注重对恢复系性状的选择。
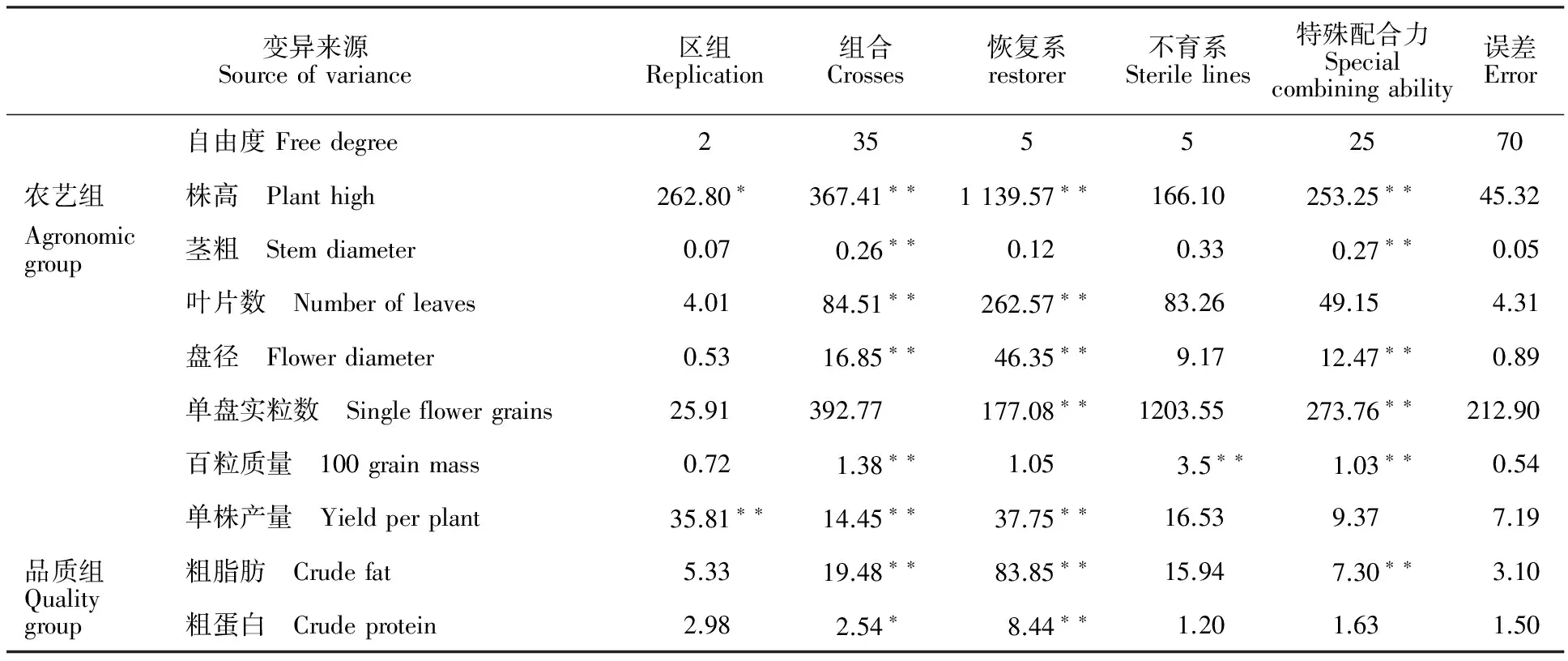
表2 各性状的方差和配合力方差分析Table 2 Variance analysis of crosses with different tested characters
注:*表示在 0.05水平差异显著,**表示在0.01水平差异显著。表7同。
Note:* and ** means significant difference level at 0.05 and 0.01, respectively.The same as table 7.
2.2 亲本的一般配合力(GCA)效应分析
一般配合力效应大小是评价亲本在杂交育种中利用价值的重要指标[12]。从表3可以看出,同一亲本不同性状间的一般配合力差异较大,表明同一亲本材料在不同形状间的一般配合力效应值不同。同时发现同一性状在不同亲本间的一般配合力值差异较大,进一步表明杂交向日葵这些性状受核基因和胞质基因互作的影响较大。
6份恢复系中,X3639R在单盘实粒数、百粒质量、单株产量上均为正向GCA效应值,其中单株产量的GCA值居供试恢复系之首,所以X3639R是提高杂交组合产量较好的亲本材料。318R在株高、茎粗、叶片、盘径上均具有较大的正向GCA效应值,在粗脂肪质量分数上较其他恢复系表现最优的正向配合力效应值,所以是选育高粗脂肪质量分数为目标的优良亲本材料。LD5009R在株高、茎粗、叶片数、单盘实粒数上均表现较好的正向GCA效应值,同时在粗蛋白质量分数上表现为最优的配合力效应,所以可认为是以粗蛋白质量分数为选育目标的优良亲本材料。FK5036R在株高上表现最低的负向GCA效应值,所以可视为以矮秆为目标的较好亲本材料。亲本9638R和3939R无论在农艺还是品质各性状上GCA均表现普通。
6份不育系中,S10-390-3A在株高、茎粗、单盘实粒数、百粒质量和单株产量上均表现较好正向GCA效应值,其中百粒质量效应值在所有供试不育系材料中为最高,所以为较好的高百粒质量不育系亲本材料。S10-336-2A在单株产量、粗脂肪上在所供试不育系材料中均表现为最好的正向GCA效应值,株高表现为负向GCA效应值,所以是培育矮秆、高产、高脂肪质量分数的理想不育系材料。S10-390-1A在叶片数、盘径、粗蛋白方面表现为正向GCA效应值,株高表现为最大负向GCA效应值,所以为较好的高蛋白、矮秆的理想不育系材料。S07-153-4-1A、S10-390-2A、S0920A无论是农艺性还是品质性状均表现一般。

表3 亲本一般配合力(GCA)效应分析Table 3 Estimates of general combining ability effects of parents for measured characters
2.3 杂交组合特殊配合力(SCA)效应分析
表4显示同一组合不同性状间或同一性状的不同组合间特殊配合力存在较大差异,这与一般配合力的分析结果相似。特殊配合力方差分析结果表明,农艺组的株高、茎粗、盘径、单盘实粒数、百粒质量和品质组的粗脂肪质量分数的特殊配合力方差达极显著水平。从特殊配合力效应值看,株高SCA变幅为-8.80~8.74,正向效应组合数为18个;茎粗SCA变幅为-17.53~15.34,正向效应组合数为16个;盘径SCA变幅为-17.81~18.28,正向效应组合数为20个;单盘实粒数SCA变幅为-2.90~1.79,正向效应组合数为20个;百粒质量SCA变幅为-4.95~7.89,正向效应组合数为15个;粗脂肪SCA变幅为-10.81~12.26,正向效应组合数为16个。其他性状组合间特殊配合力方差并不显著。
对组合SCA效应值与亲本GCA值进行比较,发现亲本各性状GCA效应与各组合SCA效应之间是相互独立的,不存在任何规律或一定的比例关系。如茎粗的SCA值较高但双亲的GCA效应值均较低;百粒质量的SCA效应值高,亲本GCAS效应值高,但亲本GCAR效应值较低;单盘实粒数SCA效应值高,亲本GCAS效应值低,但亲本GCAR效应值较高,其他各形状存在类似情况。
2.4 遗传效应和遗传参数估算
基因型方差和群体配合力方差分析(表5)表明,群体总一般配合力比较低。其中株高、茎粗、叶片数、盘径、百粒质量亲本一般配合力方差占总配合力方差为3.98%~47.92%。说明这些性状同时受基因加性效应和非加性效应的共同作用,但基因非加性效应大于加性效应。单盘实粒数、单株产量、粗蛋白、粗脂肪的亲本一般配合力方差占主要优势,占总方差71.80%~89.67%,说明在这些性状的遗传中亲本加性基因效应起主导作用。 从农艺组各性状的遗传力看,株高、茎粗、叶片数、盘径的广义遗传力较高分别为72.34%、59.43%、86.93%、86.60%,表明这些性状受加性遗传和非加性遗传共同作用,但非加性遗传占主导作用;其余性状的狭义遗传力均低于40%,大小顺序为:百粒质量>单株产量>盘实粒数。品质组中粗脂肪的狭义遗传力较高,为66.39%,说明该性状受加性遗产和非加性遗传共同作用,但非加性遗传占主导作用;粗蛋白狭义遗传力为21.92%,说明该性状受加性遗传和非加性遗传共同作用,但加性遗传较为突出。

表4 组合特殊配合力(SCA)效应值Table 4 Estimates of specific combining ability effects for measured characters

表5 各性状的遗传参数估计Table 5 Estimates of genetic parameters for measured characters
2.5 主要农艺与品质性状的杂种优势
36份杂交组合F1代主要农艺及品质性状的杂种优势,在不同组合同一形状和不同性状间存在较大差异。表6显示,向日葵三系杂交组合间存在广泛的超亲优势及竞争优势。超亲优势分析,各组合农艺性状间超亲优势表现突出,其中叶片、单盘实粒数、单株产量超亲优势变异幅度较大分别为2.60%~171.50%、40.90%~147.40%、11.20%~290.50%,平均超亲优势分别为66.13%、73.58%、61.17%。品质性状间粗脂肪超亲优势较弱,但粗蛋白超亲优势为3.0%~35.70%,其平均值为68.96%。各性状正向超亲优势组合数超过83.00%,说明向日葵是杂种优势利用研究的典范作物。
竞争优势分析,农艺组除株高外,其他各性状均存在较强的竞争优势,其中叶片竞争优势最强高98.40%;茎粗、盘径、百粒质量的最大竞争优势分别达38.30%、47.65%、25.60%;最小正向优势竞争数在55.60%以上。叶片数、百粒质量、单盘实粒数正向竞争优势均为100%。品质组粗脂肪竞争优势最强为30.60%,粗蛋白为11.10%,可见向日葵杂种优势利用研究对改善品质存在较大潜力。

表6 36个组合的超亲优势和竞争优势Table 6 Heterosis over male parent (MP) and control cultivar (CH) for measured characters %
2.6 杂种优势与GCA和SCA的关系
表7显示,茎粗的GCA与超亲优势未达显著相关关系;单盘实粒数GCA与超亲优势表现负向,但未达到显著水平;其他各性状与超亲优势呈极显著正相关关系。茎粗、叶片、粗脂肪、粗蛋白SCA值与超亲优势呈显著或极显著正相关,其他各性状未达到显著相关水平。所有性状的GCA、SCA值与对照竞争优势呈极显著正相关。可见,向日葵杂种优势表现与亲本一般配合力和组合特殊配合力具有一定的相关关系。
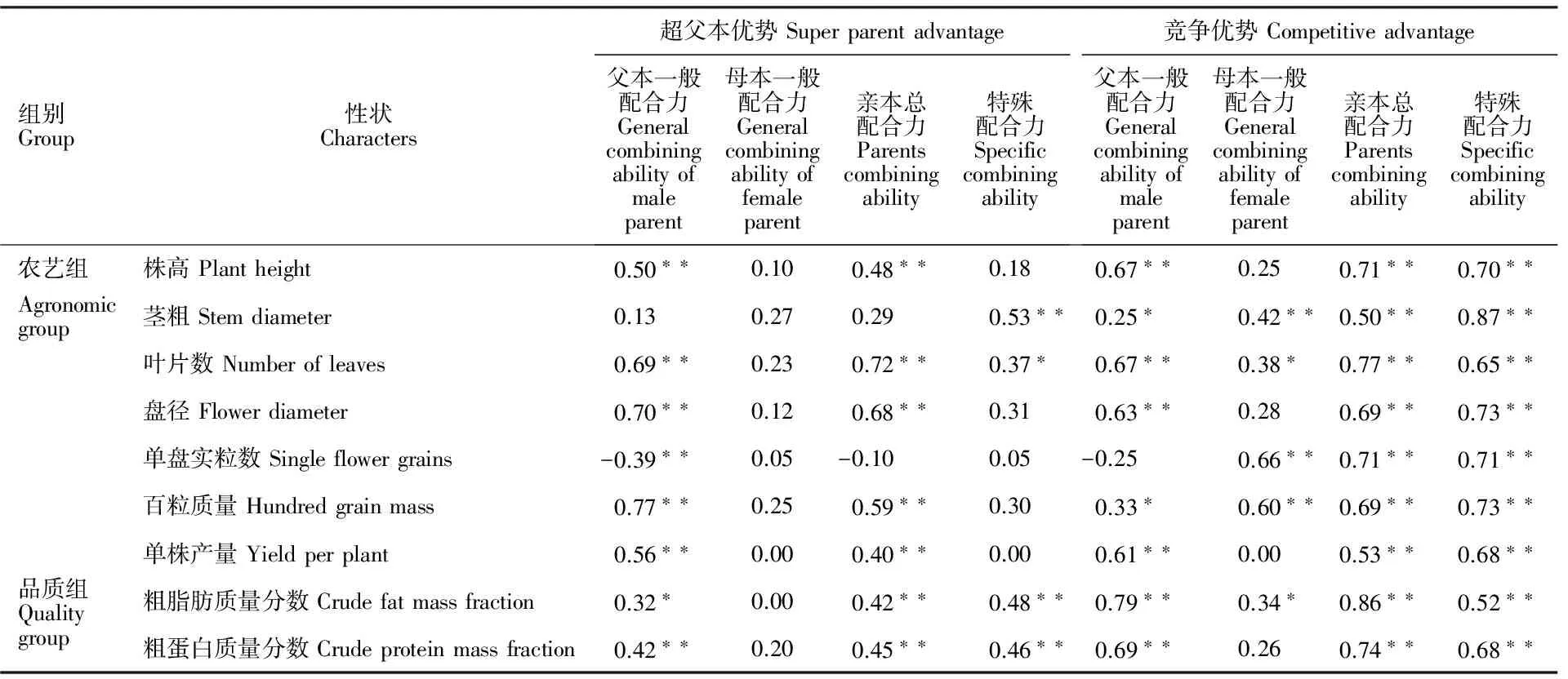
表7 各性状优势值与GCA、SCA的简单相关系数Table 7 Correlation coefficient among GCA,SCA and heterosis
3 讨论与结论
3.1 向日葵主要农艺及品质性状配合力
育种的目的是通过不同亲本间的杂交,选育出在综合性状上能优于其亲本或优良性状能够互补的杂交组合[13]。杂优利用中,配合力是作物杂种优势利用亲本选配的重要遗传基础,育种实践表明一般只有配合力好的亲本才有可能选育出农艺及品质优良的杂交组合。亲本的一般配合力反映亲本交配效应平均的强弱,即反映亲本性状在后代中的平均表现,主要用于度量基因遗传的加性效应[14],同一亲本不同性状的配合力效应差异明显,不同亲本同一性状配合力效应也不同,说明基因加性效应及非加性效应间有着本质上的差导。Shahnaz等[8]研究发现向日葵的性状主要受基因加性效应为主,同时不能排除基因的非加性效应。本试验对向日葵亲本主要农艺及品质性状研究发现,不育系除百粒质量外其他性状均达不显著水平。
恢复系除了茎粗与百粒质量外,其他各性状均达极显著水平,表明受基因加性效应控制。株高、盘经、单盘实粒数及品质组粗脂肪的一般配合力和组合特殊配合力方差均达极显著水平,表明受基因加性效应和非加性效应的共同作用。这与杨新元等[15]、戴正元等[12]的研究结果既有相似之处又有差异,这可能与不同的研究材料有关。同时进一步对组合SCA效应值与亲本GCA效应值进行比较,发现亲本各性状GCA效应与各组合SCA效应之间是相互独立的,不存在任何规律或一定的比例关系。这一方面为选育一般配合力和特殊配合力结合得很好的强优势组合提供可能,另一方面又要求在杂交组合选配中,不仅要注意亲本一般配合力的选择,而且还必须重视广泛的组合评鉴工作,这与廖伏明等[16]的研究结果相一致,所以在实际育种工作中应考虑所用材料的遗传特点,不能一概而论。
3.2一般配合力和特殊配合力效应与杂种优势关系
特殊配合力指一个亲本在与另一亲本所产生杂交组合的性状表现中偏离两亲本平均效应的特殊效应,其主要是由基因显性、上位性等效应产生;其遗传机制复杂,且不能固定遗传,但这也是产生杂种优势的内在遗传机制[5,17],杂种优势利用的关键是选育出强优势的杂交组合。研究表明[18-19]亲本一般配合力和组合特殊配合力对作物杂种优势形成具有重要作用。但侧重性状有所不同,管荣展等[20]研究认为双亲一般配合力之和越大,产量杂种优势越强,但相关程度并不高。Grant等[21]研究配合力与杂种优势间的关系,认为特殊配合力对杂种优势更重要。本研究将7个农艺和2个品质性状的优势值与GCA和SCA值进行相关分析,结果表明GCA和SCA与向日葵杂交组合F1代竞争优势呈极显著的正相关,且强优势组合的特点是双亲或亲本之一具有较高的GCA效应,或具有较高的SCA效应。可见,向日葵亲本一般配合力和组合特殊配合力与杂种优势利用具有一定的相关关系。综合分析发现,6份恢复系中,X3639R在农艺组的单盘实粒数、百粒质量、单株产量上均有较大的正向GCA效应值,其中单株产量的GCA值居供试恢复系之首,所以是一个高产较好的亲本材料;318R在品质组粗脂肪质量分数上较其他恢复系表现最优的正向配合力效应值,所以是选育高粗脂肪质量分数为目的优良亲本材料;LD5009R在粗蛋白质量分数上表现为最优的配合力效应,可视为高粗蛋白质量分数为目标的优良亲本材料。不育系S10-390-3A的百粒质量效应值在所有供试不育系材料中最高,所以为较好的高百粒质量不育系亲本材料;S10-390-1A粗蛋白方面表现为正向GCA效应值,株高表现为最大负向GCA效应值,所以为较好的高粗蛋白,矮秆的理想不育系材料。此外,通过配合力方差分析及GCA与杂种优势的相关性分析结果表明,不育系和恢复系配置F1代杂交组合中恢复系对主要农艺和品质性状的改良上具有决定性的作用,因此在向日葵杂交组合选育上要特别注重对恢复系性状的选择。
3.3 向日葵杂种优势利用研究前景
杂种优势是自然界普遍存在的一种生物学现象[22-24],已经在农业生产上得到广泛的应用。向日葵杂种优势较强,与普通品种比较,其产量和品质均有明显提升,增产幅度达15%~40%[25-26]。本研究对向日葵三系杂交组合的杂种优势分析结果表明,各组合农艺性状间超亲优势表现突出,其中叶片、单盘实粒数、单株产量超亲优势变异幅度较大分别为2.60%~171.50%、40.90%~147.40%、11.20%~290.50%,平均超亲优势分别为66.13%、73.58%、61.17%。各性状正向超亲优势组合数超过83%。茎粗、盘径、百粒质量最大竞争优势分别达38.30%、47.65%、25.60%;最小正向优势竞争数在55.60%以上。叶片数、百粒质量、单盘实粒数正向竞争优势均为100%。同时发现,竞争优势与亲本GCA和组合SCA效应值呈极显著正相关。表明向日葵是杂种优势利用研究的典范作物,因此,充分利用大量亲本资源材料进行广泛杂交组配,加强亲本一般配合力和特殊配合力的选择效应研究,对向日葵杂种优势利用将会获得突破性进展。
Reference:
[1] 安玉麟,孙瑞芬,冯万玉.我国向日葵品种改良进展及其与国外的差距[J].华北农学报,2006(S3):1-4.
AN Y L,SUN R F,FENG W Y.The progress of improvement in sunflower species in our country and the GaP between advanced level in overseas and China [J].ActaAgriculturaeBoreali-Sinica,2006(S3):1-4(in Chinese with English abstract).
[2] 张彩霞,柴守诚,郑炜君.六倍体小黑麦T型细胞质雄性不育体系杂种优势与配合力的研究[J].西北植物学报,2005,25(5):898-902.
ZHANG C X,CHAI SH CH,ZHENG W J.Study on heterosis and combining ability of male sterile system of hexaploid triticale withT.timopheevicystoplasm[J].ActaBotanicaBoreali-occidentaliaSinica,2005,25(5):898-902(in Chinese with English abstract).
[3] 张建一,杨世尧,赖群启.几个甘蓝型油菜品种的配合力分析[J].中国农业科学,1983(2):26-32.
ZHANG J Y,YANG SH Y,LAI Q Q.Analysis of the combining ability in some rapeseed(BrassicanapusL.) varieties[J].ScientiaAgriculturaSinica,1983(2):26-32(in Chinese with English abstract).
[4] 张征锋,肖本泽.利用AMMI模型分析杂交水稻配合力[J].植物遗传资源学报,2015,16(2):400-404.
ZHANG ZH F,XIAO B Z.Combining ability analysis of rice hybrids using AMMI model[J].JournalofPlantGeneticResources,2015,16(2):400-404(in Chinese with English abstract).
[5] 蒋开锋,郑家奎,赵甘霖,等.基于AMMI模型的 NCⅡ交配设计试验的配合力分析[J].作物学报,2000,26(6):959-962.
JIANG K F,ZHENG J K,ZHAO G L,etal.Analysis of combining ability based on AMMI model[J].ActaAgronomicaSinica,2000,26(6):959-962(in Chinese with English abstract).
[6] 孔繁玲.植物数量遗传学[M].北京:中国农业大学出版社,2006:401-402.
KONG F L.Genetics of Plant Quantity [M].Beijing:China Agricultural University Press,2006:401-402(in Chinese).
[7] 肖本泽,张征锋,何 亮,等.抗除草剂杂交籼稻亲本的配合力分析[J].植物遗传资源学报,2012,13(4):562-570.
XIAO B Z,ZHANG ZH F,HE L,etal.Combining ability analysis of parents of herbicide-resistant indica hybrids [J].JournalofPlantGeneticResources,2012,13(4):562-570(in Chinese with English abstract).
[8] SHAHNAZ M,MUHAMMAD J B,GUL M B,etal.Combining ability through line×tester analysis for phonological,seed yeld,and oil trait in sunflower(HelianthusannusL.) [J].Euphytica,2015,204(1):199-209.
[9] GODFREE C H,KINGSTO M.PAUL S H,Advanced cycle pedigree breeding in sunflower.Ⅱ:combining ability for oil yied and its components[J].Euphytica,2014(195):183-195.
[10] 刘 杰,莫结胜,刘公社,等.向日葵种质资源的随机扩增多态性DNA(RAPD)研究[J].植物学报,2001,43(2):151-157.
LIU J,MO J SH,LIU G SH,etal.RAPD analysis on the germplasm resources of sunflower[J].ActaBotanicaSinica,2001,43(2):151-157(in Chinese with English abstract).
[11] 国家标准GB/T 14772-2008.食品中粗脂肪的测定[S].北京:中华人民共和国质量监督检验检疫总局/中国国家标准化管理委员会,2014.
National Standard GB/T 14772-2008.Determination of Crude Fat in Foods[S].Beijing:General Administration of Inspection Quarantine and Quality Supervision of People Republic of China/Management Committee of China National Standardization,2014.
[12] 戴正元,李爱宏,刘广青,等.几个优良籼稻亲本品质性状的配合力和杂种优势分析[J].植物遗传资源学报,2006,7(2):234-238.
DAI ZH Y,LI A H,LIU G Q,etal.Combining ability and heterosis analysis for rice quality traits in some elite parents[J].ActaAgronomicaSinica,2006,7(2):234-238(in Chinese with English abstract).
[13] 魏忠芬,李德文,王 军,等.甘蓝型杂交油菜亲本含油量及主要农艺性状配合力与遗传力分析[J].西北农业学报,2014,23(6):102-108.
WEI ZH F,LI D W,WANG J,etal.Analysis on combining ability and heritability of main agronomic traits and oil contents of hybrid parents inBrassicanapusL[J].ActaAgriculturaeBoreali-occidentalisSinica,2014,23(6):102-108(in Chinese with English abstract).
[14] 孔宪辉,余 渝,刘 丽,等.陆地棉主要性状杂种优势与配合力研究[J].西北农业学报,2011,20(9):54-60.
KONG X H,YU Y,LIU L,etal.Analysis the heterosis and combining ability of main traits for upland cotton [J].ActaAgriculturaeBoreali-occidentalisSinica,2011,20(9):54-60(in Chinese with English abstract).
[15] 杨新元,黄增强.向日葵自交系主要性状的配合力分析[J].山西农业科学,1987(9):5-7.
YANG X Y,HUANG Z Q.Combining ability analysis of main characters of sunflower inbred lines[J].JournalofShanxiAgriculturalSciences,1987(9):5-7(in Chinese).
[16] 廖伏明,周坤炉,盛孝邦,等.釉型三系杂交水稻主要农艺性状配合力研究[J].作物学报,1999,25(5):622-631.
LIAO F M,ZHOU K L,SHENG X B,etal.Studies on combining ability of major agronomie characters in three-1ine indica hybrid rice[J].ActaAgronomicaSinica,1999,25(5):622-631(in Chinese with English abstract).
[17] 陈晓文,马守才,王志军,等.15个化杀杂交小麦亲本配合力和杂种优势群的初步研究[J].麦类作物学报,2011,31(4):630-636.
CHEN X W,MA SH C,WANG ZH J,etal.Study on the combining ability and heterotic grouping of 15 parents for CHA hybrid wheat [J].JournalofTriticeaeCrops,2011,31(4):630-636(in Chinese with English abstract).
[18] 杨加银,盖钧镒.黄淮地区大豆重要亲本间产量的杂种优势、配合力及其遗传基础[J].作物学报,2009,35(4):620-630.
YANG J Y,GAI J Y.Heterosis,combining ability and their genetic basis of yield among key parental materials of soybean in Huang-Huai valleys [J].ActaAgronomicaSinica,2009,35(4):620-630(in Chinese with English abstract).
[19] 史秀秀,毕晓静,马守才,等.黄淮麦区杂交小麦亲本的杂种优势和配合力分析[J].麦类作物学报,2013,33(6):1111-1118.
SHI X X,BI X J,MA SH C,etal.Combinging ability and heterosis in hybrid whert of parents from Huang-Huai wheat production area [J].JournalofTriticeaeCrops,2013,33(6):1111-1118(in Chinese with English abstract).
[20] 管荣展,盖钧镒.甘蓝型油菜杂种优势及其与亲本配合力和亲本系数间的关系[J].中国油料作物学报,1998,20(4):11-15.
GUAN R ZH,GAI J Y.A study on relationship of heterosis with combining ability and coefficient of parentage inRassicanapusL.[J].ChineseJournalofOilScieves,1998,20(4):11-15(in Chinese with English abstract).
[21] GRANT I,BEVERSDORF W D.Heterosis and combining ability estimates in spring oilseed rape(BrassicanapusL.)[J].CanadianJournalofGeneticsandCytology,1985(27):472-478.
[22] 张天真.作物育种学总论[M].北京:中国农业出版社,2005:156-157.
ZHANG T ZH.Crop Breeding[M].Beijing:China Agriculture Press,2005:156-157(in Chinese).
[23] HOECKER N,KELLER B,PIEPHO H P,etal.Manifestation of heterosisduring early maize(ZeamaysL.) root development[J].TheoreticalandAppliedGenetics,2006,112(3):421-429.
[24] MEYER S,POSPISIL H,SCHOLTEN S.Heterosis associated gene expression in maize embryos 6 days after fertilization exhibits additive,dominant and overdominant pattern[J].PlantMolecularBiology,2007,63(3):381-391.
[25] 崔会会,项 超,石英尧,等.杂种优势形成的表观遗传学研究进展[J].植物遗传资源学报,2015,16(5):933-939.
CUI H H,XIANG CH,SHI Y R,etal.The research progress of epigenetic insights into bases of heterosis [J].ActaAgronomicaSinica,2015,16(5):933-939(in Chinese with English abstract).
[26] 范丽娟.向日葵商品杂交种选育“三系”材料的利用及其杂交种选育方法[J].农业科技通讯,2014(9):227-228.
FAN L J.Utilization of “three lines” material for breeding sunflower commodity hybrid and hybrid breeding method thereof [J].AgriculturalScienceandTechnologyCommunication,2014(9):227-228(in Chinese with English abstract).
AnalysisoftheCombiningAbilityandHeterosisonMainAgronomicandQualityTraitsinSunflower
JIA Xiuping1,MAO Xuhui1,YUE Yun2,LIANG Gensheng1and ZHAO Xiaowen2
(1.Crop Research Division,Gansu Academy of Agricultural Sciences, Lanzhou 730070, China; 2.Gansu Agricultural Technology Extension Station, Lanzhou 730020, China)
The heterosis and coming ability of six agronomic characters and two quality traits were analyzed using 36 hybrid combination derived from 6 male sterile lines and 6 restorer by incomplete diallel (NCII) cross in sunflower. After analysis the combining ability and heterosis of parents in 7 agronomic and 2 quality traits, the results showed that a significant differences existed in among main agronomic and quality in 36 hybrid combinations. The plant height, stem diameter, leaf number, faceplate diameter, single disc filled grains, hundred-grain mass, and crude protein were effected by both additive and non-additive genes, but non-additive gene effect was greater than additive. The number of single disc filled grains, yield per plant, crude fat and other traits were dominantly by additive effect. The order of broad-sense heritability was hundred-grain mass>yield per plant>single disc grains. The heterosis of hybrid combina were positively correlate with GCA and SCA of its parents. Strong heterotic could be gained by higher GCA or SCA parents. In 12 parent materials, the restorers lines X3639R, 318R and LD5009R had more positive GCA in single plant yields, agronomic traits and single disc grains, but sterile lines S10-390-3A, S10-336-2A and S10-390-1A had significantly positive GCA effect in hundred-grain mass, yield per plant, crude fat, and crude protein mass fraction, they were all preferable parent materials. In addition, the GCA contribution of restorer lines to heterosis was significantly greater than that of sterile lines. The heterobeltiosis exist in agronomic characters and quality traits of sunflower. The heterobeltiosis of leaf unmber, single disc grains and yield per plant was 66.13%, 73.58% and 61.17%, respectively, and the average heterosis of crude protein was 68.96%, and the positive heterobeltiosis combination of various traits was over 83%.
Sunflower; General combining ability; Specific combining ability; Heterosis
2016-11-03
2017-01-11
Fund of National Natural Science Area(No.31360343);Fund of Condition Construction of Research and Achievement Transformantion Project of Gansu Academy of Agricultural Sciences(No.2016GAAS35);Fund of Modern Agricultural Industry Technology System Construction of Special Funds(No.CARS-16); Biological Technology Research Application and Development Project in Gansu(No.GNSW-2016-6).
JIA Xiuping, female,master student,assistant researcher.Research area:potato sunflower breeding research. E-mail: gsjxp666@163.com
S565.5
A
1004-1389(2017)09-1334-10
(责任编辑:成敏Responsibleeditor:CHENGMin)
日期:2017-09-12
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170912.1740.012.html
2016-11-03
2017-01-11
国家自然科学地区基金(31360343);甘肃省农业科学院科研条件建设及成果转化项目(2016GAAS35);现代农业产业技术体系建设专项资金(CARS-16);甘肃省生物技术研究与应用开发项目(GNSW-2016-6)。
贾秀苹,女,硕士研究生,助理研究员,从事向日葵遗传育种研究。E-mail:gsjxp666@163.com
卯旭辉,男,学士,副研究员,主要从事向日葵遗传育种研究。E-mail:wd-mxh@163.com
CorrespondingauthorMAO Xuhui, male, associate researcher. Research area: potato sunflower breeding research. E-mail: wd-mxh@163.com
