木薯钙调蛋白的原核表达及其单克隆抗体的制备
2017-09-25欧文军余厚美安飞飞罗秀芹秦于玲陈松笔中国热带农业科学院热带作物品种资源研究所农业部木薯种质资源保护与利用重点实验室海南儋州571737
欧文军,余厚美,安飞飞,罗秀芹,秦于玲,陈松笔(中国热带农业科学院 热带作物品种资源研究所/农业部木薯种质资源保护与利用重点实验室,海南儋州 571737)
木薯钙调蛋白的原核表达及其单克隆抗体的制备
欧文军,余厚美,安飞飞,罗秀芹,秦于玲,陈松笔
(中国热带农业科学院 热带作物品种资源研究所/农业部木薯种质资源保护与利用重点实验室,海南儋州 571737)
钙调蛋白(CaM)作为重要的抗逆信号转导蛋白,在调控木薯抗逆境和块根采后生理腐烂中起重要作用,为给快速检测木薯在不同生长环境中CaM蛋白的表达水平提供优良抗体,克隆木薯CaM基因并将目的基因插入原核表达载体,经Escherichiacoli表达并纯化,用纯化的融合蛋白ACP-CaM免疫Balb/C小鼠,间接ELISA测定小鼠血清效价后,取小鼠脾细胞与SP2/0细胞融合,筛选能产生抗CaM单克隆抗体的杂交瘤细胞株,用Western blot、ELISA等方法对制备的单克隆抗体进行初步鉴定。Western blot分析结果显示原核表达重组质粒在E.coli中能高效表达CaM,免疫小鼠后取效价高的2#小鼠脾细胞和SP2/0细胞融合,共获得7株效价均达到106以上、能稳定分泌抗CaM抗体的细胞株,这7 株单抗与淀粉磷酸化酶、His、BSA均无交叉反应,4D5株与标签蛋白ACP有交叉反应,表明其余6株均为CaM特异性抗体;抗体亚型鉴定结果显示7株单抗均为IgG型抗体。
木薯;钙调蛋白;原核表达;单克隆抗体
木薯(ManihotesculantaCrantz)是世界三大薯类作物之一,也是世界六大农作物之一,是全球超过8亿人的基本食粮。但木薯块根采后2~3 d就快速腐烂,限制木薯的使用并导致严重的经济损失[1-3]。对木薯采后腐烂的研究结果显示,贮存木薯的块根腐烂是一个复杂的蛋白质群协同调控过程,其影响因素包括抗氧化作用、碳水化合物及皮层能量代谢、细胞结构、信号转导、氨基酸代谢、蛋白质生物合成等[2,4]。通过Pathway Studio软件构建差异蛋白调控网络,分析‘华南5号’木薯块根差异蛋白群体在整个调控网络中的关系,结果表明,NADH-氧化酶表达上调产生大量ROS,诱导钙调蛋白(Calmodulin,CaM)上调,启动一系列的生物防御反应,提高抗逆蛋白质表达,从而延长块根贮藏时间,推测CaM在木薯块根采后腐烂中起重要的调节作用[2,4]。
CaM 是广泛存在于各种真核生物细胞中的多功能信号系统调控蛋白,为Ca2+的受体蛋白,虽然参与生物体多种生理活动调控,但它本身不具有酶活性,通过与相应靶蛋白结合发挥生物学效应,引起生物学活性改变参与钙离子相关代谢活动的调控[5],对生物体内多种Ca2+依赖的细胞功能和酶体系都有重要的调节作用,且能敏感捕获任何微量的钙[6]。CaM 在真核细胞的进化过程中高度保守,甚至在脊椎动物中具有完全相同的一级结构;在植物中,CaM的一级结构相似程度也在 90%以上[7],更低级生物中的序列差异越来越大[8]。研究者们已经从不同植物,如小麦、苜蓿、番茄、水稻、拟南芥、马铃薯、扶桑等植物中分离或克隆得到CaM基因[8]。不同植物中存在多种CaM基因表达系统,拟南芥在水分喷射、地下灌溉、机械触动和黑暗等4种外界刺激下,至少启动 4 种CaM基因的表达,刺激 30 min后,CaM的表达水平增加近百倍。但用不同激素刺激,发现CaM的mRNA在大麦糊粉层原生质体细胞质和细胞核中表达没有明显变化,表明激素处理不会影响 CaM 的分布[8]。不同生物或非生物刺激,如病虫害[9-11]、缺氧[12]、高盐[13-14]、重金属[15]和温度[16]胁迫等,可以改变植物CaM基因的表达分布,只是在不同组织和器官中的含量和分布不同[8]。作为抗逆信号转导中重要的蛋白,研究 CaM 含量在不同木薯品种、不同器官和不同时期的含量变化非常必要。笔者首次从木薯中克隆得到CaM基因,体外表达 CaM 蛋白,免疫小鼠制备其单克隆抗体,为快速检测 CaM 在木薯不同品种(系)、不同器官和不同时期的分布和含量变化提供优良抗体,也可为木薯采后生理腐烂调控机制的研究提供材料。
1 材料与方法
1.1 主要原料与试剂
限制性内切酶、TaqDNA聚合酶、DNA连接酶、Marker等均购自北京索莱宝公司;质粒、E.coli来自国家菌种保藏中心;PEG融合剂、小鼠抗体亚型鉴定试剂盒、酰基载体蛋白质(Acyl Carrier Protein,ACP)、淀粉磷酸化酶(Starch phosphorylase,SP)、组氨酸(Histidine,His)、牛血清白蛋白(Bovine serum albumin,BSA)为Sigma公司产品;QuickAntibody-mouse 5w佐剂、Clone easy培养基购自北京博奥龙免疫技术有限公司;HRP-羊抗鼠、DMEM不完全培养基、L-谷氨酰胺、青霉素-链霉素、胎牛血清、小牛血清、HAT、HT、DMSO购自GIBCO公司;吐温-20(Tween-20)、Triton X-100、磷酸二氢钠、磷酸氢二钠、氯化钠、碳酸氢钠、碳酸钠、氯化钾、甘油、丙酮、无水乙醇等化学试剂购自国药集团化学试剂有限公司;ACP单克隆抗体为北京义翘神州生物技术有限公司产品;Babl/C小鼠购自汕头大学医学中心;SP2/0保存于中国热带农业科学院热带作物品种资源研究所农业部木薯种质资源保护与利用重点实验室(以下简称“本实验室”);‘华南5号’木薯保存于农业部国家木薯种质资源圃;淀粉磷酸化酶为本实验室原核表达;KJ-2为本实验室在pET-16b的EcoRⅠ酶切位点后插入ACP-tag后改造得来。试验于2015年3月-2016年6月进行。
1.2 方 法
1.2.1 KJ-2-CaM重组质粒的构建 根据Phytozome database(https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Mesculenta) cassava4.1_018409m 数据库的CaM基因序列设计引物,预期扩增产物长度为450 bp;上游引物ACP-CaM-F:5′-CGCggatccATGGCGACTCAATTCAGCGC-3′; 下游引物ACP-CaM-R:5′-CCGctcgagTTTCGCCATCATTAC TTTCACAA-3′(小写部分为酶切位点),在上下游引物中分别引入酶切位点BamHⅠ/XhoⅠ,以‘华南5号’木薯块根cDNA为模板,将PCR扩增的目的片段回收后,与表达载体KJ-2分别用BamH I/XhoI双酶切,回收,连接,取少量连接产物转入大肠杆菌 DH5 α感受态细胞,接种至LB卡那霉素抗性固体培养基并37 ℃培养过夜,挑取单克隆至卡那霉素LB液体培养基,37 ℃ 、200 r/min培养过夜,取质粒进行测序鉴定,将鉴定正确的质粒命名为KJ-2-CaM。
1.2.2 融合蛋白的原核表达、鉴定与纯化 将测序正确的质粒KJ-2-CaM转化到宿主菌E.coliBL21中,用1.0 mmol/L IPTG诱导表达,收集菌体用超声波破菌仪裂解,离心,分别收集上清和沉淀并进行分析。沉淀用8 mol/L 尿素变性增溶, Ni-NTA亲和柱分离纯化目的蛋白。纯化蛋白依次在浓度为6、4、2、1、0.25、0.1、0 mol/L的尿素溶液中透析复性,每个浓度中透析12 h,复性蛋白分装后 20 ℃冻存,备用。取50 L纯化蛋白加入等体积2×SDS凝胶上样缓冲液,100 ℃煮沸和冰浴各处理4 min,室温10 000 r/min离心5 min,取20 μL进行SDS-PAGE,考马斯亮蓝染色检测纯化效果。采用Western blot以1∶1 000 稀释的抗 ACP 标签单克隆抗体检测重组蛋白的反应原性。
1.2.3 CaM单克隆抗体的制备 将纯化后的融合蛋白ACP-CaM 按每鼠50 g的剂量与QuickAntibody-mouse 5w佐剂等剂量混匀,后腿肌肉注射免疫5只小鼠,第21 天按同样方式再免疫 1 次,第35 天断尾采血,全血37 ℃ 放置 2 h后4 ℃ 放置12 h,4 ℃ 10 000 r/min 离心15 min,收集血清,以纯化后的融合蛋白ACP-CaM作为包被原,ELISA间接法检测血清效价。取50 g 纯化后的融合蛋白ACP-CaM用生理盐水定容至500 L,腹腔冲击免疫抗体效价高的小鼠,冲击免疫后72 h 左右摘眼球取全血分离血清保存,备用。将小鼠在φ=75% 酒精中浸泡10 min,无菌条件下取出小鼠脾脏并分离脾细胞,与 SP2/0 细胞在 PEG 作用下融合,HAT 筛选培养,培养7~10 d后用间接ELISA 检测细胞上清,阳性细胞用有限稀释法克隆直至阳性单克隆率为 100%,小鼠体内诱生腹水法制备抗体,将阳性杂交瘤细胞于液氮中长期保存。
1.2.4 腹水纯化 将新采集的腹水4 ℃、10 000 r/min离心15 min,去沉淀,用0.5 mol/L、pH 7.0的PBS 5倍稀释,在冰浴条件下等体积逐滴加入饱和硫酸铵溶液,冰浴搅拌30 min后4 ℃静置12 h,4 ℃、10 000 r/min离心15 min,弃上清,沉淀用与腹水等体积的0.02 mol/L、pH 7.2 的PBS溶解,冰浴条件下再逐滴加入总PBS体积半量的饱和硫酸铵溶液使之达到33%饱和度,连续搅拌30 min后,4 ℃、10 000 r/min离心15 min,弃上清,沉淀用0.02 mol/L、pH 7.2 的PBS透析72 h后分装, 20 ℃冻存。
1.2.5 抗体鉴定 抗体特异性检测:采用棋盘法测定抗体效价,即将纯化的融合蛋白ACP-CaM用pH 9.6、0.05 mol/L的碳酸盐缓冲液稀释,按照250 ng/mL、500 ng/mL、1 g/mL、2 g/mL的质量浓度梯度包被酶标板,每孔100 μL,37 ℃ 包被2 h,每孔120 μL(含φ=5% 小牛血清的PBST),37 ℃封闭1.5 h,每孔加100 μL梯度稀释的抗体,37 ℃反应30 min,PBST洗涤 4~5 次,每孔加100 μL酶标羊抗鼠二抗,37 ℃反应30 min,PBST 洗涤4~5 次,TMB底物液37 ℃显色反应15 min,0.5 mol/L H2SO4终止反应后酶标仪检测每孔的吸光度(OD)。选择阳性OD值大于阴性OD值2.1倍的抗体最高稀释倍数为抗体的效价。然后分别在酶标板上包被1 g/mL ACP-CaM、ACP、SP、His、BSA,采用ELISA方法检测抗体的特异性。
抗体亚型鉴定:用小鼠抗体亚型鉴定试剂盒检测7株单抗的亚型。
抗体的应用:取‘华南5号’鲜薯块根1 g,液氮中研磨,用丙酮沉淀法提取块根蛋白,金属浴10 min后13 000 r/min室温高速离心取上清,得木薯块根CaM蛋白初提液,CaM蛋白初提液SDS-PAGE后转移至 NC 膜,用 5 g/L脱脂奶粉的 PBST 室温封闭 2 h;加 1∶2 000 稀释的抗CaM单克隆抗体室温轻摇作用 1 h;PBST 洗涤 3 次;加入 1∶3 000 稀释的 AP 标记羊抗鼠 IgG 室温反应 1 h,再PBST 洗涤 3 次;BCIP/NBT 底物显色。
2 结果与分析
2.1 重组质粒的构建
PCR法从木薯块根基因中获得CaM基因片段,经10 g/L琼脂糖凝胶电泳后450 bp的目的条带清晰可见(图1),片段大小与预期相同,提取的阳性克隆质粒经测序鉴定证实序列准确。
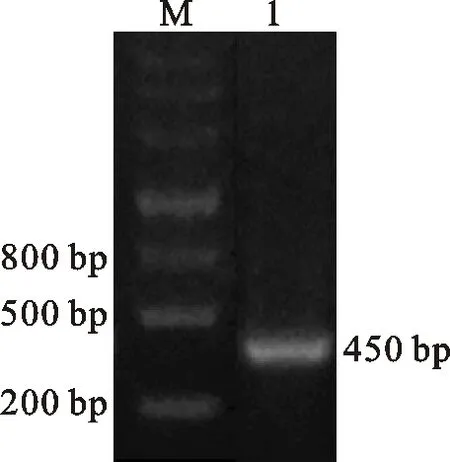
M.Marker;1. PCR回收片段 PCR recovered fragments
2.2 Western blot分析
将KJ-2-CaM质粒转化至E.coliBL21感受态细胞。1.0 mmol/L IPTG诱导5 h,经12% 分离胶SDS-PAGE,诱导后的菌液样品在相对分子质量约为21 ku 处出现目的条带,未经诱导的菌液上清样品则无相应条带(图2)。说明目的蛋白在E.coliBL21中获得表达,且以包涵体表达,主要存在于沉淀中。表达产物的Western blot结果表明(图3),印迹条带约在21 ku处,说明蛋白表达成功。

M.Marker;1.上清液 Supernatant; 2.表达上清液 Expression supernatant; 3.包涵体 Inclusion
图2CaM蛋白的SDS-PAGE检测
Fig.2DetectionofCaMexpressionusingSDS-PAGE

M.Marker;1.表达上清纯化产物 Expression supernatant purified product; 2.上清液 Supernatant
图3CaM蛋白的Westernblot检测
Fig.3DeterminationofCaMexpressionusingWesternblot
2.3 小鼠血清效价检测
用免疫前的小鼠血清作为阴性对照,抗原包被2 g/mL,5只小鼠血清分别梯度稀释,得到5只小鼠血清效价(图4),挑选效价高的2#小鼠进行细胞融合。

图4 小鼠血清的效价Fig.4 Detection result of serum titer of mice
2.4 抗体纯化后效价、亚型测定和特异性测定
融合后的杂交瘤细胞经过2~4次的有限稀释克隆,得到7株阳性单克隆细胞(表1),7株阳性单克隆细胞体外诱导法制备的腹水效价均达到106及以上,抗体亚型均为IgG型。所有抗体均与SP、His、BSA无交叉反应,4D5株与载体ACP有交叉反应,说明4D5株细胞产生的抗体为标签蛋白特异性抗体,其余6株为CaM的特异性抗体。

表1 纯化后的腹水效价和抗体亚型Table 1 Ascites titer and subtypes testing after purified
2.5 单克隆抗体的应用
丙酮沉淀法提取‘华南5号’鲜薯块根CaM蛋白,经Western Blot分析,结果如图5所示,木薯块根样本所在的1~3泳道在16.7 ku处有明显印迹条带,而无关蛋白BSA泳道未出现明显条带,说明该抗体能和木薯中CaM蛋白特异性结合,具有良好的反应原性。

M.Marker;1~3. 木薯块根蛋白质 Proteins extracted from cassava storage root; 4. BSA
图5Westernblot检测木薯块根中CaM的表达
Fig.5CaMexpressionoftuberousrootsincassavaSC5usingWesternblot
3 讨 论
免疫学分析技术已广泛应用于生命科学的各个研究领域,如人类临床诊断中早孕HCG测试[17-18]、乙肝检测[19-21]、肿瘤检测[22]、动物疾病诊断[23-24]、食品安全检测[25-28]等领域。随着木薯全基因组测序工作的完成及基因数据库的构建[29],免疫学技术结合蛋白质组学的研究在木薯高效、高产、特用育种和关键蛋白表达调控机制研究通路中的应用将会越来越重要[1]。
目前,CaM的测定方法主要有免疫学方法、凝胶电泳法和酶法[5]。凝胶电泳法因检测不准确应用较少;酶法能检测有活性的CaM,但是测定过程中容易受到干扰而导致测定值偏低;免疫学方法主要有放射免疫分析法(RIA)和酶联免疫吸附测定法(ELISA),这2种方法检测都是以抗原抗体特异性结合为基础,两者都需要制备CaM抗体[5]。免疫学检测方法特异性强,受待测样本中非CaM成分干扰相对小,能检测CaM总量,但是不能确定CaM活性含量[30]。
CaM是由148个氨基酸残基组成的小分子蛋白,由于其一级结构在进化上高度保守,抗原表位较少,用常规方法极难制备出特异性强的优质抗体[30]。本试验以木薯为研究对象,成功表达木薯CaM融合蛋白,免疫小鼠制备单克隆抗体。由于是用重组表达的ACP-CaM免疫小鼠制备单克隆抗体,小鼠的B细胞中有可能存在抗标签蛋白ACP的抗体,并且单抗制备整个过程也是用ACP-CaM作为包被原来筛选特异性抗体,难免会有单克隆细胞株产生的抗体是针对标签蛋白的,所以在后期进行特异性筛选时,用ACP直接包被ELISA板,检测其和单克隆抗体是否结合,排除1株单克隆细胞株,其余为CaM特异性单克隆抗体。
作为获得单克隆抗体细胞株最关键的一步就是细胞融合,影响细胞融合效率的因素包括SP2/0细胞形态和数量、脾细胞数量、融合时间、融合剂等,为得到更高的融合率,SP2/0在融合前2周复苏,再用8-氮鸟嘌呤处理1周以上,将细胞培养到形态最好的对数生长期再进行融合。培养更多的SP2/0,使融合时B细胞的选择更多,融合成功率大。分离脾细胞后,2 000 r/min离心10 min,虽然相对其他报道,这个转速较高,但是可以获得更多的B细胞,能提高融合的成功率。
在杂交瘤细胞制备过程中,细胞融合、克隆和扩大培养阶段都加入不同浓度含细胞生长因子的饲养细胞替代品Clone easy,以减少制备腹腔细胞作为饲养细胞试验过程中带来的污染以及不杀小鼠取腹腔巨噬细胞体现的动物福利。本研究制备6株能稳定分泌抗CaM蛋白的单克隆杂交瘤细胞株所产生的抗体,为快速检测木薯中不同品种(系)、不同形成时期、不同部位的CaM含量提供优质抗体,有利于Ca2+作为信号分子和Ca2+-CaM复合系统在整个木薯生长发育和块根采后生理代谢中的机制研究。
Reference:
[1] 陈松笔,安飞飞,朱文丽,等.蛋白质组学在木薯育种中的应用[J].生物技术通报,2015,31(11):18-26.
CHEN S B,AN F F,ZHU W L,etal.Applecation of proteomics in cassava breeding[J].BiotechnologyBulletin,2015,31(11):18-26(in Chinese with English abstract).
[2] 简纯平.采后木薯块根贮存能力及蛋白质组学分析[D].海口:海南大学,2013.
JIAN CH P.Storage ability and proteomic analysis of post-harvest tuberous roots of cassava[D].Haikou:Hainan University,2013(in Chinese with English abstract).
[3] GHOSH S P,RAMANUJAM T,JOS J S.etal.Tuber Crops[M].Oxford and IBH Publishing Co.Pvt.Ltd.,New Dehli,1988:3-146.
[4] 张振文.木薯块根采后生理的蛋白质调控机理研究[D].海口:海南大学,2012.
ZHANG ZH W.Postharvest physiology of cassava(ManihotesculentaCrantz) tuberous roots comparative proteomics analysis[D].Haikou:Hainan University,2012(in Chinese with English abstract).
[5] 梁 婧,张晓蓉,许艳华.钙调蛋白的研究进展[J].临床口腔医学杂志,2010,26(8):500-501.
LIANG J,ZHANG X R,XU Y H.The research progress of calmodulin[J].JournalofClinicalStomatology,2010,26(8):500-501(in Chinese).
[6] 罗 梅,董章勇,宾淑英,等.扶桑棉粉蚧钙调蛋白基因的克隆与生物信息学分析[J].华中农业大学学报,2012,31(3):320-324.
LUO M,DONG ZH Y,BIN SH Y,etal.Molecular cloning and bioinformatics analysis of calmodulin genes inPhenacoccussolenopsisTinsley[J].JournalofHuazhongAgriculturalUniversity,2012,31(3):320-324(in Chinese with English abstract).
[7] ZHAO S,CHEN D,GENG Q,etal.The highly conserved LAMMER/CLK2 protein kinases prevent germ cell overproliferation inDrosophila[J].DevelopmentalBiology,2013,376(2):163-170.
[8] 刘 维.番茄钙调蛋白和类钙调蛋白的抗病调控功能分析[D].杭州:浙江大学,2015.
LIU W.Functional analyses of tomato calmodulin and calmodulin-like genes in disease resistance[D].Hangzhou:Zhejiang University,2015(in Chinese with English abstract).
[9] 朱友林,吴健胜,王金生.水稻对白叶枯病菌抗性相关蛋白的双向电泳分析[J].中国农业科学,2000,33(4):91-93.
ZHU Y L,WU J SH,WANG J SH.Analysis of resistance-related proteins in rice againstXanthomonasoryzaepv.oryzaeby two-dimensional electrophoresis[J].ScientiaAgriculturaSinica,2000,33(4):91-93(in Chinese with English abstract).
[10] 陈荣智,翁清妹,黄 臻,等.水稻对褐飞虱抗性相关蛋白的双向电泳分析(英文)[J].植物学报(英文版),2002,44(4):427-432.
CHEN R ZH,WENG Q M,HUANG ZH,etal.Analysis of resistance-related proteins in rice against brown planthopper by two dimensional electrophoresis[J].ActaBotanicaSinica,2002,44(4):427-432(in English with Chinese abstract).
[11] ALI G S ,REDDY V S,LINDGREN P B,etal.Differential expression of genes encoding calmodulin-binding proteins in response to bacterial pathogens and inducers of defense responses[J].PlantMolecularBiology,2003,51(6):803-815.
[12] SHI J X,CHEN S,NATAN G,etal.Effects of anaerobic stress on the proteome of citrus fruit[J].PlantScience,2008,175(4):478-486.
[13] 蔡 伦,张富春,曾幼玲,等.新疆盐生植物的钙调蛋白基因克隆与序列分析[J].植物生理学通讯,2005,41(2):163-167.
CAI L,ZHANG F CH,ZENG Y L,etal.Cloning and sequence analysis of the calmodulin gene from halophytes in Xinjiang[J].PlantPhysiologyCommunications,2005,41(2):163-167(in Chinese with English abstract).
[14] CHEN S,GOLLOP N,HEUER B.Proteomic analysis of salt-stressed tomato(Solanumlycopersicum)seedlings:effect of genotype and exogenous application of glycinebetaine[J].JournalofExperimentalBotany,2009,60(7):2005-2019.
[15] 孙 鹂.铅对细胞内钙离子及钙调蛋白影响的研究进展[J].国外医学卫生学分册,2002,29(6):328-330.
SUN L.Lead to the intracellular calcium ion and the research advances in the effect of calmodulin[J].ForeignMedicalSciences(SectionofHygiene),2002,29(6):328-330(in chinese).
[16] MAJOUL T,BANCEL E,TRIBOI E,etal.Proteomic analysis of the effect of heat stress on hexaploid wheat grain:Characterization of heat-responsive proteins from non-prolamins fraction [J].Proteomics,2004,3(2):175-183.
[17] 王蜜霞,刘军涛,刘儒平,等.基于化学发光磁酶免疫的HCG检测系统研究[J].仪器仪表学报,2010,31(2):312-317.
WANG M X,LIU J T,LIU R P,etal.Development of HCG detecting system based on chemiluminescent magnetic enzyme-linked immunoassay[J].ChineseJournalofScientificInstrument,2010,31(2):312-317(in Chinese with English abstract).
[18] 时洪娟.血清孕酮、HCG检测在先兆流产中的临床诊断意义[J].中国实用医药,2015,10(32):40-41.
SHI H J.Detection of serum progesterone and HCG in the clinical diagnosis value of threatened abortion[J].ChinaPracticalMedical,2015,10(32):40-41(in chinese).
[19] 宋秀丽,唐 慧,丁伶灵,等.2007-2012年中国大学生乙肝表面抗原阳性检出率的meta分析[J].中国微生态学杂志,2013,25(3):264-268.
SONG X L,TANG H,DING L L,etal.A meta-analysis of positive rates of hepatitis B surface antigen in Chinese undergraduate students during 2007 to 2012[J].ChineseJournalofMicroecology,2013,25(3):264-268(in Chinese with English abstract).
[20] 杨 燕.大理市2009年健康人群麻疹、白喉、乙脑及乙肝抗体水平监测结果分析[J].中国卫生检验杂志,2010,20(3):636-637.
YANG Y.An analysis of antibody level of measles diphtheria meningitis B and hepatitis B of healthy population in Dali in 2009[J].ChineseJournalofHealthLaboratoryTechnology,2010,20(3):636-637(in Chinese with English abstract).
[21] 马福宝,张龙华,陈海平,等.母传乙肝抗体水平不同的新生儿接种乙肝疫苗免疫学效果研究[J].实用预防医学,2011,18(11):2079-2080.
MA F B,ZHANG L H,CHEN H P,etal.Study on immunity effects of hepatitis B vaccine for infants with the different levels of maternal transferred antibody[J].PracticalPreventiveMedicine,2011,18(11):2079-2080(in Chinese with English abstract).
[22] 郭亚军.基于单克隆抗体的肿瘤免疫疗法研究进展[J].生物工程学报,2015,31(6):857-870.
GUO Y J.Progress in monoclonal antibody-based immunotherapy for cancer treatmeng[J].ChineseJournalofBiotechnology,2015,31(6):857-870(in Chinese with English abstract).
[23] LLOYD J R,LANDSCHUTZE V,KOSSMANN J.Simultaneous antisense inhibition of two starch synthase isoforms in potato tubers leads to accumulation of grossly modified amylopectin[J].BiochemicalJournal,1999,338(Pt2):515-521.
[24] ABEL G J.SPRINGER F,WILLMITZER L,etal.Cloning and functional analysis of a cDNA encoding a novel 139 kDa starch synthase from potato(SolanumtuberosumL.) [J].PlantJournal,1996,10(6):981-991.
[25] AKATSUKA T,NELSON O E.Starch granule-bound adenosine diphosphata glucose-starch glucosyltransferases of maize seeds[J].JournalofBiologicalChemistry,1966,241(10):2280-2285.
[26] OZBUN J L,GREENBERG E,PREISS J,etal.Starch synthetase,phosphorylase,ADP glucose pyrophosphorylase,and UDP glucose pyrophosphorylase in developing maize kernels[J].PlantPhysiology,1973,51(1):1-5.
[27] DUWENIG E,STEUP M,KOSSMANN J.Induction of genes encoding plastidic phosphorylase from spinach(SpinaciaoleraceaL.)and potato(SolanumtuberosumL.) by exogenously supplied carbohydrates in excised leaf discs[J].Planta,1997,203(1):111-120.
[28] 王妍入,李培武,张 奇,等.噬菌体展示纳米抗体模拟黄曲霉毒素抗原的活性表征[J].中国农业科学,2014,47(4):685-692.
WANG Y R,LI P W,ZHANG Q,etal.Characterization of a phage-displayed nanobody imitating aflatoxin antigen[J].ScientiaAgriculturaSinica,2014,47(4):685-692(in Chinese with English abstract).
[29] WANG W,FENG B,XIAO J,etal.Cassava genome from a wild ancestor to cultivated varieties[J].NatureCommunications,2013,10(5):5110.
[30] 王 娟,李晓军,潘继文,等.钙调素ELISA定量检测方法的建立与实验条件优化[J].医学研究生学报,2003,16(6):401-403.
WANG J,LI X J,PAN J W,etal.Development and optimization of quantitative ELISA for calmodulin[J].JournalofMedicalPostgraduates,2003,16(6):401-403(in Chinese with English abstract).
ProkaryoticExpressionandMonoclonalAntibodyPreparationofCalmodulinOriginatedfromCassava
OU Wenjun,YU Houmei,AN Feifei,LUO Xiuqin,QIN Yuling and CHEN Songbi
(Tropical Crops Genetic Resources Institute,Chinese Academy of Tropical Agricultural Sciences /Key Laboratory of Ministry of Agriculture for Germplasm Resources Conservation and Utilization of Cassava,Danzhou Hainan 571737,China)
Calmodulin(CaM) is an important signal-transduction protein. It will play an important role in regulating tolerance to extreme conditions and postharvest physiology deterioration. In order to provide good antibody in rapid determination of CaM expression in cassava growing at different environments,in the present study,cDNA of cassava CaM was cloned into the prokaryotic expression vector,CaM was expressed inE.coliand then purified. The purified fusion protein ACP-CaM was used to immunize Balb/c mice and the titer of mice serum was determined by indirect ELISA. The spleen lymphocyte was fused with SP2/0 hybridoma to produce monoclonal antibody against CaM. Western blot and ELISA were carried out to preliminarily identify the monoclonal antibody produced by these hybridomas. Western blot result showed that recombinant plasmid inE.colican expressed CaM efficiently. SP2/0 cells and spleen cells of 2# mice were fused,then 7 strains,which antibody titer was more than 106,were
,no cross reaction with starch phosphorylase,His and bovine serum albumin. 4D5 strains has cross reaction with carrier protein ACP,6 strains of CaM were specificity antibodies. All of strains were IgG type.
Cassava(ManihotesculantaCrantz); Calmodulin; Prokaryotic expression; Monoclonal antibody
2016-08-03
2016-11-10
Central Public-interest Scientific Institution Basal Research(No. 0315014); NSFC-CGIAR International(Regional) Cooperation and Exchange Programs(No.31361140366);Chinese Cassava Agro-technology Research System(No.CARS-12).
OU Wenjun,male,associate researcher,Ph.D.Research area:cassava germplasm resources. E-mail:cassava6973@163.com
Q94-336
A
1004-1389(2017)09-1317-07
(责任编辑:顾玉兰Responsibleeditor:GUYulan)

西北农业学报 2017,26(9):1324⁃1333ActaAgriculturaeBoreali⁃occidentalisSinicadoi:10.7606/j.issn.1004-1389.2017.09.009
日期:2017-09-12
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170912.1740.018.html
2016-08-03
2016-11-10
中央级公益性科研院所基本科研业务费专项(0315014);NSFC-CGIAR国际合作重点项目(31361140366);现代农业产业技术体系专项资金(CARS-12)。
欧文军,男,副研究员,博士,研究方向为木薯种质资源。E-mail:cassava6973@163.com
陈松笔,男,研究员,研究方向为木薯蛋白质组学和作物遗传育种。E-mail:songbichen@catas.cn
CorrespondingauthorCHEN Songbi,male,research fellow.Research area:cassava proteomics and crop genetics and breeding. E-mail: songbichen@catas.cn
