大肠埃希菌O111黏附牦牛子宫内膜上皮细胞致炎模型的建立
2017-09-25吴庆侠董海龙朱洪云刘忠艳西藏农牧学院西藏林芝860000
吴庆侠,董海龙,朱洪云,刘忠艳(西藏农牧学院,西藏林芝 860000)
大肠埃希菌O111黏附牦牛子宫内膜上皮细胞致炎模型的建立
吴庆侠,董海龙,朱洪云,刘忠艳
(西藏农牧学院,西藏林芝 860000)
为建立大肠埃希菌O111黏附牦牛子宫内膜上皮细胞的致炎模型,体外分离、培养牦牛子宫内膜上皮细胞,培养5 d后,加入大肠埃希菌O111,通过计算细胞上细菌的黏附数量研究细菌数量和孵育时间对黏附效果的影响,得到最佳黏附条件,通过ELISA法测定细胞培养上清液中肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6)、白细胞介素-1β(IL-1β)的质量浓度变化来鉴定模型。结果表明,大肠埃希菌为1×105CFU/mL、孵育时间为120 min是黏附的最佳条件。ELISA 法检测结果表明,与对照组相比,60、90、120 min组TNF-α、IL-6、IL-1β的质量浓度显著升高,致炎效果明显,表明致炎模型建立成功。
大肠埃希菌O111;牦牛子宫内膜上皮细胞;致炎模型
奶牛子宫内膜炎是奶牛产后的常发病,大肠杆菌是其致病的一种重要的条件性致病菌,发病率在15%左右[1],严重危害奶牛产业的经济效益。大肠埃希菌O111是具有高致病力的菌株,王孝武等[2]研究表明,从患病奶牛子宫内膜10 份样本中分离出12 株细菌,其中7 株为大肠杆菌;杭苏琴等[3]研究表明,从患有子宫内膜炎的奶牛子宫黏液中分离大肠杆菌,血清学分型结果表现为O111型占41.84%,占有比例最高。
对于奶牛子宫内膜炎的研究,国内外普遍采用体外培养子宫内膜细胞,建立相应炎症模型的方式[4]。目前,牦牛作为藏区牧民的主要生活资料,同样面临产后子宫内膜炎的困扰,临床观察统计,西藏林芝地区牦牛子宫内膜炎的发病率已达到18%,尤其是胎衣不下所引起的子宫内膜炎尤为多见。羊云飞[5]通过16S rRNA对藏区牦牛源大肠杆菌进行分离、血清学鉴定,发现O111所占比例较高,达到13.6%。然而体外建立O111菌株感染牦牛子宫内膜细胞的致炎模型未见报道。
本研究通过体外分离、培养的牦牛子宫内膜上皮细胞为模型,分别评价大肠埃希菌O111的数量,以及子宫内膜细胞孵育时间对该菌黏附性的影响,最后通过双抗体夹心ELISA法检测细胞培养上清液中肿瘤坏死因子α(TNF-α)、白细胞介素-6(IL-6)、白细胞介素-1β(IL-1β)的质量浓度变化,从而成功构建模型,为进一步研究其致病及抗病机制提供条件。
1 材料与方法
1.1 材 料
1.1.1 牦牛子宫和菌种来源 牦牛子宫取自西藏林芝市巴宜区牦牛屠宰厂。大肠埃希O111(Escherichiacoli,ATCC43887)由中国农业大学王九峰教授惠赠。
1.1.2 菌种与试剂 D-MEM/F12培养液、胎牛血清、胶原酶Ⅱ购自Gibco公司;TNF-α、IL-6、IL-1β检测试剂盒均购自博士德生物;双抗(青霉素和链霉素)为哈药集团生产;营养肉汤、PBS缓冲液均为自配。
1.1.3 主要仪器和设备 6 孔和96 孔细胞培养板为Costar公司产品;CO2培养箱(上海福玛试验设备有限公司)、酶标仪(美国伯腾仪器有限公司)、倒置生物显微镜(Nikon公司)、离心机(Sigma公司)、双人超净工作台(苏净净化设备有限公司)。
1.2 方 法
1.2.1 大肠埃希菌O111悬液制备 将大肠埃希菌O111于营养肉汤内培养,37 ℃振荡12 h后取出,3 000 r/min离心10 min,去上清液,使用无菌PBS缓冲液悬浮菌种,再次3 000 r/min离心10 min,去上清液,使用D-MEM/F12培养液将菌种重悬,并将其调至1×107CFU/mL,备用。
1.2.2 牦牛子宫内膜上皮细胞的培养 参照吴庆侠等[6]的方法进行,具体操作如下:将采取的牦牛子宫于含有双抗的无菌PBS溶液中反复冲洗后剪开子宫角,剪取子宫内膜层,把剪取好的子宫内膜层用无菌PBS溶液反复冲洗,直至溶液清亮。将其放入空的血清瓶中,剪成肉糜状以便消化,用无菌PBS溶液冲洗数次后使用2 g/L的胶原酶Ⅱ消化4 h,消化过程中注意每隔30 min摇动消化瓶1次,消化结束后,74 μm滤网过滤消化液,400 r/min离心5 min,收集沉淀(主要为上皮细胞),无菌PBS溶液重悬后自然沉降,下方的沉降物即为纯度较高的上皮细胞。去上清液,使用含φ= 5%胎牛血清的D-MEM/F12培养液制成细胞悬液,移入到预先放置有无菌盖玻片(22 mm×22 mm)的6 孔板中,每孔2 mL,培养5 d后,观察细胞形态,爬满盖玻片时进行黏附试验。
1.2.3 大肠埃希菌O111对牦牛子宫内膜上皮细胞的黏附 大肠埃希菌不同数量分组黏附:使用D-MEM/F12培养液,将“1.2.1”中备用的菌液调整细菌数量为1×107、1×106、1×105、1×104和1×103CFU/mL,D-MEM/F12培养液作为空白对照,依次加入到“1.2.2”中培养好的牦牛子宫内膜上皮细胞进行黏附试验。CO2培养箱中孵育2 h。孵育结束后,取出盖玻片,使用无菌PBS缓冲液冲洗5 次,去除杂质和未黏附细菌等。自然晾干或低温烘干,甲醇固定20 min,革兰氏染色观察大肠埃希菌O111对牦牛子宫内膜上皮细胞的黏附性。每组在显微镜下随机挑选10 个视野,统计每个视野下每个细胞黏附细菌的数量。
孵育不同时间分组黏附:将“1.2.1”中备用的细菌数量为1×107CFU/mL的菌液,加入到“1.2.2”中培养好的牦牛子宫内膜上皮细胞进行黏附试验。孵育时间在30、60、90、120和150 min时分别取样,同“大肠埃希菌不同数量分组黏附”试验中计算方法,计算细胞上细菌的黏附数量。1.2.4 黏附过程中TNF-α、IL-6、IL-1β的质量浓度检测 选取已培养好上皮细胞的6孔板2个,取其中10 孔取上清液,每孔取3 次,每次取0.1 mL,保证每个检测因子有10 个待检样,将上清液移入到96 孔板中,作为对照组待测。将数量为1×107CFU/mL的菌液加入2 个培养好的牦牛子宫内膜上皮细胞6 孔板中进行孵育,孵育时间在60、90、120 min时选取其中10 孔采取上清液,每孔取3 次,每次0.1 mL,同样保证每个检测因子有10 个待检样,将上清液移入到96 孔板中待测,采用双抗体夹心ELISA法测定上清液中TNF-α、IL-6、IL-1β的质量浓度,检测严格按照说明书进行。
1.2.5 数据的统计分析 试验结果重复3 次,采用SPSS 21.0统计软件,对试验数据进行统计分析,结果以“平均数±标准差”表示,用 SigmaPlot 12.5软件绘制柱状图。
2 结果与分析
2.1 牦牛子宫内膜上皮细胞的培养
分离的上皮细胞经培养18 h后开始贴壁,以细胞团为中心向四周辐射生长(图1)。5 d后,上皮细胞在培养皿底部呈铺路石样排列生长(图2)。上皮细胞纯度较高,看不到明显杂质或其他细胞。

图1 培养18 h后上皮细胞生长形态(100×)Fig.1 Growth morphology of epithelialcells after 18 h culture(100×)
2.2 不同细菌数量对黏附效果的影响
在染色前,通过倒置显微镜观察发现,随着添加大肠埃希菌O111数量的增加,部分细胞变圆脱壁,脱壁细胞的数量随着细菌数量的增加而增加;仍然贴壁的细胞形态上没有明显变化。大肠埃希菌对牦牛子宫内膜上皮的黏附性,随着细菌数量的增加,每个细胞上平均黏附的细菌数呈逐步增加(图3和图4),大肠埃希菌数量为1×103、1×104、1×105CFU/mL时黏附细菌的数量差异显著(P<0.05),而大肠埃希菌数量为1×105、1×106、1×107CFU/mL时黏附细菌数量的组间差异不显著(P>0.05)。提示细菌数量为1×105CFU/mL时,黏附细菌数开始稳定,单个牦牛子宫内膜上皮细胞的黏附细菌数量已经达到最大,使大肠埃希菌O111黏附量趋于饱和,因此最佳黏附效果的细菌数量为1×105CFU/mL。
2.3 不同孵育时间对黏附效果的影响
如图5所示,随着时间的增加,黏附细菌数量逐步增加,孵育时间为30、60、90和120 min时黏附细菌的数量差异显著(P<0.05),孵育120 min与150 min相比较,无显著差异(P>0.05),提示在孵育120 min时,黏附细菌数量趋于稳定,黏附细菌量达到最大,因此最佳黏附的孵育时间为120 min。
图2 培养5 d后上皮细胞生长形态(200×)Fig.2 Growth morphology of epithelialcells after 5 d culture(200×)

A~C.大肠埃希菌O111的数量依次为103、104、105CFU/mL Concentration ofEscherichiacoliO111is 103CFU/mL,104CFU/mL,105CFU/mL
图3不同数量大肠埃希菌O111在牦牛子宫内膜上皮细胞的黏附(革兰氏染色,1000×)
Fig.3AdhesionofEscherichiacoliO111onendometrialepitheliumcellsofyakatdifferentconcentrations(Gram’sstaining,1000×)

不同小写字母表示差异显著(P<0.05),下同。
Different lowercase letters are significantly differente at level ofP<0.05,the same below.
图4不同数量的大肠埃希菌O111对牦牛子宫内膜上皮细胞黏附效果
Fig.4EffectsofdifferentnumbersofEscherichiacoliO111onadhesiontoendometrialepitheliumcells
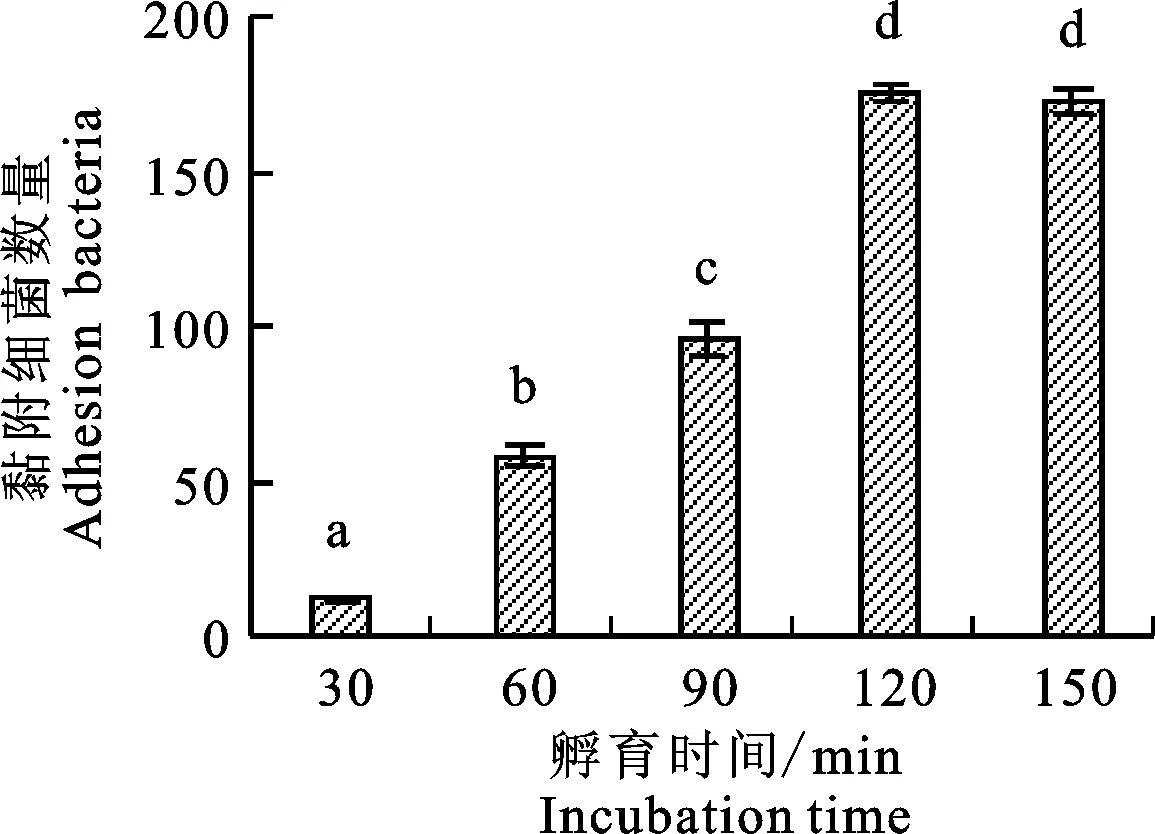
图5 不同孵育时间的黏附效果Fig.5 Effects of different incubation time on adhesion
2.4ELISA法检测细胞培养上清液中TNF-α、IL-6、IL-1β的质量浓度变化
由表1可知,90 和120 min组比较,上清液中TNF-α、IL-6、IL-1β的质量浓度均差异不显著(P>0.05),而60、90、120 min组与对照组均差异显著(P<0.05),提示在大肠埃希菌的作用下,炎性反应变化明显。
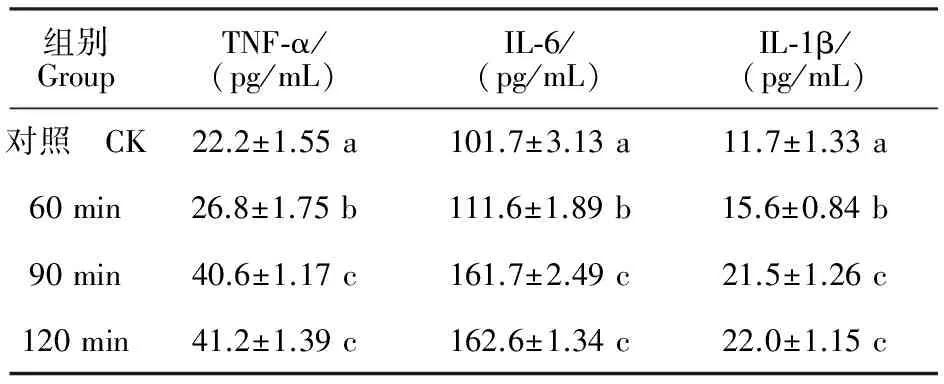
表1 不同时间内TNF-α、IL-6、IL-1β的质量浓度Table 1 Mass concentration changes of TNF-, IL-6 and IL-1 in different time
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters in the same column are significantly different at level ofP<0.05.
3 讨 论
在生殖系统中,子宫占据着重要地位,其内膜上皮直接接触微生物,起到免疫作用,有研究表明,子宫内膜上皮细胞与免疫细胞一样,具有分泌和免疫应答的功能[7],能够释放炎症介质。近年来人们对牛子宫相关炎性疾病的研究都涉及到体外细胞的培养技术,本试验中,牦牛子宫内膜上皮细胞的培养参照吴庆侠等[6]的培养方法,得到纯度较高的上皮细胞,并使用原代细胞进行试验。
大肠埃希菌是子宫内膜炎的重要致病菌,目前体外子宫内膜致炎模型的建立,多是使用大肠杆菌脂多糖(LPS)进行诱导,模型中LPS的致炎效果均很明显,已成功建立的模型有人[8]、小鼠[9]、奶牛[10]、山羊[11]等,这些模型的建立有的是为揭示致炎症发生过程中子宫内膜细胞相关炎性因子的一系列表达变化[12],有的是用于检验药物或者中药提取物对子宫内膜炎的抑制效果等。而直接使用细菌对体外培养的细胞进行黏附的试验很少,本试验直接使用大肠埃希菌对体外培养的牦牛子宫内膜细胞进行黏附建立致炎模型,主要是天然模拟大肠埃希菌感染细胞的过程,从而探讨其定植机制。余抒等[13]研究发现大肠埃希菌黏附HeLa细胞的最佳条件是加入细菌量为1×105CFU/mL,孵育时间为4 h,本试验结果表明,最佳的加入细菌量也为1×105CFU/mL,与其结果一致,但孵育2 h即达到最佳效果,最佳孵育时间有所不同,分析可能是由于黏附的细胞对象不同所造成的,本试验所用细胞为原代子宫内膜上皮细胞,而余抒等[13]试验使用的是瘤细胞,瘤细胞的细胞表面所表达的一系列因子水平与常规细胞相比有很大差异。
Fischer等[14]研究奶牛在产后一定时间内,子宫内膜上皮细胞中相关炎性因子基因表达变化与临床型和隐性型子宫内膜炎的关系,认为炎性趋化因子(CXCL5)、IL-1β、IL-6和TNF的基因可作为隐性子宫内膜炎诊断和治疗监测的标志基因。Ghasemi等[15]对患有亚临床型子宫内膜炎的奶牛血液中TNF-α、IL-6等进行检测,结果其表达水平远远高于健康奶牛,赵立香等[16]的试验表明,通过直接对奶牛子宫内灌注大肠杆菌菌液导致TNF-α、IL-6、IL-1β等表达增多,于晓红等[17]试验表明在LPS的作用下奶牛子宫内膜上皮细胞TNF-α、IL-6、IL-1β表达量均有不同程度的增加。本试验结果也表明,在大肠埃希菌的作用下,上清液中TNF-α、IL-6、IL-1β的质量浓度与对照组相比均显著升高(P<0.05),与相关报道[16-17]一致。
本试验确定大肠埃希菌O111对体外培养的牦牛子宫内膜上皮细胞黏附最佳条件,即细菌数量为1×105CFU/mL,孵育时间为120 min,同时使用ELISA法检测细胞培养上清液中TNF-α、IL-6、IL-1β的质量浓度变化,表明在大肠埃希菌的作用下,炎性反应明显,从而成功建立体外细菌黏附子宫内膜致炎模型。该模型将进一步用于大肠杆菌致牦牛子宫内膜炎症机理及益生菌防治牦牛子宫内膜炎的研究。
Reference:
[1] LEBLANC S J,OSAWA T,DUBUC J.Reproductive tract defense and disease in postpartum dairy cows[J].Theriogenology,2011,76(9):1610-1618.
[2] 王孝武,王旭荣,杨志强,等.甘肃省某牛场奶牛子宫内膜炎病原菌的分离鉴定及其耐药性分析[J].西北农业学报,2014,23(6):16-21.
WANG X W,WANG X R,YANG ZH Q,etal.Isolation,identification and drug-resistance of pathogenic bacteria from the dairy cows infected by edometritis in gansu[J].ActaAgriculturaeBoreali-occidentalisSinica,2014,23(6):16-21(in Chinese with English abstract).
[3] 杭苏琴,王一昊,郑 婕,等.产后奶牛子宫内大肠杆菌的分离和血清型鉴定[J].畜牧与兽医,2015,47(3):104-106.
HANG S Q,WANG Y H,ZHENG J,etal.Isolation and identification ofEscherichiacolifrom the postpartum dairy cows[J].AnimalHusbandry&VeterinaryMedicine,2015,47(3):104-106(in Chinese ).
[4] DACIES D,MEADE K G,SHAN H,etal.Toll-like receptor and antimicrobial peptide expression in the bovine endometrium[J].ReproductiveBiology&Endocrinology,2008,6(1):1-12.
[5] 羊云飞.牦牛、牧民源大肠杆菌分离鉴定、耐药基因检测、PFGE分析及耐药性传递的研究[D].成都:四川农业大学,2011.
YANG Y F.Study on detection of sulfonamides resistance and florfeniol resistance genes and PFGE ofE.colifrom yak and herdsman[D].Chengdu:Sichuan Agricultural University,2011(in Chinese with English abstract).
[6] 吴庆侠,董海龙,芮亚培.牦牛子宫内膜腺上皮细胞分离培养方法比较[J].安徽农业科学,2011,39(18):11085-11087.
WU Q X,DONG H L,RUI Y P.Comparison on in-vitro culture methods of yak endometrial gland epithelial cells[J].JournalofAnhuiAgricultureScience,2011,39(18):11085-11087(in Chinese with English abstract).
[7] LAGOW E,DESOUZA M M ,CARSON D D.Mammalian reproductive tract mucins[J].HumanReproductionUpdate,1999,5(4):280-292.
[8] JING Y Y,HAN Z P,SUN K,etal.Toll-like receptor 4 signaling promotes epithelial-mesenchymal transition in human hepatocellular carcinoma induced by lipopolysaccharide[J].BMCMedicine,2012,10(1):98.
[9] 杨 倩,史万玉,赵驻军,等.小鼠子宫内膜细胞炎症模型的建立[J].河北农业大学学报,2011,34(6):87-90.
YANG Q,SHI W Y,ZHAO ZH J,etal.Inflammatory model of endometrial cells in mice[J].JournaofAgriculturalUniversityofHebei,2011,34(6):87-90(in Chinese with English abstract).
[10] 张桂林,崔晓妮,曹荣峰.脂多糖诱导奶牛子宫内膜上皮细胞核因子kB的表达[J].中国兽医科学,2010,40(10):1063-1066.
ZHANG G L,CUI X N,CAO R F.Expression of NF-kB LPS-induced in bovine endometrial epithelial cel[J].ChineseVeterinaryScience,2010,40(10):1063-1066(in Chinese with English abstract).
[11] 邝晓娇,张世栋,董书伟,等.丹参水提物对山羊子宫内膜上皮细胞炎症模型中基质金属蛋白酶-2表达的影响[J].动物医学进展,2015,36(3):54-59.
KUANG X J,ZHANG SH D,DONG SH W,etal.Effect of extract of salvia miltiorrhiza on MMP-2 in endometrium epithelial cell inflamamation model of goat[J].ProgressinVeterinaryMedicine,2015,36(3):54-59(in Chinese with English abstract).
[12] SWANGCHANUTHAI T,LAVENDER CRM,CHENG Z,etal.Time course of defense mechanisms in bovine endometrium in response to lipopolysaccharide[J].Biologyofreproduction,2012,87(6):1-13,135.
[13] 余 抒,顾 江,曾 浩,等.肠出血性大肠埃希菌O157∶H7黏附HeLa细胞模型的建立[J].第三军医大学报,2007,29(20):1933-1935.
YU SH,GU J,ZENG H,etal.Establishm ent of an enterohem orrhagicEscherichiacoliO 157∶H7 attached HeLa cell model[J].ActaAcademiaeMedicinaeMilitarisTertiae,2007,29(20):1933-1935(in Chinese with English abstract).
[14] FISCHER C,DRILLICH M,ODAU S,etal.Selected pro-inflammatory factor transcripts in bovine endometrial epithelial cells are regulated during the oestrous cycle and elevated in case of subclinical or clinical endometritis[J].ReproductionFertility&Development,2010,22(5):818-829.
[15] GHASEMI F,GONZALEZ-CANO P,GRIEBEL PJ,etal.Proinflam-matory cytokine gene expression inendometrial eytobrush samples harvested from cows with and withour sublinical endome-tritis[J].Theriogenology,2012,78(7):1538-1547.
[16] 赵立香,郭梦尧,李丰阳,等.大肠杆菌引发奶牛子宫内膜炎的致病机制研究[J].黑龙江畜牧兽医,2013(10):14-17.
ZHAO L X,GUO M Y,LI F Y,etal.Research on the pathogenic mechanism of endometritis caused byEscherichiacoliin dairy cattle[J].HeilongjiangAnimalScienceandVeterinaryMedicine,2013(10):14-17(in Chinese with English abstract).
[17] 于晓红,白云静,程 鹏,等.苦参碱对奶牛子宫内膜上皮细胞抗炎作用及其机制[J].北京农学院学报,2015,30(3):35-39.
YU X H,BAI Y J,CHENG P,etal.Anti-inflammatory effect of matrine on the bovine endometrial epithelial cell and its mechanism[J].JournalofBeijingUniversityofAgriculture,2015,30(3):35-39(in Chinese with English abstract).
EstablishmentofInflammationModelsforEscherichiacoliO111AdheredtoYakEndometrialEpithelialCells
WU Qingxia,DONG Hailong,ZHONG Hunyun and LIU Zhongyan
(Xizang Agricultural and Animal Husbandry College,Linzhi Tibet 860000,China)
The yak endometrial epithelial cells were isolated and cultured in vitro to establish inflammation models caused byE.coliO111adhesion yak endometrial epithelial cells. After 5 days’ culture,EscherichiacoliO111was added in, the optimum condition was obtained by calculation of bacteria number to study the cell adhesion effect under different conditions of bacterial number and incubation time. The optimization model was identified by DAS-ELISA method to determine the content change of tumor necrosis alpha (TNF-α), leukocyte mediated IL-6 and IL-1 factors in cell culture supernatant. The results showed that the number ofE.coliwas 1×105CFU/mL, and the incubation time was 120 min, which was the best condition for adhesion. ELISA test results proved that the TNF-α, IL-6, IL-1β mass concentration increased significantly and had obvious inflammation effect in 60 min, 90 min and 120 min test groups than control group. The model of inflammation was established successfully after series of test.
EscherichiacoliO111; Yak endometrial epithelial cells; Inflammatory model
2016-07-05
2016-08-02
Tibet Natural Science Foundation(No.2015212-14-34);Funded by Key Lab Construction Program of Clinical Veterinary Medicine for Medical Universities and Colleges in Tibet.
WU Qingxia, female, associate professor. Research area:animal clinical disease.E-mail:goodwqx@163.com
S857.23
A
1004-1389(2017)09-1289-06
(责任编辑:顾玉兰Responsibleeditor:GUYulan)
日期:2017-09-12
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170912.1740.008.html
2016-07-05
2016-08-02
西藏自治区自然科学基金(2015212-14-34);西藏自治区临床兽医学高校重点试验室建设资助项目。
吴庆侠,女,副教授,从事动物临床疾病研究。E-mail:goodwqx@163.com
董海龙,男,副教授,主要从事产科疾病相关研究。E-mail: 984718586@qq.com
CorrespondingauthorDONG Hailong, male, associate professor. Research area:animal obstetric diseases. E-mail:984718586@qq.com
