基于静态模拟模型的青藏高原东缘高寒草甸区草畜平衡研究
2017-09-20刘海波李林渊蒲小剑杜文华
刘海波,李林渊,蒲小剑,杜文华
(1.甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070; 2.重庆西藏中学,重庆 400036)
基于静态模拟模型的青藏高原东缘高寒草甸区草畜平衡研究
刘海波1,李林渊2,蒲小剑1,杜文华1
(1.甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070; 2.重庆西藏中学,重庆 400036)
以澳大利亚反刍动物营养标准为参考,Stage one模型为框架,家庭牧场为研究尺度,从家畜营养角度出发,将代谢能(Metabolisable energy,ME)和氮维持需要量作为放牧家畜的营养单位,建立营养模型,模拟该地区典型家庭牧场欧拉型藏羊能、氮需求的月际变化及冬季暖棚利用对于藏羊体况维持的作用,研究青藏高原东缘高寒草甸区典型藏羊生产体系下的草畜平衡问题。结果表明:高寒草甸草地牧草消化率高,能量浓度高,高放牧强度(> 3.5 头/hm2)下,藏羊仍能获得足够的代谢能用于增重,而冷季由于草地消化率低,能量浓度差,在低放牧强度(<3.5头/hm2)即使放牧率低于暖季藏羊摄取能量仍不足以满足维持代谢,表现为减重(成年母羊平均5 kg/头)。冷季低温造成典型牧户藏羊畜群维持代谢能每羊单位提高22%,且怀孕后期(1到2月)母羊能量需求高,草畜能量不平衡严重,因此,冷季补饲应以精饲料为主。暖棚可降低家畜御寒代谢能消耗,并降低冷季维持代谢能水平,有助于减少家畜掉膘,典型牧户藏羊畜群维持代谢能1到4月可分别降低1.08、1.23、1.28、1.26 MJ/(头/d)。冷季精饲料补饲+暖棚是较为可行的管理方式。
高寒草甸;欧拉羊;代谢能;维持氮平衡
随着人们饮食习惯的改变,不断增长的人口数量给有限的农用土地和自然资源带来了巨大压力。预计到2050年,全球人口将增至93亿,农产品产量(食物供给)需要在现有基础上提高60%,预计每年产量达85亿t[1]。要实现这一目标,一是增加生产资源(农用土地面积,家畜数量等),二是提高现有资源的生产力,或者单位面积产量,即提高生产效率[2]。农业生产与迅速扩展的城市化进程对有限土地和水资源的竞争空前激烈。而人口、技术进步等因素,也与气候变化互相作用,给农业和环境可持续发展带来了极大挑战。
在全球范围内,随着人们对畜产品需求的不断增长,小型家庭农场成为保证全球动物蛋白安全的重要组成部分。全球约13亿的小农户人口,为发展中国家提供了80%的粮食供给[3]。小型农场为全球生产肉19%、奶12%[4]。作为最大的土地利用单元,家畜生产的管理是解决可持续供养世界人口问题的重要方法来源[3]。自绿色革命以来,单一追求高产的集约化模式已经无法满足未来可持续农业发展的需要。在全球气候变化的大环境下,小农户农业生产对于气候变化更为敏感。有不少学者预测未来10年发展中国家小农户作物产量和畜产品产量都可能因全球环境变化而减少[5]。也有学者表示,一些热点区域的小农户在合理利用农业生态学方法的条件下,未来10年有可能增加一倍的产量[6]。无论未来的前景如何,小农户生产体系的集约化发展一定要在可持续发展的框架下,要注意最大化外部投入(无机化肥等)和修复内部资源(环境服务等)之间的平衡[7]。
青藏高原是典型的以家庭牧场为单位的小农户粗放经营的代表。青藏高原敏感脆弱的生态系统发挥着重要的生态服务功能,同时也维持着世界最大游牧人口的生计(超过980万藏区牧民)[8]。近年来由于气候变暖、超载过牧等影响,青藏高原草地退化问题日益严重。超载过牧也被认为是引起草地退化的主要原因。有学者总结青藏高原草地家畜系统可持续发展的最大问题就是饲草供应和家畜生产的矛盾[8]。青藏高原放牧生态系统中草畜平衡是核心[9]。很多学者利用不同研究手段研究了青藏高原的草畜平衡问题[10-14]。但是研究存在以下问题:1)研究尺度不统一,没有考虑家畜营养需求的动态变化,这会造成研究结果的偏差;2)部分研究仅从能量角度出发,缺乏对蛋白营养的讨论;3)未能选择适宜的能量体系,能氮估算存在偏差。目前以家庭牧场为研究尺度,从家畜营养角度出发,将代谢能(ME)作为放牧家畜的营养单位,利用营养模型模拟青藏高原东缘高寒草甸区典型家庭牧场家畜能、氮需求变化和草地供能的研究较少。笔者以甘肃省玛曲县典型家庭牧场为研究对象,以澳大利亚2007年发表的反刍动物营养标准为参考[15](以下简称为CSIRO),以Stage one模型[16]为框架,研究欧拉型藏羊生产体系能、氮需求的月际变化,以期为青藏高原东缘高寒草甸区欧拉型藏羊生产体系的草畜平衡问题提供基础研究资料。
1 材料和方法
1.1试验区概况
试验地位于青藏高原东北部,甘肃省甘南藏族自治州玛曲县欧拉乡,E 101.88823°,N 33.86721°。海拔3 650 m,年均温1.1℃,极端最低温-29.6℃,无绝对无霜期,年日照时长2 583.9 h,辐射33 kJ/cm2,降水量615.5 mm,蒸发量1 353.4 mm。气候类型为高寒阴湿高原大陆性季风气候。植被类型为高寒草甸,优势牧草主要有莎草科的线叶嵩草(Kobresiacapillifolia)、矮生嵩草(Kobresiahumilis)、矮藨草(Scirpuspumilus)以及禾本科的异针茅(Stipaaliena)、垂穗披碱草(Elymusnutans)、匍匐翦股颖(Agrostisstolonifera)、羊茅(Festucaovina)、波伐早熟禾(Poapoophagorum)等[17]。牧草于5月上旬返青,9月中旬枯黄,绿草期约120 d,枯草期长达240 d[18]。家畜5月底由冬春草场转入夏秋草场,10月底再转入冬春草场。
1.2基础数据调查与模型构建
于2010年开始连续2年对选定的8个家庭牧场进行问卷调查,调查内容包括草地面积、羊只数量、生产投入、市场价格、棚圈建设、畜群管理、补饲和防疫等。基于这些生产基础性数据,建立一个典型家庭牧场(表1)。

表1 模型有关的典型家庭牧场基础信息和初步设定
注:母羊数量包含育成羊数量;产羔时间为2~3月。羔羊自然断奶,试验中为了模型运行方便,统一设定为6个月。标准参考体重指骨骼发育完全,且膘情指数为体况中等欧拉羊的活体重(去除毛重和孕体重量)
1.3模型基础数据测定
地上生物量:2011~2012年,在典型家庭牧场的夏秋和冬春草场,根据草场地形,分别于家畜转场前(6月和10月中旬),在坡顶、坡中和坡底随机设置1.5 m×1.5 m围笼各3个,以限制家畜采食。7~10月在夏秋场的3个围笼内分别选取0.25 m2样方, 1~6月在冬春场的3个围笼内分别选取0.25 m2样方,齐地面剪取样方内草样,测定围笼内外的生物量[19]。保留草样用于营养测定。
牧草生长速率:利用围笼内地上生物量月际差额估算草地生长速率。
欧拉羊体重:在牧草生长季(5~10月)按不同年龄家畜比例的15%进行抽样称重,逐月测定不同年龄段家畜平均体重。家畜称重均在清晨空腹进行,称重仪器为上海友声公司生产的家畜称重仪。
营养价值:笼外放牧草地利用手拔法模拟家畜采食,获得草样用于消化率测定。同围笼内草样一起103℃杀青5 min,75℃烘6~7 h至恒重(2次称重差额小于1 g)。利用概略养分分析法测定干物质和粗蛋白含量[19]。
干物质消化率:利用纤维袋法(insacco)在瘤胃瘘管羊体内测定降解率。试验羊为德国美利奴♂×临洮土种羊♀杂交的二代健康公羊,平均体重53 kg。测定方法参照文献[20]进行。
气象资料:查找当地气象资料,确定试验地月均温(℃)、降水量(mm/d)和风力(km/h)等指标。
1.4模型构建
应用Stage one草畜平衡模型为框架,以代谢能为基础,衡量月际间草畜平衡的变化。模型模拟的基础是畜群的平均水平,因此,家畜参数为畜群平均值(如母羊,羯羊,羔羊等的平均体重)。输入家庭牧场的相关参数后,计算家畜采食量和代谢能摄入量,与相应的日维持代谢能需要量(MJ/d)进行比较,计算能量平衡状况(盈余或亏缺),并以此计算家畜日体重(g/d)变化。该模型可以利用前一月家畜的体重和地上生物量估算下一月家畜体重。同时,模型默认家畜的实际体重和估测体重是相互独立的,可为模型校正和参数调整提供方便。
1.5模型算法更新及比较
Stage one模型是一个静态模型,一些变量需要手动输入,以此来模拟家庭牧场的实际情况[16]。模型主体算法主要参考澳大利亚2007年反刍动物营养标准[15]及其配套模型中动物营养计算部分,并做了适当简化。
模型算法和家畜能量需求方面的公式亦可参照文献[13,16,21-23]。
1.5.1 采食量及草地干物质供应的计算 在Stage one模型中,草地被分为喜食牧草与不喜食牧草两个功能组,每个功能组分别依据如下公式计算干物质采食量(DMI,kg DM/d):
DMI=PI×PU×RIDMD×RIDMA
式中:PI(kg DM/d)表示最大采食量,即干物质消化率80%以上时的理论采食量,PU表示最大采食剩余量,模型默认家畜优先选择补饲,最大采食量扣除补饲干物质采食量后余下的未被满足的采食量占最大采食量的比例即为PU。由于模拟区域不补饲或者少补饲,因此试验PU默认为1。RIDMD及RIDMA分别表示干物质消化率及草地干物质供应量对于采食量的抑制作用,计算公式分别为:
RIDMD=
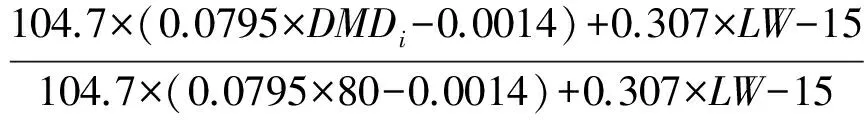
RIDMA=1-e-2Hi
式中:Hi表示放牧草场喜食或不喜食牧草月初生物量(t DM),Hi的计算公式为:
Hi(TDM)=上月牧草留存量+当月生长量-家畜采食量-采食浪费量
式中:采食浪费量默认为生物量的10 %。
根据CSIRO标准[15],不同生理类群欧拉型藏羊的最大采食量计算公式分别为:(1)干奶羊和羯羊最大采食量(PI,kg DM/d)的公式为:
PI(kgDM/d)=0.04×SRW×RS×(1.7-RS)×CF
式中:SRW为标准参考体重,RS(Relative size)是家畜标准体重(NW,kg)和SRW的比值,CF是欧拉羊的体况指数。CF的计算公式为:
CF=RC×(1.5-RC)/0.5
式中:RC是欧拉羊活重(LW,kg)与标准体重(NW)的比值。如果RC>1.0,说明家畜膘情好,利用上式计算CF,反之,CF=1。
在Stage one模型中,开发者对干奶羊和羯羊最大采食量的公式做了相应简化,默认RS=1(即成年欧拉羊),CF=1(表明欧拉羊膘情均处于中下水平)。因此,对于成年家畜最大采食量的简化公式为:
PI=0.04×SRW×0.7
(2)泌乳羊最大采食量的计算公式为:
泌乳羊最大采食量=干奶羊最大采食量×泌乳系数
泌乳系数(m)是用于计算泌乳家畜最大采食量的系数。在Stage one模型中,其计算公式为:
m=1+0.66M1.4exp(1.4(1-M))L
式中:M=T/28,T表示泌乳天数(出生为第1 d,以此类推),28表示最大采食量的泌乳天数;L=0.5+0.5×RCb,其中RCb为产羔时母畜的RC值。在该模型中,默认母羊产羔时RC=1,即膘情指数为中下。
(3)羔羊的最大采食量公式为:
PI=0.04×SRW×RS×(1.7-RS)×CF
式中:CF=1;RS=NW/SRW,NW的计算公式:
NW(kg)=SRW-(SRW-BW)×exp(-0.0157×T×SRW-0.27)
式中:T是幼畜年龄(d),BW是初生重(5 kg)。
1.5.2 代谢能需求的计算 模型能量部分的计算主要分为代谢能摄入和代谢能需求两部分。根据喜食牧草和不喜食牧草两功能组的采食量、消化率,分别计算出两功能组各自的代谢能摄入量,相加即为家畜代谢能摄入量。摄入代谢能要扣除维持需要量、妊娠需要量(仅母羊)、泌乳需要量(仅母羊),剩余部分(代谢能盈余或亏缺)用于计算活体重的变化。
(1)代谢能浓度及代谢能输入量的计算 在CSIRO标准下,饲草和精饲料代谢能浓度的计算公式相同。由于精饲料中粗脂肪含量较一般饲草高,旧版公式完全没有考虑这一因素,会有误差。因此Freer等[24]学者提出了代谢能浓度的修正公式为:
式中:M/D表示代谢能浓度(MJ/kg DM),DMDf、DMDs、DMDh分别代表饲草、精饲料和放牧草地的消化率(%)。EEs表示精饲料的粗脂肪含量(%)。
放牧状态下代谢能摄入量(MEI)的计算公式为:
MEI(MI/d)=DMI×(0.17×DMDh-2)
式中:DMI为干物质采食量(kg DM/d)。
(2)维持代谢能需要量的计算 在放牧状态下维持代谢能的计算公式为:
MEm=MEbase+MEgraze+MEcold[15]
式中:MEbase为家畜在静止且无温度应激状态下,无额外生产消耗时,代谢能的维持需要量,MEgraze为家畜的放牧代谢能消耗,MEcold是气温低于热中性下限温度时额外消耗的能量。前人研究中[13,21-23],仅仅将MEbase当作MEm,而未将MEgraze和MEcold纳入维持代谢范围。按照澳大利亚反刍动物营养标准[15],在放牧状态下将MEgraze和MEcold纳入维持代谢范围内。计算公式为:
MEm(MJ/d)=K×S×M[0.26W0.75exp(-0.03×A)]/km+0.09×MEI+MEgraze+MEcold
式中:K=1.0;母羊或羯羊的S=1.0,公羊的S=1.15;M=1+(0.23×母乳供能/日能量摄入)。
由于放牧状态下确定仔畜的母乳消耗量十分困难,因此Stage one模型中,M统一取值1。A为家畜年龄,在Stage one模型中畜群结构划分相对简单,成年家畜的A值统一简化为4,即将所有母羊和羯羊的年龄确定为4龄,羔羊则按照产羔时间进行换算。
MEbase(MJ/d)=K×S×M×[0.26W0.75exp(-0.03×A)]/km+0.09×MEI
km=(0.02M/D+0.05)×PS+0.85×(1-PS)
式中:km为代谢能用于维持的效率,M/D为代谢能浓度(MJ/kg),PS为家畜日粮中非母乳成分所占比例,在Stage one中PS默认为1。
MEgraze(MJ/d)=[0.02×DMI×(0.9-DMDh/100)+0.0026×H]×LW/km
式中:DMI为家畜采食量(kg/d);DMDh为放牧草地的消化率(%);LW为家畜平均活重(kg);km为代谢能用于维持的效率;H表示家畜运动水平的折算距离(km),其计算公式为:
式中:T根据地势变化(平地到坡地),数值在1到2范围内波动,Stageone模型中,为了计算方便,T值全部简化为1;SR为放牧率(头/hm2)或单位面积的羊只数量;GF为单位面积草地的牧草干物质量(tDM/hm2)。
MEcold(MJ/d)=max(0,0.09×LW0.66×(Tlc-Tair)/(lt+le))
式中:LW为家畜平均活体重(kg);Tair为月平均气温(℃);It为组织绝热系数(Tissueinsulation)(℃m2d/MJ),数值为1.3;Ie为外部绝热系数(Externalinsulation)(℃m2d/MJ);Tlc为热中性下限温度(Lowerlimitationofthermoneutraltemperature)(℃)。It+Ie用如下公式计算:
It+Ie=It+(0.7+0.3×(1-e0.25×R/F))×le
式中:R为平均降水量(mm/d),F为家畜毛被厚度(mm)。
Ie=[r/r+F][1/(0.481+0.326 v0.5)]+rln[(r+F)/r](0.141-0.017 v0.5)
式中:r为家畜体半径,用如下公式计算:
r(mm)=41×LW0.33
Tlc(℃)=39-HP×lt-(HP-1.3)×le
式中:HP是家畜单位面积产热量(MJ/m2),计算公式如下为:
HP(MJ/m2)=
式中:MEI为家畜代谢能日摄入量(MJ/d),Wc为胚胎重量(kg),计算公式为:
Wc(kg)=(Wbirth/4)×e5.17-9.39×exp(-0.006 43×DOC)
式中:DOC指怀孕天数(第1d为1,以此类推)。
(3)妊娠代谢能需要量的计算 妊娠代谢能需求(MEc)计算公式为:
式中:DOC指怀孕天数;Wbirth为初生重(kg),在Stageone模型中统一定为5;0.133为代谢能用于妊娠的效率系数。
(4)泌乳代谢能需要量的计算 泌乳能量的公式参照文献[25],泌乳代谢能(ME1)为家畜最大产奶代谢能需要量(MElmax)和幼畜母乳消耗代谢能(MEl1)中的较小值。计算公式为:
式中:BCbirth为母畜产仔后的相对体况指数(Relative body condition),是LW和SRW的比值,在Stage one模型中统一设定为1;Tl为泌乳时间(月);LWl为羔羊活体重(kg);kl为代谢能用于泌乳的系数,计算公式为:
Tl=DOL+/122
kl=0.4+0.02×M/D
DOL(d)为泌乳天数,在Stage one模型中计算公式为:
DOL(d)=泌乳月份×30-15
其中泌乳月份的数值为0~6。
(5)代谢能平衡及活体重变化的计算 摄入的代谢能扣除以上所有消耗后,可以求出家畜特定月份的平均能量平衡状况(MEbalance=MEintake-MEm-MEc-MEl),正平衡则表明有剩余能量,家畜可以用于增重;如果是负平衡,则表明需要家畜消耗体重,以维持以上能量消耗。家畜活体重变化(LWC)则由以下公式计算:
LWC(g/d)=1 000×MEbalance×kg/EVG
式中:kg为代谢能用于增重的效率,计算公式为:
kg=0.043×M/D
EVG(Empty Body Gain)为增加单位空腹体重需要的能量(MJ/kg),对于成年家畜而言,计算公式为:
EVG(MJ/kg)=0.92×(13.2+13.8×RC)
式中:RC为活体重与标准体重的比值。幼畜的EVG计算公式如下:
EVG(MJ/kg)=
式中:FL为家畜饲养水平,计算公式为:
式中:RS为NW和SRW的比值。
1.6欧拉型藏羊冷季暖棚的模拟
根据郑阳等[21]的研究结果,冬春季暖棚内外的温度差夜间为5℃,白天为10℃。试验假设用暖棚后,日平均温度将提高5℃,以比较冷季条件下暖棚与自然放牧对欧拉型藏羊体况和营养平衡的影响。
1.7欧拉型藏羊氮代谢的模拟
参考Zhou等[26]的结果,以0.5kg/W0.75作为藏羊每日氮维持需要量。研究以测定的粗蛋白含量为对照,建立氮素月份间平衡模型。
2 结果与分析
2.1典型家庭牧场放牧条件下藏羊草畜能量平衡的月际变化
2.1.1 所有家畜的代谢能摄入量、维持代谢能需要量和总代谢能需要量的月际变化 以羊单位(SE,50kg)为标准,典型家庭牧场单位羊只的代谢能摄入量呈钟形曲线,其中1月最低,为4.46MJ/(头·d),7月最高,为21.48MJ/(头·d),7月以后代谢能摄入量迅速下降。与代谢能摄入量相反,单位羊只的维持代谢能需要量整体呈倒钟形,2月最高,为16.08MJ/(头·d),3~7月逐渐下降,7月最低,为9.64MJ/(头·d),之后开始缓慢增加(图1)。其中6~9月家畜代谢能摄入量超过维持需要,10月到翌年5月,代谢能摄入量无法满足维持需要。总代谢能需要量(MEm、MEc、MEl三者之和)表现为冬春高,夏秋低,其中2月最高,为21.94 MJ/(头·d),7月最低,为9.64 MJ/(头·d)。

图1 典型家庭牧场所有家畜的代谢能摄入量、维持代谢能需要量和总代谢能需要量(折算成单位家畜)Fig.1 Actual metabolisable energy intake,maintenance metabolisable energy requirements and total metabolisable energy requirement of all the livestocks on the typical farm (averaged as SE)
2.1.2 母羊代谢能摄入量、维持代谢能需要量和总代谢能需要量的月际变化 母羊的代谢能摄入量呈较平缓的钟形曲线(图2),1月最低,4.89 MJ/(头·d),6月最高,为21.26 MJ/(头·d);而维持代谢能需要量以6月最低,为10.14 MJ/(头·d),8月最高,为16.57 MJ/(头·d);总代谢能需要量以3月最高,达到27.76 MJ/(头·d),7月最低,为10.47 MJ/(头·d)。可知,6~9月母羊的代谢能摄入量超过维持代谢能需要量,10月到翌年5月,代谢能摄入量无法满足需要(图2)。
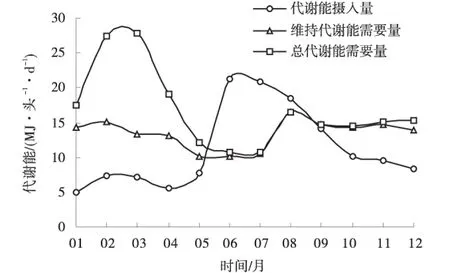
图2 典型家庭牧场所有母羊的代谢能摄入量、维持代谢能需要量和总代谢能需要量Fig.2 Actual metabolisable energy intake,maintenance metabolisable energy requirements and total metabolisable energy requirement of all ewes on typical farmer
2.1.3 放牧率、草地现存量及母羊体重的月际变化 夏秋季草场(7~10月)的放牧率较高,其中9月最高,为3.95 头/hm2;而冬春季草场(11月至翌年6月)的放牧率较低,最低值出现在2月,为2.29 头/hm2(图3)。草地现存量,冬季草场11月到翌年5月呈现逐渐降低趋势,5月之后开始增加;暖季草场先增高后降低,10月最低。母羊体质量4~9月逐渐增加,9月最高(61 kg),10~翌年4月逐渐降低,体质量最低值出现在4月,为51 kg(图4)。
2.1.4 怀孕母羊、干奶羊和羯羊采食量的月际变化 除怀孕母羊外,家畜日干物质采食量(每头)均表现出夏季>秋季>冬春季的特点(图4)。怀孕母羊在泌乳期(2~4月)日干物质采食量较干奶羊分别提高53%,56%和24%。

图3 典型家庭牧场的放牧率、草地现存量和母羊体质量Fig.3 Grazing intensity, herbage mass and ewe liveweight on typical farmer
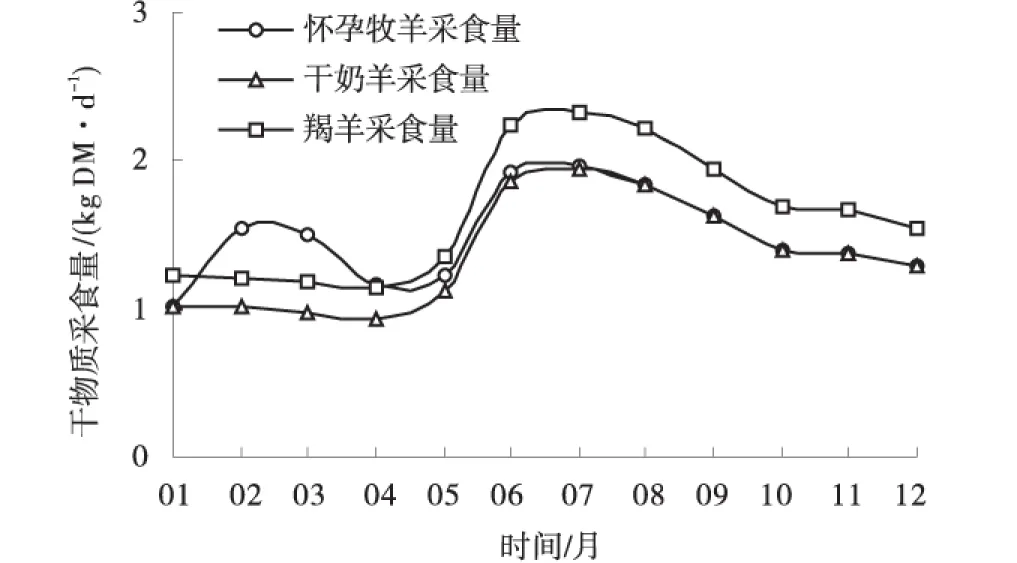
图4 典型家庭牧场不同畜群的平均日干物质采食量Fig.4 Daily averaged dry matter intake of different classes of animals on typical farmer
2.1.5 消化率及能量平衡的分析 此次研究利用模型模拟得到的家畜选择性采食所获得的干物质消化率和实际测定的牧草干物质消化率基本吻合,其中6月的消化率最高,喜食牧草的消化率达到78%,选择性采食后获得的消化率达76.98%;消化率最低值都出现在1~4月,均为40%。模型6~9月代谢能摄入量大于需求量(维持+怀孕+泌乳),代谢能盈余可用于家畜增重(图1~3)。但在5月母羊的能量模拟与实测体质量不符(图2,3),模型模拟代谢能摄入不足,而实际母羊表现出增重。可从二方面分析:1)DMD测定存在误差,由于手拔法无法精确模拟放牧绵羊的选择采食性(5月中下旬草层下层部分返青,藏羊选择性采食了嫩绿的饲草,消化率高,代谢能浓度高),造成消化率测定结果偏低。M/D数值的高低,主要取决于DMD,即准确测定藏羊选择性采食后的牧草消化率是确定采食代谢能浓度M/D的关键。实际状态下,由于选测性采食,消化率会比人为测定的高。所以模型模拟的结果属于保守预测;(2) 家畜称重存在误差,5月是藏羊体况恶劣期,为了保护藏羊,称重样本筛选可能会人为选择体况较好、体重较高的母畜。总代谢能需要量冷季高暖季低,而8月维持代谢能突然提高的原因,是因为剪毛后毛被厚度为最低,使得Ie降低,进而提高了Tlc,使得MEcold有小幅度增加,但摄入代谢能仍然高于维持代谢能(图1,2)。
2.2冷季暖棚试验的模拟
冷季模拟使用暖棚后,1~4月的日维持代谢能需求均减小,MEm分别减少1.08、1.23、1.28和1.26 MJ/(头·d)(表2)。4个月怀孕母羊、干奶羊和羯羊掉膘量分别减少1.43,1.44 和1.46 kg/头,羔羊无明显效果(表3)。
2.3藏羊氮平衡的模拟
欧拉型藏羊摄入氮基本呈正态分布, 4~11月摄入氮能够满足维持需要,12月~翌年3月则呈负平衡状态(图5)。暖季和冷季摄入氮差异较大,暖季每个羊单位平均可摄入39.6 g,冷季则降至最低(5.9 g),说明冬春季是藏羊氮缺乏的关键时期。

图5 典型牧户草场上所有家畜的氮摄入和维持需要(折算成单位家畜)Fig.5 Nitrogen intake and maintenance requirements of all the livestocks on the typical farm (averaged as SE)
3 讨论
3.1冷季草畜平衡研究重点的探讨及补饲饲料的选择

表2 暖棚对家畜日维持代谢能的影响
注:暖棚减少的维持代谢能消耗量=放牧消耗-暖棚消耗
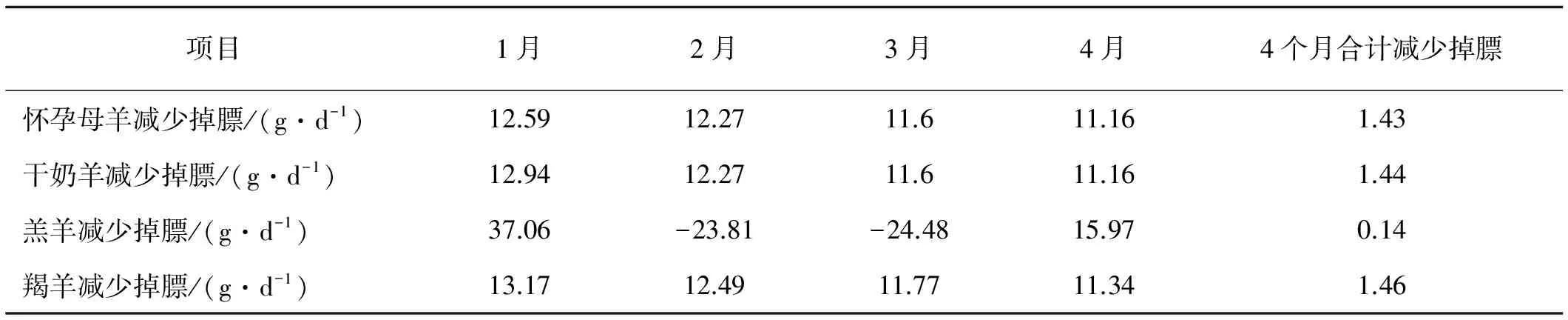
表3 暖棚模拟试验不同级别家畜体质量的变化
家畜需要和可利用饲料供给是高效畜牧业生产的关键[27]。目前我国草畜平衡研究仍然注重载畜量和放牧率研究[23]。高寒草甸适宜载畜量和放牧率研究较多,苏大学[28]认为,高寒草地载畜量很低,高寒草甸类草地的载畜率为1.19绵羊单位/hm2。高寒草甸的理论承载力为0.91羊单位/hm2[29]。Dong等[30]利用Jones和Sandland的模型提出高寒草地暖季草场牦牛适宜放牧率为1.67头/hm2,冷季牦牛适宜放牧率为0.72头/hm2,全年牦牛适宜放牧率为0.63头/hm2。在青藏高原长达7个月的冷季时间内,如何实现草畜平衡是目前研究的热点,但草畜平衡问题若仅从干物质供需平衡的角度考虑是不全面的,家畜10月转场后,现存量在11月出现高峰,放牧率下降明显,但家畜仍处于掉膘状态(图3)。冷季牧草现存量低仅是草畜不平衡问题的一方面,另一方面则是能量浓度较低。放牧状态下除6~9月草地的M/D水平>8 MJ/kg DM外,其余月份的M/D水平均较低(5~8 MJ/kg DM)。暖季草场面积小,但现存量高,能量浓度也高,因此家畜体重大,采食量大,放牧率高,是藏羊每年重要的增重期。青藏高原冷季草地能够提供的干物质及代谢能均为一年中的较低水平。但冷季的代谢能供需严重不平衡仍是冷季草畜平衡评价的重点。Sun等[31]研究发现无论放牧率水平达到或者小于理论载畜量,家畜在冬春草场均会减重,并推论相对于放牧率,冬春季低温可能对家畜生产的影响更大。试验中也可得出相同结论,但从模型角度出发,该现象的原因可用“摄入低、需求高”概括。试验模拟成年羊的最大采食量为2.1 kg(SRW以75 kg计算),可视为干物质采食量的上限,由于M/D水平低,最大采食量获得的代谢能也无法满足家畜的需要,因此表现减重,即冬春季藏羊的代谢能摄入低。青藏高原冷季平均温度远低于CSIRO关于成年绵羊维持代谢状态热中性下限的标准(-3℃),由模型运行结果可知,1~5月由于MEcold的产生,藏羊维持需要量增高;母羊因为妊娠,总代谢能需要量也提高,产羔后泌乳初期(3月)能量需求也较高,即冬春季藏羊的代谢能需求高。季节性草畜不平衡问题,其实就是低饲养水平的营养调控问题[32]。如何提高冬春季饲草饲料的消化率和降低冬春季由于低温造成的维持代谢能的增加,是解决冬春季能量缺乏的关键。大多数发展中国家,尤其是草地生产或者混合生产体系下的反刍家畜大多要经受全年或季节性营养不足。营养缺乏是限制小农户家畜生产的重要因素[33]。现行的反刍动物营养标准有代谢能体系和净能体系,而对于放牧家畜而言,代谢能体系则相对准确。已有不少学者对利用放牧率或载畜量来描述草地供需平衡的科学性提出质疑,并提出了放牧反应指数(Grazing response index)、放牧压力指数(Grazing pressure index)[42]和立枯物生物量(Standing herbage mass)等术语。笔者认为,从生产角度出发放牧率和载畜量不应该作为衡量草畜平衡的关键,也不能作为草地群落健康演替的唯一单纯指标。在放牧系统下,家畜冷季掉膘是不可避免的,管理者需要对短期营养供需不平衡做出考量,并从生产和经济角度做出补饲与否的决策[34]。本研究表明在传统生产模式下,藏羊在冷季均缺乏能量,春季较缺氮,且能量缺口较为明显,营养管理的目标是提高能量浓度即饲草的消化率。从生物学角度讲,冷季补饲能量饲料生物学效率较高,但是精料补饲藏羊的经济效益分析仍需后续探讨。
3.2青藏高原放牧家畜营养研究进展
由暖棚模拟结果(表2)看出,冬春季暖棚的使用,可一定程度上降低家畜冷季维持需要量,但与郑阳等[21]的结果相比,试验模拟节约的家畜掉膘量较少,除去羊品种差异,可能说明模型增重部分的参数需要进一步测定和校正。校正工作需要营养研究来支持,而目前有关青藏高原家畜营养代谢的研究仍然不全面,尤其在藏羊方面,对于维持代谢能至今无正式研究报道。而有关牦牛营养方面的研究则较多。Ding等[35]和Han等[36]研究了成年牦牛在放牧和舍饲状态下维持代谢能的需要量,并指出放牧牦牛每公斤代谢体重的维持代谢能需要高于放牧肉牛。Shao等[37]报道,高寒草地的动物肾脏可能存在某种特殊的肾小球滤过率和嘌呤衍生物调控机制,能够帮助他们更好地应对高原环境。Zhou等[38-39]研究表明,藏羊在低氮日粮下的氮利用率比高山细毛羊高,但是高氮日粮下利用效率则低于高山细毛羊。藏羊因肾脏较高的尿氮重吸收速率,存在高效利用低氮日粮的机制,且每公斤藏羊代谢体重的氮维持需要量仅为NRC标准的66%[26]。牦牛,藏羊蛋白质在妊娠、泌乳和增重方面的利用效率还未见报道。且以上研究,试验动物均为成年动物,不同氮素水平日粮的代谢能浓度相同,有关藏羔羊和牦牛犊牛方面的营养调控研究,藏羊,牦牛能、氮平衡的研究,仍需要进一步探索。已有研究表明夏季舍饲补充精饲料能显著改善舍饲牦牛的生长性能,即牦牛暖季可能存在能量摄入不足,限制其高效增重问题[40]。而藏羊也可能存在相同的问题。由模型可知,暖季藏羊氮摄入量远高于其维持需要量,但青藏高原和大多数陆地生态系统一样,是氮素限制性生态系统,模型结果也不能完全证明暖季藏羊不缺氮。对于不同能、氮水平下,藏羊的生长潜力仍然需要实验研究予以确认,而这些对于该地区生产效率水平的分析和可持续草地畜牧业的发展无疑是至关重要。
3.3青藏高原放牧家畜采食的研究和模拟
放牧条件下,确定家畜日粮采食的组份及采食量是放牧家畜营养研究的难点[34]也是放牧采食量模型模拟的挑战[41]。自由放牧下,影响反刍动物采食量的因素有很多。采食量的控制也是多因子。在前人大量的工作基础上,不少学者已经对影响和调控采食量的因素做了相关综述。现行的自由采食量测定方法较多,各方法都有优缺点。对于藏羊自由采食量的报道仍然较少。将模型模拟的羯羊采食量变化同淡瑞芳等[42]的研究对比,线性回归R2值为0.841。而母羊的模拟结果和吴铁成等[43]的研究进行比较,模拟结果普遍偏高。淡瑞芳的方法是差额法,Undi等[44]在放牧肉牛的研究中提出,差额法测得的干物质采食量较其他方法偏高。因此,该方法不适宜测定个体家畜的采食量,适用于测定畜群的采食量。吴铁城等采用的外援指示剂法,其缺陷在于指示剂在粪中的回收率无法达到100%的理想值,且粪中指示剂浓度日间的变化较大。因此推荐自由采食量的测定要用多种方法互相比较[45]。采食量也可通过模型模拟进行研究,CSIRO标准有专门开发的采食量预测模型[41],CSIRO标准也因此在模拟放牧家畜营养代谢方面有其他主流营养标准无法比拟的优势。但由于其采食量模拟的复杂性,在简单的excel平台无法完全实现。因此在Stage one模型中进行了简化,草地植物被区分为喜食牧草和不喜食牧草。假定家畜优先选择补饲饲料(消化率大于等于80%),然后放牧,同时Stage one模型中不再详细划分不同的消化率组,而是利用实验测定的消化率数据,代表整个草地平均消化率水平。放牧状态下采食量的模型模拟仍将是未来的研究热点,CSIRO标准的采食量模型如何应用到草原区,仍需要进一步的研究。
3.4系统模型在研究放牧系统中的作用及展望
农业系统学模型的研究起源于20世纪50年代,最初是用于作物光合作用和产量的模拟[46]。从20世纪60年代C.T.deWit和W.G.Ducan农业系统模型的雏形,到70年代美国政府投资开发CERES-WHEAT和CERES-MAIZE模型,再到2010年AgMIP项目的实施。农业系统模型在近半个多世纪的发展速度相当迅猛。如今农业系统模型已经深入到农业研究的各个领域。并且和生态学和农业经济学等学科发展交叉领域研究,产生了如农业生态学模型和农业经济学模型等众多应用研究领域[46]。由于放牧系统的复杂性,专门化草地模型的出现与作物模型相比稍晚,首个综合性草地模拟模型ELM在上世纪70年代末由美国开发[47]。在过去10多年中,主流作物模型不断发展,众多子模块的开发是为了将作物模型发展成为综合性的农业生产体系模型[48],例如澳大利亚开发,在澳大利亚和新西兰利用广泛的作物模型APSIM就在尝试加入动物模块,发展成为适用于草地农业生产体系的综合模型[49]。有关作物模型和草地放牧模型的综述较多,最新资料可以参考Holzworth[48]和Snow[50]。现今模型的发展趋势不再是创造新的模型公式,而是在已有模型的基础上,不断整合,加工,完善形成新的模型[51]。计算机及互联网技术的快速发展促使农业模型的功能不断强大。纵使模型的功能日益完善,但是模型不是万能的。模型的开发初期一定要明确模型需要解决的问题及模型框架的构建。以Stage one模型为例,其优势在于能够快速模拟或预测年度草畜营养平衡,源代码开放,构建平台简便,用户可以根据自身情况,修改模型内部参数。对于草畜月际代谢能平衡的模拟方便直观。从功能上分析,它是研究模型,即帮助科研工作者了解整个放牧系统的变化模式,而不是应用模型,因为Stage one里并没有插入任何用于决策支持系统(DSS)或优化模拟的插件。现今农业模型的研究越来越重视气候变化下农业生产体系的生产效率和对环境的影响。而在简单的excel平台下,引入随机变量较为困难,因此Stage one只能是静态模型,而不能作为动态模型,预测系统多年的变化。同时Stage one模型使用界面不友好,容易出现人为的修改错误,无法较好的划分年龄类群,导致模拟计算都默认为成年家畜,容易过高估计畜群的采食量和能量消耗,模型预测结果较为保守。因此,模型的使用,要求使用者对于每个模型的优缺点有较为深刻的理解。且模型开发者在开发过程中会对模型的复杂性进行控制,因此,即使是现在主流的草地系统模型,也无法详细地模拟瘤胃的反刍过程,采食量的模拟也是相对简单的。目前也存在很多复杂的机械化模型专门针对这些细节性问题[50]。我国草地放牧系统模型的研究仍然处于起步阶段。在Stage one基础上,针对藏羊和牦牛的动态生物经济学模型正在开发之中。模型开发及后续应用需要不断地校正和修改,而有关基础数据的收集,尤其是草畜互作界面的数据,仍然需要大量开展相关研究。
4 结论
试验利用澳大利亚反刍动物营养标准,在青藏高原东缘高寒草甸区,建立了一个藏羊的静态、月际能、氮平衡模型。结果表明,藏羊在暖季(6~9月)牧草消化率高,草地能量浓度高,能够获得较多的代谢能用于体重增加。而在冷季,由于气温较低,维持代谢能需要量提高。春季是母羊怀孕末期能量需求的旺季,因此冷季尤其是春季藏羊的能量需求最高,补饲应以能量饲料为主。暖棚可减少藏羊冷季维持代谢能需要量,并一定程度上降低掉膘量。藏羊仅在春季表现为氮素维持负平衡,其余季节氮素摄入均在维持水平以上。因此高寒草甸区冷季精饲料补饲+暖棚节约御寒代谢能需要是较为可行的管理方式。从生产角度看,代谢能平衡是草畜平衡的核心,放牧率并不是生产角度下评价草畜平衡的最佳指标。CSIRO标准的采食量模型较为复杂,不适用于简单的模型中,且CSIRO标准模拟藏羊增重方面仍然存在修改和提升的空间。
[1] FAO.Building a common vision for sustainable food and agriculture:Principles and approaches[R].Rome:Food and Agriculture Organization,2014.
[2] Phalan B,Green R,Dicks L,etal.How can higher-yield farming help to spare nature[J].Science,2016,351(6272):450-451.
[3] Herrero M,Thornton P K.Livestock and global change:Emerging issues for sustainable food systems[J].Proceedings of the National Academy of Sciences of the United States of America,2013,110(52):20878.
[4] García-Martínez A,Rivas-Rangel J,Rangel-Quintos J,etal.A methodological approach to evaluate livestock innovations on small-scale farms in developing countries[J].Future Internet,2016,8(2):25.
[5] Vignola R,Harvey C A,Bautista-Solis P,etal.Ecosystem-based adaptation for smallholder farmers:Definitions,opportunities and constraints[J].Agriculture,Ecosystems & Environment,2015,211:126-132.
[6] Altieri M A,Funes-Monzote F R,Petersen P.Agroecologically efficient agricultural systems for smallholder farmers:Contributions to food sovereignty[J].Agronomy for Sustainable Development,2012,32(1):1-13.
[7] Himmelstein J,Ares A,van Houweling E.Sustainable intensification:A multifaceted,systemic approach to international development[J].Journal of the science of food and agriculture,2016,96(15):4833-4839.
[8] Shang Z H,Gibb M J,Leiber F,etal.The sustainable development of grassland-livestock systems on the Tibetan plateau:Problems,strategies and prospects[J].Rangeland Journal,2014,36(3):267-296.
[9] 付伟,赵俊权,杜国祯.青藏高原高寒草地放牧生态系统可持续发展研究[J].草原与草坪,2013,33(1):84-88.
[10] 钱拴,毛留喜,侯英雨,等.青藏高原载畜能力及草畜平衡状况研究[J].自然资源学报,2007,22(3):389-397+498.
[11] 刘兴元,冯琦胜,梁天刚,等.甘南牧区草地生产力与载畜量时空动态平衡研究[J].中国草地学报,2010,32(1):99-106.
[12] 梁天刚,冯琦胜,夏文韬,等.甘南牧区草畜平衡优化方案与管理决策[J].生态学报,2011,31(4):1111-1123.
[13] 杨博,吴建平,杨联,等.中国北方草原草畜代谢能平衡分析与对策研究[J].草业学报,2012,21(2):187-195.
[14] 郭雅婧,薛冉,李春涛,等.青藏高原草畜供求的月际平衡模式研究[J].中国草地学报,2014,36(2):29-35.
[15] Primary Industries Standing Committee.Nutrient requir ements of domesticated ruminants[M].Melbourne:CSIRO Publishing,2007.
[16] Takahashi T,Jones R,Kemp D.Steady-state modelling for better understanding of current livestock production systems and for exploring optimal short-term strategies[A]∥.Kemp D,Michalk D.Development of Sustainable Livestock Systems on Grasslands in North-western China.Hohhot:Australian Centre for International Agricultural Research (ACIAR),2011:26-35.
[17] Li Q,Yang X,Soininen J,etal.Relative importance of spatial processes and environmental factors in shaping alpine meadow communities[J].Journal of Plant Ecology,2011,4(4):249-258.
[18] 王宏博,丁学智,郎侠,等.甘南玛曲夏季牧场欧拉型藏羊牧食行为的研究[J].草地学报,2012,20(3):583-588.
[19] Liu H B,Wu J P,Tian X H,etal.Dynamic of abo veground biomass and soil moisture as affected by short-term grazing exclusion on eastern alpine meadow of Qinghai-Tibet plateau,China[J].Chilean Journal of Agricultural Research,2016,76(3):321-329.
[20] 梁建勇,焦婷,吴建平,等.不同类型草地牧草消化率季节动态与营养品质的关系研究[J].草业学报,2015,24(6):108-115.
[21] 郑阳,徐柱,郝峰,等.内蒙古典型草原家庭牧场暖棚舍饲养羊效益分析[J].家畜生态学报,2010,31(2):64-69.
[22] Li Z G,Han G D,Zhao M L,etal.Identifying management strategies to improve sustainability and household income for herders on the desert steppe in inner mongolia,China[J].Agricultural Systems,2015,132:62-72.
[23] 李江文,李治国,王静,等.草畜能量平衡模型在内蒙古草甸草原上的应用[J].中国草地学报,2016,38(2):79-85.
[24] Freer M,Moore A D,Donnelly J R.The GRAZPLAN animal biology model for sheep and cattle and the GrazFeed decision support tool[EB/OL].http://www.grazplan.csiro.au/files/TechPaperMay12.pdf,2012-12.
[25] Freer M,Moore A D,Donnelly J R.GRAZPLAN:Decision support systems for australian grazing enterprises-ii.The animal biology model for feed intake,production and reproduction and the GrazFeed dss[J].Agricultural Systems,1997,54(1):77-126.
[26] Zhou J W,Guo X S,Degen A A,etal.Urea kinetics and nitrogen balance and requirements for maintenance in tibetan sheep when fed oat hay[J].Small Ruminant Research,2015,129:60-68.
[27] Kemp D,Brown C,Han G D,etal.Chinese grasslands:Problems,dilemmas and finding solutions[C]∥Kemp D,Michalk D.Development of Sustainable Livestock Systems on Grasslands in North-western China.Hohhot:Australian Centre for International Agricultural Research (ACIAR),2011:12-23.
[28] 苏大学.中国草地资源的区域分布与生产力结构[J].草地学报,1994,2(1):71-77.
[29] 中华人民共和国农业部畜牧兽医司.中国草地资源数据[M].北京:中国农业科技出版社,1994.
[30] Dong Q M,Zhao X Q,Wu G L,etal.Optimization yak grazing stocking rate in an alpine grassland of Qinghai-Tibetan plateau,China[J].Environmental Earth Sciences,2015,73(5):2497-2503.
[31] Sun Y,Angerer J P,Hou F J.Effects of grazing systems on herbage mass and liveweight gain of Tibetan sheep in eastern Qinghai-Tibetan plateau,China[J].The Rangeland Journal,2015,37(2):181-190.
[32] 郝正里,刘世民,孟宪政.反刍动物营养学[M].兰州:甘肃民族出版社,2000.
[33] Thornton P K.Livestock production:Recent trends,future prospects[J].Philosophical Transactions of the Royal Society of London:Biological Sciences,2010,365(1554):2853-2867.
[34] Dove H.Balancing nutrient supply and nutrient requirements in grazing sheep[J].Small Ruminant Research,2010,92(1):36-40.
[35] Ding L M,Wang Y P,Brosh A,etal.Seasonal heat production and energy balance of grazing yaks on the Qinghai-Tibetan plateau[J].Animal Feed Science and Technology,2014,198:83-93.
[36] Han X T,Xie A Y,Bi X C,etal.Effects of high altitude and season on fasting heat production in the yak Bos grunniens or Poephagus grunniens[J].British Journal of Nutrition,2002,88(2):189-197.
[37] Shao B,Long R J,Ding Y,etal.Morphological adaptations of yak (Bos grunniens) tongue to the foraging environment of the Qinghai-Tibetan plateau[J].Journal of Animal Science,2010,88(8):2594-2603.
[38] Zhou J W,Mi J D,Degen A A,etal.Apparent digestibility,rumen fermentation and nitrogen balance in Tibetan and fine-wool sheep offered forage-concentrate diets differing in nitrogen concentration[J].Journal of Agricultural Science,2015,153(6):1135-1145.
[39] Zhou J W,Mi J D,Titgemeyer E C,etal.A comparison of nitrogen utilization and urea metabolism between Tibetan and fine-wool sheep[J].Journal of Animal Science,2015,93(6):3006-3017.
[40] 方雷.饲养方式对夏季牦牛生长性能、屠宰性能、牛肉品质和瘤胃细菌多样性的影响[D].北京:中国农业大学,2015.
[41] Pulina G,Avondo M,Molle G,etal.Models for estimating feed intake in small ruminants[J].Revista Brasileira de Zootecnia,2013,42(9):675-690.
[42] 淡瑞芳,龙瑞军,张海涛,等.藏系绵羊瘤胃细菌区系的季节动态分析[J].动物营养学报,2009,21(5):798-802.
[43] 吴铁成,吴建平,赵生国,等.甘南玛曲不同年龄欧拉型藏羊放牧采食量季节动态研究[J].草地学报,2016,24(2):317-321.
[44] Undi M,Wilson C,Ominski K H,etal.Comparison of techniques for estimation of forage dry matter intake by grazing beef cattle[J].Canadian Journal of Animal Science,2008,88(4):693-701.
[45] Mayes R W,Dove H.Measurement of dietary nutrient intake in free-ranging mammalian herbivores[J].Nutrition Research Reviews,2000:107-138.
[46] Jones J W,Antle J M,Basso B,etal.Brief history of agricultural systems modeling[J].Agricultural systems,2016,155:240-254.
[47] Moore A D,Holzworth D P,Herrmann N I,etal.Modelling the manager:Representing rule-based management in farming systems simulation models[J].Environmental Modelling and Software,2014,62:399-410.
[48] Holzworth D P,Snow V,Janssen S,etal.Agricultural production systems modelling and software:Current status and future prospects[J].Environmental Modelling and Software,2015,72:276-286.
[49] Holzworth D P,Huth N I,Zurcher E J,etal.APSIM-evolution towards a new generation of agricultural systems simulation[J].Environmental Modelling and Software,2014,62:327-350.
[50] Snow V O,Rotz C A,Moore A D,etal.The challenges-and some solutions-to process-based modelling of grazed agricultural systems[J].Environmental Modelling & Software,2014,62:420-436.
[51] Kumar S,Shamim M,Bansal M,etal.Computational modeling and emerging trend in agriculture[C]∥Jaimini U et al.International Conference on Computing for Sustainable Global Development.New Delhi:IEEE,2015:1156-1160.
Studyonthegrassland-livestockbalancebasedonstaticmodelinthealpinemeadowontheeasternedgeoftheQinghaiTibetanPlateau
LIU Hai-bo1,LI Lin-yuan2,PU Xiao-jian1,DU Wen-hua1
(1.CollegeofPrataculturalScience,GansuAgriculturalUniversity/KeyLaboratoryofGrasslandEcosystem,MinistryofEducation/PrataculturalEngineeringLaboratoryofGansuProvince/Sino-U.S.CentersforGrazinglandEcosystemSustainability,Lanzhou730070,China;2.TibetanhighschoolinChongqing,Chongqing400036,China)
In order to study the grassland-livestock balance in alpine meadow area on the eastern edge of Qinghai-Tibetan Plateau,a nutrition model was constructed using Australian ruminant feeding standard as reference,stage one model as frame,typical family farm as study scale,and metabolisable energy (ME) and nitrogen requirement as nutrition unit.The nutrition model was used to stimulate the monthly variation of ME and nitrogen requirement of the Oula Tibetan sheep in this area and to study effects of winter warm sheds on weights of the sheep.The results showed that the Tibetan sheep could obtain enough energy for increase of weight under high stocking rate (>3.5 SE/ha) during warm seasons in the alpine meadow with high forage digestibility and ME content.The Tibetan sheep could not obtain enough energy for keeping of weight under low stocking rate (<3.5 SE/ha) during cold seasons in the alpine meadow with low forage digestibility and ME content.The energy requirement of the Oula Tibetan sheep increased greatly and grassland-livestock was not kept in good balance due to the low air temperature during the cold seasons in the alpine meadow.Concentrate feeds should be considered as the priority supplements during cold season.Warm shed could help to reduce the ME expenditure and the ME metabolic status during cold seasons.The ME of the Oula sheep reduced 1.08, 1.23, 1.28 and 1.26 MJ/SE/d from January to April, respectively, after the Oula sheep being kept in warm shed. Concentrate supplement+warm shed would be a practical management during the cold seasons in this area.
alpine meadow;Oula Tibetan sheep;metabolisable energy;grassland-livestock balance
2017-05-05;
:2017-05-25
国家自然科学基金(31360577);现代农业产业技术体系建设专项资金资助(CARS-40-09B)资助
刘海波(1989-),男,甘肃平凉人,博士研究生。 E-mail:710453724@qq.com 杜文华为通讯作者。
S 812
:A
:1009-5500(2017)04-0026-13
