去除优势种后高寒针茅化草甸群落对增温增雨及其协同作用的响应
2017-09-20史雅楠钟梦莹李晶晶索南加尕藏太邵新庆
史雅楠,钟梦莹,李晶晶,索南加,尕藏太,邵新庆
(1.中国农业大学 动物科技学院,北京 100093;2.青海贵南县草原站,青海 贵南 813100)
去除优势种后高寒针茅化草甸群落对增温增雨及其协同作用的响应
史雅楠1,钟梦莹1,李晶晶1,索南加2,尕藏太2,邵新庆1
(1.中国农业大学 动物科技学院,北京 100093;2.青海贵南县草原站,青海 贵南 813100)
2015年6~8月在青海省海北州高寒针茅化草甸草原进行了模拟气候变化及去除优势种紫花针茅(Stipapurpurea)野外控制试验。通过设置增温2℃,增加降雨量20%,研究植物优势种丧失后气候因素对物种多样性和群落稳定性的影响。结果表明:去除紫花针茅后,增温,增雨及两者的交互作用对植物高度、盖度及地上生物量均没有显著影响;增温处理下Shannon-Wiener 多样性指数、Simpson 优势度指数、Pielou 均匀度指数均与对照差异显著(P<0.05),说明增温可以提高植物群落的丰富度和均匀度,增雨处理下3个多样性指数均为最高值;去除优势植物紫花针茅后,植物物种数与植物盖度,植物盖度及优势度指数均呈显著正相关。
优势种去除;增温;增雨;生物量;多样性
物种多样性对于维系生态系统多功能性有着极其重要的作用,一直是生态学领域研究的重点内容[1]。在人类活动干扰和气候变化的影响下,我国北方高寒草原生态系统发生了不同程度的退化,生态环境恶化,第一性生产力降低,草地群落结构单一,物种多样性下降[2-3]。近些年全球气候变化对草地生态系统的影响呈逐渐加强的趋势[4]。IPCC第5次评估报告(AR5)指出,到21世纪末,全球温度增幅可达到2℃以上,中国青藏高原区的降水量将提高10%~20%,这种趋势必对草地生态环境和物种多样性产生深刻的影响。另一方面,超载过牧以及家畜选择性采食,在一定程度上会加剧群落内一些次优势种和伴生种的丧失,导致群落稳定性下降[5]。
青藏高原高寒草甸生态系统因其脆弱的生态环境而对气候变化和放牧干扰的响应极为敏感[6]。近年来,该系统区域气候发生了明显的变化,生长季温度平均每年增加0.071℃,高于全国的0.046℃。同时受到全球降水格局变化的影响,降水量呈现升高的趋势[7]。随着降水格局变化和温度升高,高寒草甸生态系统在初级生产力、群落物种组成以及多样性等方面均产生了改变[8-11]。诸多的学者通过模拟试验,研究了气候变化对植物生理生态[12],土壤理化性质及CN循环生态过程的影响[13-15]。模拟放牧家畜选择性采食研究多用去除试验,研究集中在去除功能群后对群落结构以及稳定性的影响[16-17]。但就全球气候变化及放牧对高寒草甸草原稳定性和多样性的研究仍不够系统和完善,笔者通过设置增温增雨模拟试验,同时去除优势植物紫花针茅模拟放牧家畜选择性采食,以期探讨高寒草甸生态系统植物生产力和物种多样性的变化趋势,为未来高寒草甸生态系统的适应性管理提供基础数据和理论依据。
1 材料和方法
1.1试验地概况
试验地位于青海省海北藏族自治州海晏县海北高原现代生态畜牧业科技示范试验示范园,地处青藏高原东北部青海湖东岸,地理位置E 100°57′,N 36°55′,海拔3 029 m。属高原大陆性季风气候,年均温1.5℃,年均降水量580 mm,降水主要集中在7~9月。土壤类型为高山草甸土,土层较薄,植被类型为高寒针茅化草甸草植被,植被总盖度80%,建群种为紫花针茅(Stipapurpurea),草地早熟禾(Poapratensis),冷地早熟禾(Poacrymophila),披碱草(Elymusdahuaricus),线叶嵩草(Kobresiacapillifolia)等。
1.2试验设计
样地始建于2014年6月,样地内设16个小区,设计增温(W)、增雨(P),增温增雨(WP)及对照(CK)4个处理,每个处理重复4次,采用4×4拉丁方设计。每个小区为直径2.2 m圆形,小区与小区间隔3 m。增温采用圆底式开顶箱增温装置(OTC)增温2℃,并设计了自动控温系统和抽气式风扇,当OTC内外空气温度差值超过2℃时,风扇自动启动,保证增温不超过设定值,维持温度稳定性。OTC永久性安装于样地里,规格是底部直径为2.2 m,顶部直径1.5 m,高度0.7 m。增雨采用人工增加,即每次降雨后在原降雨水平下,用喷壶增加20%的水量。在16个小区内分别设置一个25 cm×25 cm的样方,于2015年6月进行紫花针茅剔除控制试验,将紫花针茅齐地面刈割,每1周剔除1次,同年8月进行样方调查和材料收集。
1.3测定指标
多度测定:将样方框用细线均分为100个小格,严格统计记录每个物种的个数。
株高测定:在样方内每种植株随机选取10株,用钢卷尺测量从地面到植株自然高度的最高点,求平均值。
盖度测定:将样方框用细线均分为100个小格,严格记录样方中植物的盖度。
植物生物量测定:将所有物种齐地面刈割,装入自封袋中,立即带回实验室,称量植物鲜重,记录数据后,在65℃恒温干燥箱干燥48 h至恒重,将干燥后的植物样本称重,得到样方内所有植物种的地上生物量数据。
多样性指数:植被群落多样性指数采用Shannon-Wiener 多样性指数、Simpson 优势度指数、Pielou均匀度指数进行分析,并用公式计算[18]。
重要值=
(1)
Shannon-Wiener 多样性指数
H=-ΣPilnPi
(2)
式中:Pi=Ni/N;Ni其中为样方中第i种植物的重要值;N为样方植物重要值总和。
Simpson 优势度指数
D=1-ΣPi2
(3)
Pielou 均匀度指数
E1=H/lnS
(4)
E2=(1-ΣPi2)/(1-1/S)
(5)
式中:H为Shannon-Wiener 多样性指数;S为植物物种数。
1.4数据处理
采用Excel和Sigma Plot 10.0制作图表,使用SPSS 17.0进行数据处理和统计分析。通过SPSS软件的LSD多重比较的方法分析在不同处理下植物生长及多样性的差异(one-way ANOVA,SPSS 17.0)。使用GLM 双因素分析检验去除紫花针茅,增温、增雨、增温增雨对植物生长等指标的影响(two-way ANOVA,SPSS 17.0)。
2 结果与分析
2.1剔除紫花针茅后不同处理对物种数和地上生物量的影响
地上生物量是草地生态系统基本的数量特征,试验中的地上生物量是在植物生长季达到最高峰时进行的采集和称量,因此,地上生物量实际上是植物的净初级生产力。分析发现,在剔除紫花针茅的前提下,增温,增雨及增温增雨交互处理下植物物种数和地上生物量均无显著影响(P>0.05),处理之间也没有表现出差异(表1)。增温处理和增温增雨交互处理的物种数与对照相比物种多样性减少,分别减少了6.8%和13.6%,增雨处理使物种多样性增加了4.5%,均未达到显著水平(P>0.05),可以看出,只是单纯的剔除一个物种对群落的多样性影响不大。3种处理(W,P和WP)下地上生物量与对照相比都减少了,并且交互作用(WP)下达到最低值,但差异均不显著(P>0.05)。气候变化对植物物种数的影响程度表现为P>CK>W>WP,对地上生物量的影响表现为P>W>WP>CK,在不同处理下生物量与物种数的变化并不一致。增雨处理下物种数和地上生物量都呈增加趋势,但未达到显著水平(P>0.05)。

表1 剔除紫花针茅后不同处理下物种数和地上生物量
注:同列不同小写字母表示差异显著(P<0.05)
2.2剔除紫花针茅后不同处理对群落盖度和高度的影响
在4种处理下剔除紫花针茅后植物盖度和平均高度并没有发生显著变化(图1,2)。气候因素对植物盖度的影响程度表现为P>CK>WP>W,在高度上的则表现为WP>W>P>CK。与对照比,增温使植物盖度降低了13.2%,增温增雨的交互作用导致盖度降低了17.2%,而增雨增加了植物盖度,增加了2.6%,但三者处理都未达到显著水平(P>0.05)。与对照相比,3种处理都使植株高度有所增加,分别增加了22.4%,8.0%和28.4%,但差异不显著(P>0.05)。
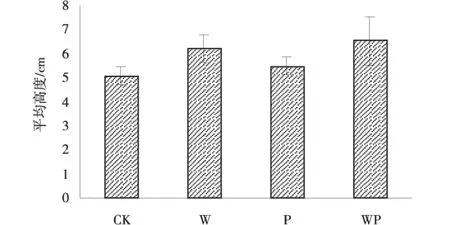
图1 植物高度Fig.1 Plant height

图2 植物盖度Fig.2 Plant coverage
2.3剔除紫花针茅后不同处理对植物多样性的影响
Shannon-Wiener 多样性指数、Simpson 优势度指数、Pielou 均匀度指数显著性对比分析显示(图3),4个多样性指数在不同处理下的变化趋势一致,均表现为P>CK>WP>W。分析Shannon-Wiener 多样性指数,增雨处理显著高于增温处理(P<0.05),其他处理间无显著性差异。在Simpson 优势度指数上,增雨处理与对照和增温处理差异显著(P<0.05),且数值最小。Pielou 均匀度指数E1和E2的变化规律一致,增温处理显著低于其他3个处理(P<0.05)。与对照相比,增温使得群落优势度指数和均匀度指数降低,其他处理(P,WP)无显著影响。
2.4增温和增雨的互作效应分析
通过双因素方差分析可知,增温对植物Simpson指数和Pielou指数有极显著影响作用(P<0.001),对Shannon-Wiener多样性指数有显著影响(P<0.01),但是对植物的生物量、物种数,高度和盖度无显著作用(P>0.05)。增雨处理对植物生产力无显著影响,但是对均匀度指数(E2)有显著提高的作用(P<0.001)。增温和增雨的交互处理对Pielou均匀度指数-E1有显著作用(P<0.01),但对植物盖度、高度、物种数、多样性指数、生态优势指数和均匀度指数E2并无显著影响(表2)。
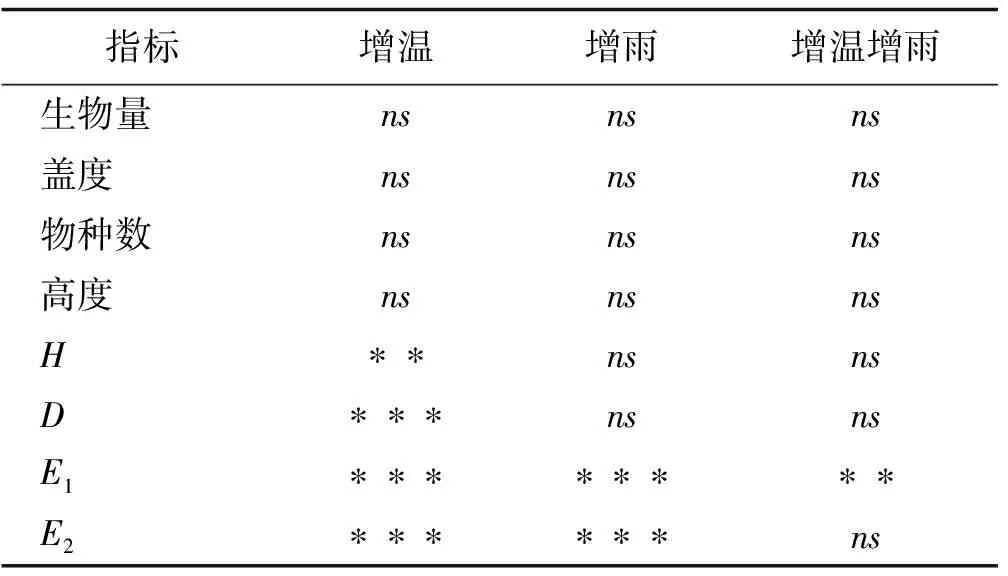
表2 增温和增雨处理对剔除针茅后植物各指标影响的双因素方差分析
注:“*”表示P<0.05,“**”表示P<0.01,“***”表示P<0.001,“ns”表示无显著
2.5相关性分析

图3 针茅化草甸草原去除紫花针茅后植物功能多样性指数±s)Fig.3 The diversity indices and uniformity indices of plants after removal of Stipa purpurea.
对剔除紫花针茅后的处理数据进行Person相关性分析后可知(表3),物种数分别与Shannon-Wiener多样性指数(r=0.88,P<0.01)及Simpson 优势度指数(r=0.74,P<0.01)呈显著正相关;植物盖度与物种数(r=0.77,P<0.01)、Shannon-Wiener 多样性指数(r=0.84,P<0.01)和Simpson 优势度指数(r=0.72,P<0.01)均呈显著正相关;对于植物多样性指数之间的关系而言,Shannon-Wiener 多样性指数与优势度指数(r=0.94,P<0.01)、Pielou 均匀度指数-E1(r=0.53,P<0.05)、Pielou 均匀度指数-E2(r=0.64,P<0.01)呈显著正相关;优势度指数与均与度指数E1(r=0.66,P<0.01)和E2(r=0.83,P<0.01)均呈显著正相关。
植物物种数与Shannon-Wiener 多样性指数(图4a),Simpson 优势度指数(图4b)及植物盖度(图4c)均呈显著正相关(P<0.01)。植物盖度分别与Shannon-Wiener多样性指数(图4d),Simpson 优势度指数(图4e)呈显著正相关(P<0.01)。

表3 剔除紫花针茅后地上植物功能与多样性的相关性
注:“*”表示P<0.05,“**”表示P<0.01,“***”表示P<0.001,“ns”表示无显著
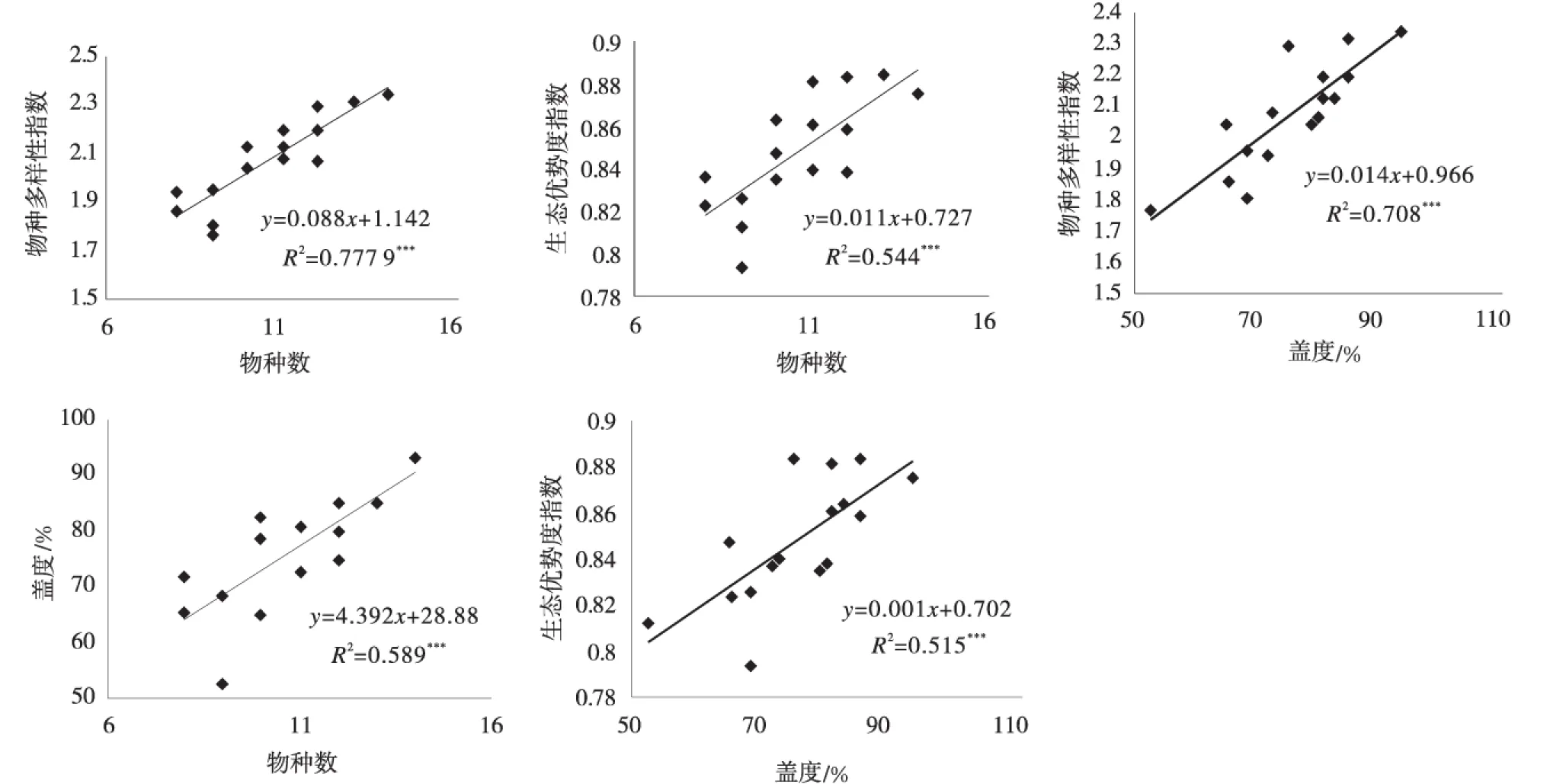
图4 物种数/盖度与物种多样性指数,优势度指数的回归分析Fig.4 Regression analysis between plant species diversity and coverage indices,and between coverage indices and dominance indices
3 讨论
3.1去除优势种对植物群落的影响
草地系统的平衡稳定在很大程度上依赖于群落多样性,而放牧影响着植物多样性。有研究结果表明,家畜采食优势物种可以增加植物多样性[20]或通过采食喜食物种导致植物多样性的减少或无显著影响[20-22]。试验通过去除紫花针茅这一优势物种来模拟家畜对草原上层优势牧草的采食,研究对群落其他物种的影响。研究结果表明,去除优势种后植物群落维持相对稳定,对群落生物量无显著影响,这与杨中领等[23]的研究结果相吻合。优势植物去除后,植物物种数与物种多样性指数及优势度指数均成正相关,这代表短期的物种丧失并不会破坏群落稳定。Wardle等[24]的模拟试验结果显示,物种的去除促进了保留种的生长,功能群间的竞争关系通过补偿生长进而维持稳定。优势种的去除,必然导致群落性质的改变,因此,适当的放牧,可以使植物群落丰富度增加,维持植物群落的稳定,从而有利于提高群落的生产力[25]。
3.2气候变化对植物群落的影响
温度作为影响植物生长的关键因子之一,其变化会影响植物的生长。增温2℃使得植物物种数和植物盖度有下降的趋势,这与Bai[5]在青藏高原草甸的增温试验及Wang等[26]在高寒草原的增温试验结果一致,发现增温使得物种丰富度下降。在增温对生物量影响的研究中,李英年等[27]认为长期增温会导致草地生物量降低,试验结果与之相反,可能是因为优势种的去除减少了对其他物种的竞争作用,而短期增温促进了植物的生长。优势种的去除加上温度的共同作用使得群落多样性也发生了改变,其中生态优势度指数和均匀度指数显著降低(P<0.05),去除优势种使得其他物种获得更多生长资源,分布逐渐均匀。
温度变化常伴随降雨的改变,降水可以改变植物对水分的利用情况,从而影响植物生长。试验结果表明增加降雨量对植物群落并没有显著影响,而王长庭等[28]通过模拟降雨试验研究发现在植物生长期间增加水量可增加群落物种多样性指数和均匀度指数。但也有研究发现增加降雨能使优势种比例增加,从而导致群落物种均匀度下降[29]。研究结果中,增加降雨量使得植物生物量和盖度有少量的增加,但并不明显,而植物多样性指数和优势度及均匀度都没有过大的改变。这可能与生长季内降雨量的浮动或土壤类型、团聚体结构不同有关系,这些因素影响植物对水分的利用,从而导致研究结果不一致。
关于温度和水分的交互作用,有研究发现在限制性增温和增雨条件下,可以增加植物生物量和平均高度[30]。在内蒙古克氏针茅草原上增温增雨试验增加了植物群落的高度,但对群落的物种多样性和均匀度指数均没有显著影响[14]。在此次试验中,双因素方差分析得出增温增雨的交互作用只对植物生物量和物种均匀度有显著影响,在两者交互作用下植物生物量和高度有所增加,此结论与试验结果相符。而试验结果效果不显著与去除优势种试验只进行2个月有关,需要今后试验进一步探讨。试验表明适当增加温度和降雨有助于植物生长和群落的温度。
3.3综合效应
在青藏高原半干旱地区,增温和降水量的增加将有利于该地区的植物生长,而草原植物作为当地气候长期协同进化的产物,由于不同植物或植物功能群对气候变化的响应不同,也导致群落结构发生变异[31]。有研究认为去除优势种会导致其他物种间存在竞争作用,可能会影响植物的均匀分布[32]。综合分析,本研究结果在去除高寒草甸上的优势植物后,首先为其他物种的生长提供了有益环境,其次在温度和降雨的影响下,植物多样性发生了改变,尤其是增加温度处理作用显著,减少了物种优势度。
4 结论
研究通过去除群落优势种紫花针茅,并结合气候变化探讨高寒针茅化草甸群落特征的变化。增温显著增加了物种多样性指数,优势度指数和均匀度指数,增温条件下植物高度有增加的趋势。增雨和增温增雨交互作用,则对植物群落无显著影响。因此,在青藏高原草原中,由于其平均气温较低,温度对植物生长的影响较大,升高温度会对其生长产生积极影响。在相关性和回归分析结果中,发现随着物种数增多植物盖度,多样性指数和物种优势度指数都呈上升趋势,证明去除紫花针茅后其他物种的贡献效应较为明显,增加植物多样性有助于提高群落稳定性。
[1] Tilman D,Isbell F.Biodiversity:Recovery as nitrogen declines[J].Nature,2015,528(7582):336-337.
[2] Bakker E,Ritchic M,Olff H,etal.Herbivore impact on grassland plant diversity depends in habitat productivity and herbivore size[J].Ecol Lett,2006,9:780-788.
[3] 戎郁萍,韩建国,王培,等.放牧强度对牧草再生性能的影响[J].草地学报,2001,9(2):92-98.
[4] Thebaut E,Loreau M.Trophic interactions and the relationship between species diversity and ecosystem stability[J].Am Nat,2005,166:95-114.
[5] Bai Y F,Wu J G,Pan Q M,etal.Positive linear relationship between productivity and diversity:Evidence from the Eurasian Steppe[J].Journal of Applied Ecology,2007,44:1023-1034.
[6] 高宁宁,陈俊,张鹏莉,等.放牧对高寒嵩草草甸地上生物量空间分布的影响[J].草地学报,2014,22(2):255-260.
[7] 胡芩,姜大膀,范广洲.青藏高原未来气候变化预估:CMIP5模式结果[J].大气科学,2015,39(2):261-270.
[8] Wang S,Duan J,Xu G,etal.Effects of warming and grazing on soil N availability,species composition,and ANPP in an alpine meadow[J].Ecology,2012,93:2356-2376.
[9] Lareher W.Physiological Plant Ecology[M].Berlin:Springer-Verlag,2003.
[10] Suttle K B,Thomsen M A,Power M E.Species Interactions Reverse Grassland Responses to Changing Climate[J].Science,2007,315:640-642.
[11] 周毛措,赵德,王晓明.青海湖流域降水量季节分配对高寒草甸草原地上植物量的影响[J].草原与草坪,2013,33(4):84-87.
[12] Heisler-White J L,Knapp A K,Kelly E F.Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland[J].Oecology,2008,158:129-140.
[13] 刘琳,祝霞,孙庚.模拟增温和施肥对高寒草甸土壤酶活性的影响[J].草业科学,2011,28(8):1405-1410.
[14] 谷蕊.模拟气候变化对内蒙古克氏针茅草原生态系统及其碳交换的影响[D].呼和浩特:内蒙古大学,2015.
[15] Rui Y,Wang S,Xu Z,etal.Warming and grazing affect soil labile carbon and nitrogen pools differently in an alpine meadow of the Qinghai-Tibet Plateau in China[J].Soils Sediment,2011(11):903-914.
[17] 沙琼,黄建辉,白永飞,等.植物功能群去除对内蒙古典型草原羊草群落土壤碳、氮库的影响[J].应用生态学报,2009,20(6):1305-1309.
[18] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究[J].生态学报,1995,15(3):268-277.
[19] 杜岩功,梁东营,曹广民,等.放牧强度对嵩草草甸草毡表层及草地营养和水分利用的影响[J].草业学报,2008,17(3):146-150.
[20] Olff H,Ritchie M E.Effects of herbivores on grassland plant diversity[J].Trends Ecol Evol,1998,13:261-265.
[21] Howe H F,Brown J S,Zorn-Arnold B.A rodent plague on prairie diversity[J].Ecol Lett,2002,5:30-36.
[22] Adle P B,Milchunas D G,Sala O E,etal.Plant traits and ecosystem grazing effects comparison of US steppe and Patagonian steppe[J].Ecol Appl,2005,15:774-792.
[23] 杨中领,张家洋,楚莉莉,等.施肥和刈割对青藏高原东部高寒草甸群落生物量偿效应的影响[J].生态学杂志,2012,31(9):2276-2282.
[24] Wardle D A,Bonner K I,Barker G M,etal.Plant removals in perennial grassland:vegetation dynamics,decomposers,soil biodiversity,and ecosystem properties[J].Ecological Monographs,1999,69(4):535-568.
[25] 刘文亭,卫智军,吕世杰,等.放牧对短花针茅荒漠草原植物多样性的影响[J].生态学报,2016,37(10):1-9.
[26] 李英年,赵亮,赵新全,等.5年模拟增温后矮嵩草草甸群落结构及生产量的变化[J].草地学报,2004,12(3):236-239.
[27] 王长庭,王后基,沈振西,等.高寒矮嵩草草甸群落植物多样性和初级生产力对模拟降水的响应[J].西北植物学报,2003,23(10):1713-1718.
[28] Paul K,Courtney E C,Lara S,etal.Climate change effects on plant biomass after dominance patterns and community evenness in an experimental old-feld ecosystem[J].Global change Biology,2010,16:2672-2687.
[29] 干珠扎布.增温增雨对藏北小嵩草草甸生态系统碳交换的影响[D].北京:中国农业科学院,2013.
[30] 汪诗平,王艳芬,陈佐忠.气候变化和放牧活动对糙隐子草种群的影响[J].植物生态学报,2003,27(3)337-343.
[31] EHTLICH P R,EHRLICH A H.Extinction.The causes and consequences of the disappearance of species[M].New York:Random House,1981.
ResponseofAlpineStipapurpureaMeadowcommunitytocombinationincreaseoftemperatureandrainfallafterremovalofdominantspecies
SHI Ya-nan1,ZHONG Meng-ying1,LI Jing-jing1,SUO Nan-jia2, GA Zang-tai2,SHAO Xin-qing1
(1.CollegeofAnimalScienceandTechnology,ChinaAgriculturalUniversity,Beijing100093,China; 2.QinghaiGuinanCountygrasslandstation,Guinan813100,China)
Simulated experiment on climate changing was conducted in Haibei Prefecture,Qinghai Province,from June to August 2015.Effects of increase of air temperature (2℃) and precipitation (20%) on species diversity and community stability of Stipa purpurea was studied after removal of dominant species in the Alpine Meadow .The results showed that increase of air temperature and precipitation,and their interaction had no significant effect on plant height,coverage and aboveground biomass.Increase of air temperature significantly increased the diversity indices of Shannon-Wiener,uniformity indices of Simpson and dominance indices of Pielou.Increase of precipitation significantly increased the diversity indices of Shannon-Wiener,Simpson and Pielou.There was a positive correlation between coverage indices and plant species diversity,and between coverage indices and dominance indices.
dominate species removal;increase of precipitation;increase of air temperature;biomass;diversity
2017-04-25;
:2017-05-15
国家重点研发计划项目(2016YFC0501902)资助
史雅楠(1992- ),女,山西沁源人,硕士研究生。 E-mail:shiyanan818@126.com 邵新庆为通讯作者。
S 812
:A
:1009-5500(2017)04-0054-07
