不同丛枝菌根真菌对苜蓿生长的影响
2017-09-20张仲娟马骢毓
刘 欢,姚 拓,刘 婷,张仲娟,马骢毓
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
不同丛枝菌根真菌对苜蓿生长的影响
刘 欢,姚 拓,刘 婷,张仲娟,马骢毓
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
利用盆栽试验接种5种不同的AM真菌即地表球囊霉(Glomusversiforme,简称GV)、单孢球囊霉(Glomusmonosporum,简称GM)、根内球囊霉(Glomusintraradices,简称GI)、光壁无梗囊霉(Acaulosporalaevis,简称AL)和无梗囊霉(Acaulospora, 简称PJ-1)及未接种处理(CK),比较了不同AM菌剂处理下对苜蓿菌根侵染率、总生物量、地上生物量、地下生物量、株高等生长指标和光合作用生理指标的影响。结果表明:5种菌剂均能与苜蓿根系形成菌根,GV、GM对苜蓿根侵染率最高,分别为53.7%、50.2%;5种AM真菌对苜蓿生长、光合作用均具有促进作用,各处理间差异显著。其中,GM和GV对苜蓿株高、地上生物量、地下生物量、总生物量、光合作用具有显著的促进作用,与对照相比GV提高了苜蓿株高14.35%;GM对苜蓿地上、地下和总生物量分别增加了45.4%,40.2%和43.1%;GM、GV可显著增大气孔导度、减小胞间CO2浓度,较CK显著提高植株叶片净光合速率159.6%、163.4%。不同的AM真菌对同一宿主的促生效应不尽相同,GV、GM对苜蓿植株的促进效应最为显著,具有开发为苜蓿菌根菌剂的潜力。
丛枝菌根真菌;苜蓿;促生效应
苜蓿(Medicagosativa)是一种优质、高产、适应性强、蛋白质高的多年生豆科牧草,它的饲用价值高,营养丰富,在饲喂奶牛和肉牛方面效果显著,同时苜蓿也是一种优良的改土培肥植物[1]。随着草畜供需矛盾的日益加剧,人们为了追求经济利益盲目提高草原载畜量,使得草原急剧退化[2]。苜蓿作为主要的栽培牧草,不仅是农牧结合的纽带,更是对整个农业结构中种植业、畜牧业及加工业等产生重要的影响[3]。甘肃省苜蓿草产业种植面积稳定上升,从2004年到2013年增加了1.90万hm2[4]。虽然苜蓿可通过根瘤固氮获得生长发育所需要的氮素营养,但作为一种需磷量大的多年生作物,其对磷素的含量极为敏感[5]。由于管理模式和施肥方式较为粗放,主要依靠大量施用化肥来获取较高产量,使得田间土壤磷肥用量逐渐增加,而有效磷含量仍较低,造成苜蓿在生长过程中产生明显的缺磷症状[6-7]。一方面,增加了农民的购买性投入,推高了生产成本;另一方面,化肥的大量使用造成了一系列严重的环境污染及食品安全问题[8],这与日益增长的养殖业对优质牧草的需求形成了很大的矛盾[9]。因此,这一现象已成为限制苜蓿产业发展的瓶颈,寻找一种安全、绿色、环保的新型技术成为一项极为迫切的工作,而近些年来发展的丛枝菌根真菌接种技术为这一问题的解决带来了新的思路。
菌根(Mycorrhizae)是自然界中普遍存在的一种共生现象,是高等植物根系与土壤中一类特定真菌形成的互惠共生体,以丛枝菌根在自然界的分布最为广泛[10]。丛枝菌根真菌(Arbuscular mycorrhizae,AM)是分布广泛的土居微生物,能够与地球上大部分植物形成共生体[11]。AM真菌与宿主植物形成共生体后能以不同的途径和方式影响宿主植物的整个生长代谢过程,对植物的生长发育、养分吸收、产量、品质及抗逆性等有着很好的促进作用[12]。AM真菌通过根外菌丝为宿主植物供给营养物质,刺激植物生长,加强植物根部的保护作用,不仅可以促进植物对土壤中P、N、Cu、Fe等营养元素和水分的吸收,提高植物的产量和品质,增强植物耐受盐碱胁迫[13-19]、干旱胁迫[20]、重金属污染和病虫害[21-23]等各种逆境胁迫的能力,同时接种AM真菌能有效促进宿主植物的生长,改善宿主植物的营养状况,对植株的高度、鲜重等一系列生物量指标有显著的影响。AM真菌为植物的生长提供有效的磷元素,使豆科植物的生物量明显增加[24]。
试验选择了5种不同的AM真菌菌剂,研究其对苜蓿的生长的影响,进而探究AM真菌与苜蓿生长的相互作用机理,以期利用AM真菌提高苜蓿产量和改善品质,为我国苜蓿产业的发展提供新思路,同时为菌根技术在苜蓿产业的应用和推广提供理论依据。
1 材料和方法
1.1供试植物
紫花苜蓿品种为甘肃农业大学草业学院提供的甘农3号。
1.2供试菌种
供试菌株皆由甘肃农业大学草业学院草地生物多样性实验室提供。分别为单孢球囊霉(Glomusmonosporum,简称GM);根内球囊霉(Glomusintraradices,简称GI);地表球囊霉(Glomusversiforme,简称GV);光壁无梗囊霉(Acaulosporalaevis,简称A1);无梗囊霉属(Acaulospora,简称PJ-1)。5种AM真菌在接种之前,皆采用玉米为宿主植物进行扩繁,接种物中包括AM真菌孢子、根外菌丝及被侵染宿主植物根段的沙土混合物。
1.3试验地条件及供试土壤
盆栽试验在甘肃农业大学温室进行,试验期间温室温度为(25±4)℃,光照时间为14~15 h。供试土壤采于甘肃省农业科学院试验地,河砂购买于市场。供试土壤、河砂均需过2 mm土壤筛,过筛之后在1×105Pa下进行湿热灭菌2 h。采用规格为16 cm×20 cm×12 cm的塑料花盆作为培养容器,装土之前,用70%乙醇对花盆进行系统消毒,供试土壤理化性质(表1)。

表1 供试土壤理化性质
1.4试验设计
以苜蓿为宿主植物,设置5个接种处理和1个未接种对照(CK),每个处理重复3次。挑选饱满、均一的宿主植物种子,采用次氯酸钠溶液(有效氯成份≤10%)消毒,用蒸馏水多次冲洗,转移至内含滤纸的培养皿,放置于培养箱发芽。等到种子发芽后,移栽至花盆,每盆栽种20株苜蓿。播种时,每个花盆装灭菌基质2.5 kg,然后将15 g接种剂(1 g菌剂约含60个孢子)平铺其上,再覆盖0.5 kg灭菌基质。未接种处理每盆施用15 g灭菌接种物和15 mL无灭菌接种物水滤液,以保证微生物区系的一致性。花盆随机摆放,种植期间每隔一天浇水一次,以保证苜蓿的正常生长。
1.5样品收获及分析测定
1.5.1 AMF菌根侵染率测定 采用曲利苯兰染色法[24]测定菌根侵染率,于种植后第90 d,收获苜蓿植株。用自来水不断冲洗苜蓿根系,洗净后,置于10% KOH溶液中,85℃水浴加热40 min。清水冲洗后置于1% HCl中酸化,1~2 min后将根系放置于曲利苯蓝染色液中,85℃水浴染色40 min,取出后用自来水反复冲洗,将染色根切成长度为1 cm的根段,在解剖镜下进行观察,利用交叉网格线法测定菌根侵染率。
1.5.2 苜蓿植株株高的测量和地上、地下生物量的测量 试验结束(90 d)时对苜蓿植株的株高进行测量;经105℃杀青15 min后在70℃下烘干至恒重,分别称量地上和地下部分干重。
1.5.3 苜蓿叶片光合作用的测定 采用LI-6400 便携式光合仪活体测定苜蓿叶片的光合生理生态指标。选择生长健康、长势一致、无病斑、光照均一的同一苜蓿叶位作为测定对象(取样部位为植株从上往下数第3片叶片),主要测定指标包括:叶片的光合速率、胞间CO2浓度、植株叶片气孔导度等生理指标[25]。
1.5.4 计算公式 菌根侵染率F=(被侵染的根段数/镜检的总根段数)×100%
1.6数据分析
试验数据采用SPSS(v17.0)软件计算平均值±标准误(SE)。所有测定数据的平均值采用LSD法在显著水平为5%条件下进行比较,采用Excel 2007软件制图。
2 结果与分析
2.1接种AM真菌对苜蓿根侵染率的影响
不同AM真菌菌剂处理对宿主植物的菌根侵染率各不相同,未接种处理中均加入了灭菌后的接种物,其侵染率为0;对于接种处理,苜蓿与5种AM真菌均建立了良好的菌根共生关系。5种菌剂中,菌剂GV、GM表现较好,侵染率分别是(53.7±0.86)%、(50.2±0.43)%;菌剂PJ-1侵染率最低(37.8±0.20)%;菌剂GI、AL差异显著(P<0.05),分别为(44.5±0.53)%、(39.2±0.52)%。各菌剂间均差异显著,菌剂GV、GM侵染率显著高于其他菌株(P<0.05)(图1)。

图1 不同AMF处理下的苜蓿菌根的侵染率Fig.1 Effect of AMF on of AM colonization rate
2.2接种AM真菌对苜蓿生长影响
2.2.1 接种AM真菌对苜蓿株高的影响 不同AM真菌处理对苜蓿株高产生了显著的影响(图2)。接菌处理与对照相比在株高的生长动态上存在一定的差异,菌剂GV表现最佳,与其他菌剂处理有显著差异(P<0.05),其处理下的株高为(36.57±0.25)cm,与CK相比提高了14.35%;在菌剂PJ-1处理下株高变化与CK相比变化不显著。其中5个接菌处理显著高于CK(P<0.05),除菌剂GI、AL外,其他接菌处理间均差异显著(P<0.05)。
2.2.2 接种AM真菌对苜蓿生物量的影响 苜蓿接种AM真菌后,与CK相比,5种AM真菌均显著增加了苜蓿地上地下生物量和总生物量(图3-5)。菌剂GV、GM表现最佳,与其他菌剂处理具有显著性差异(P<0.05);菌剂GM相对于CK地上、地下和总生物量分别增加了45.4%,40.2%和43.1%;菌剂GV表现也较为优秀,与对照相比,可显著增加苜蓿地上、地下生物量及总生物量;菌剂PJ-1,GI和AL对苜蓿生物量的促进效果相近,三者间无显著性差异(P>0.05)。

图2 不同AMF处理下苜蓿的株高Fig.2 Effect of AMF on plant height of alfalfa

图3 不同AMF处理下的苜蓿地上生物量Fig.3 Effect of AMF on shoot biomass of alfalfa
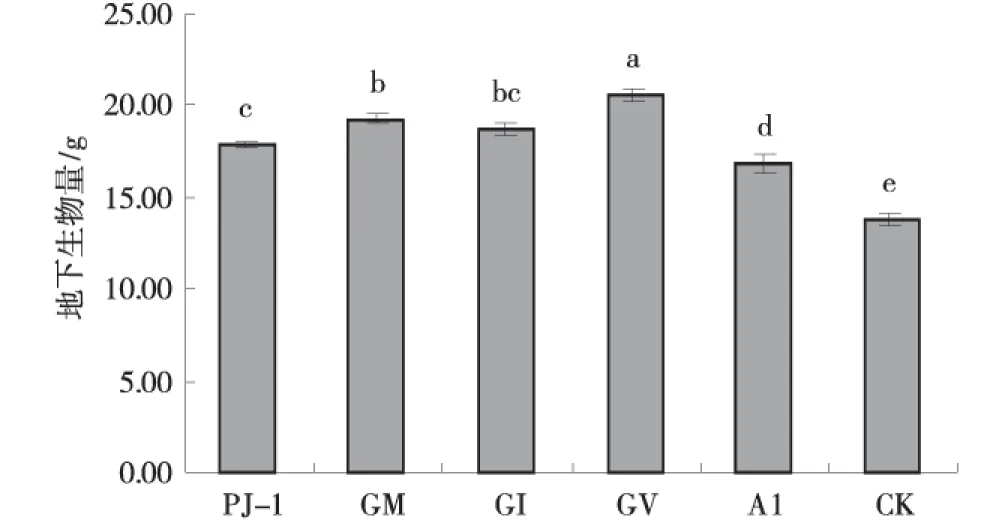
图4 不同AMF处理下的苜蓿地下生物量Fig.4 Effect of AMF on root biomass of alfalfa

图5 不同AMF处理下的苜蓿总生物量Fig.5 Effect of AMF on total biomass of alfalfa
2.3接种AM真菌对苜蓿光合作用的影响
5种接菌处理均促进了苜蓿叶片的光合速率。其中GM、GV促进效果最好,分别为(10.873 3±0.1411)μmol/(m2·s)、(11.065 0±0.073 3)μmol/(m2·s),与CK相比分别增加了159.6%、163.4%,但其二者间无显著性差异(P>0.05)。PJ-1与AL表现效果很相似,二者与CK相比分别增加了82.4%、86.2%,二者之间也无显著性差异。接种处理下苜蓿叶片的气孔导度变化趋势与光合作用相似。5种处理均显著增加了苜蓿叶片的气孔导度,其中GM、GV表现最佳,与CK相比,增加了155.5%、174.4%,其二者之间无显著差异(P>0.05)。PJ-1表现较弱,与CK相比仅增加59.4%,但仍与CK差异显著(P<0.05)。接种AM真菌后,苜蓿叶片胞间CO2浓度的变化趋势与光合速率、气孔导度相反。其中,未接种处理下苜蓿叶片胞间CO2浓度最高,为378.468 4 μmol/mol,5种接菌处理下植株叶片胞间CO2浓度的大小分别为GV 图6 不同AMF处理下的苜蓿光合速率Fig.6 Effect of AMF on net photosynthetic rate 图7 不同AMF处理下的苜蓿叶片气孔导度Fig.7 Effect of AMF on stomatal conductance 图8 不同AMF处理下的苜蓿胞间CO2浓度Fig.8 Effect of AMF on intercellular of alfalfa 2.4AMF对苜蓿生长的重要性 将供试菌株的侵染率、地上生物量、地下生物量、总生物量、株高、苜蓿叶片光合速率、气孔导度及胞间CO2浓度8个性状作为自变量,进行主成分分析。在8个因子中,第1个因子的贡献率达到92.309%,因此,在主成分分析中只需取第1个主成分。从综合得分及排名分析,GV得分最高,促进作用最好;PJ-1得分较低,促进较弱(表2)。 表2 苜蓿接种AMF主成分得分及综合得分 3.1AM真菌对植物菌根侵染率的影响 菌根侵染率是评价AM真菌与植物建立共生关系与否的重要指标,是讨论菌根对植物抗逆性影响的前提。AM真菌与宿主植物根系亲和力及二者之间的相互选择性决定着菌根的生长发育和其功能效应的发挥程度[26-27]。当菌根侵染率越高,植物菌根化程度也会相应越高,对植物促进作用、抗逆性能力的增加也会越明显。试验中,所选用的5种丛枝菌根真菌均能有效侵染苜蓿,其菌根侵染率为37.86%~53.73%,由此说明苜蓿与5种AM真菌均建立了良好的共生关系。而前人采用同种真菌在小麦、白三叶、番茄、梨等植物上试验[28-31],也得到了相似的结果,说明这些真菌具有较广的宿主适应性。此外,研究还发现,不同AM菌剂的侵染率亦有明显差异。表明宿主植物对不同AM真菌的响应不同,同时AM真菌对宿主植物存在选择性。Johnson等[32]的理论能较好的解释这一现象,不同种类的植物可直接或间接的通过自身根系环境,对群落中不同的AM真菌进行选择。另外,前人在葡萄、杨梅、黄瓜、牡丹等植物上也已经验证了这一理论[33-36]。由此可见,AM真菌的侵染率是受宿主植物和AM真菌本身特性双重因素调控,这一结果也提醒人们,在AM真菌的应用过程中,要注意宿主植物和所选用菌株的匹配性。 外界环境条件对菌根侵染率的高低也是一个重要的因素,如接种AM真菌多使用的基质类型、肥力状况、疏松程度[37],温度,水分、通气状况等因素,另外土壤中微生物如荧光假单孢菌、芽孢杆菌、根瘤菌、解磷细菌、固氮菌等大多微生物均与AM真菌具有协同作用[39-40]。秦芳玲等[4]通过对红三叶根际接种AM真菌及3种解磷细菌后发现,其中两种解磷细菌与AM真菌双接种处理显著高于单接种处理下的菌根侵染率。丁效东等[42]通过对大豆根际接种根瘤菌与AM真菌,结果表明,双接种处理下的侵染率显著高于单接种处理下的菌根侵染率。试验表明,所选用的土壤基质均经过高温灭菌,基质中几乎没有可与AM真菌共生的菌群,因此,在一定的程度上降低了菌根的侵染率,在实际的大田应用中,AM真菌对植物的侵染率可能会高于此次试验中侵染率的数值。 3.2AM真菌对植物生长发育的影响 生物量是衡量植株生长状况的重要指标之一[43-44]。紫花苜蓿具有发达直根系的植物,其对菌根的依赖性较强,接种AM真菌后可显著增加苜蓿根部的吸收面积,便于苜蓿根系在土壤中生长,从而促进苜蓿地上部分的生长。试验中,接种不同种AM真菌后,苜蓿的生物量均有不同程度增加,表现出了一定程度的菌根生态效应。表明,不同AM真菌对不同宿主植物的生长产生促进效应,可能是由于AM真菌侵染植物根系之后,通过根外菌丝的介入,扩大了植物根系的吸收范围,增强了植物对水分和营养元素的吸收。而较为充足的营养元素和水分的供应又促进了作物地上部分的生长,从而对植物的生长起到了良好促进作用。张瑞[45]对紫花苜蓿接种AM真菌,发现接种株株高、地上和地下生物量均明显高于对照株,表明AM真菌对紫花苜蓿的生长具有正效应。任爱天等[46]通过设置不同磷水平,对苜蓿接种AM真菌发现,不同AM处理下均显著增加了紫花苜蓿地上干重、根重、根长度、株高等。同时试验还发现,接种不同菌剂导致植株生物量之间存在明显差异。推测其可能的原因一是不同的AM真菌对营养元素和水分的吸收能力不同;二是与该种AM真菌和植物的匹配性有关。这与王茜等[47]对垂穗披碱草接种AM真菌的研究结果相似。 3.3AM真菌对植物光合作用的影响 植物生活史中最重要和基础的代谢过程是光合作用,而光合作用的强弱能够用作判断植物抗逆性强弱的指标[48]。试验结果发现,接种AM真菌后能够增加植物的光合作用,而光合作用的强弱主要与植物叶片气孔通透性和叶肉细胞的光合活性息息相关[49]。苜蓿接种AM真菌后,增加了其叶片的光合速率和气孔导度,同时降低了植物叶片的胞间CO2浓度。植物叶皮气孔导度的增大,使得外界环境中更多CO2进入了叶肉细胞中,为叶片光合作用提供足量原料;同时促进了CO2同化利用效率,降低了胞间CO2浓度,进一步提高了光合效率,促进苜蓿植株生长,因而改善植物的光合参数,提高植株叶片光合速率,间接地使植物获得更多的碳水化合物。宋会兴等[50]通过对干旱生境中三叶鬼针草接种AM真菌后,研究其光合特征发现,光合速率会随着干旱程度的加深而降低,但接种处理下植株的光合速率明显高于未接种处理下植株的光合速率。同时,在中度水分胁迫条件下,接种植株的气孔导度明显高于未接种植株,而其胞间CO2浓度低于未接种处理。孙秀秀等[51]利用两种复合丛枝菌根真菌菌剂对苗期中的黄瓜进行接种,结果表明,与未接种植株相比,两种接种菌剂均可显著提高黄瓜叶片的光合速率及气孔导度,并且使得植物叶片CO2浓度有所降低。也有研究报道,无论是在常温及低温下对玉米接种AM真菌,接种处理的叶片光合速率及气孔导度均大于未接种处理,而在两个温度处理下,接种AM真菌均可降低玉米叶片的胞间CO2浓度[52]。这均与本次试验结果相似。同时苜蓿光合速率、气孔导度的变化规律与植株地上生物量变化规律相同,这也印证了接种AM真菌可提高植株光合特性,从而促进植株生长这一结论。 试验所采用的5种AM真菌均能促进苜蓿株高、地上生物量、地下生物量、总生物量等生长指标,且不同菌株对苜蓿促进效果不同,其中单孢球囊霉(GM)和地表球囊霉(GV)促进效果较为显著,具有开发成为菌根真菌菌肥的潜力。 [1] 张立全,张凤英,哈斯阿古拉.紫花苜蓿耐盐性研究进展[J].草业学报,2012,21(6):296-305. [2] 李重祥.紫花苜蓿高效丛枝菌根菌的筛选及利用研究[D].呼和浩特:内蒙古农业大学,2010. [3] 冯国胜,康国芳,施进文.紫花苜蓿的栽培及应用[J].中国畜牧兽医文摘,2014(6):196+192. [4] 张洁冰,南志标,唐增.美国苜蓿草产业成功经验对甘肃省苜蓿草产业之借鉴[J].草业科学,2015(8):1337-1343. [5] 葛选良,杨恒山,刘晶,等.施磷水平对紫花苜蓿生长及草产量的影响[J].内蒙古民族大学学报(自然科学版),2009(5):509-513. [6] 邰继承,杨恒山,张军,内蒙古通辽市玉米田改种紫苜蓿的优势分析[J].草业科学,2012,29(1):150-155. [7] 冯博正,刘晓静,郝凤,等.NO3-N/NH4+-N 混合氮不同水平对紫花苜蓿生长特性的研究[J].草原与草坪,2015,35(6):35-39. [8] 张凡凡,于磊,鲁为华,等.高效利用磷肥提高我国苜蓿生产力的研究进展[J].草食家畜,2013(5):6-11. [9] Li Y Y,Wang X J,Dou C Y,etal.Effects of host plant and different cultivation densities on the propagation of arbuscularmycorrhizalfungi[J].Acta Prataculture Sinca,2013,22(5):128-135. [10] 李媛媛,王晓娟,豆存艳,等.四种宿主植物及其不同栽培密度对AM真菌扩繁的影响[J].草业学报,2013,22(5):128-135. [11] Simth S E,Read D J.Mycorrhizal Symbiosis.[M].NewYork:Academic Press,2008. [12] 王强,王茜,王晓娟,等.AM真菌在有机农业发展中的机遇[J].生态学报,2016(1):11-21. [13] Baird J,Walley F L,Shirtliffe S J.Arbuscularmtcorrhizal fungi colonization and phosphorus nutrition in organic filed pea and lentil[J].Mycorrhiza,2010,20:541-549. [14] Atul-Nayyar A,Hamel C,Hanson K,etal.The arbuscularmycorrhizal symbiosis links N mineralization to plant demand[J].Mycorrhiza,2009,19:239-246. [15] Liu A,Hamel C,Hamilton R I,etal.Acquisition of Cu,Zn,Mn and Fe by mycorrhizal maize(ZeamaysL.) grown in soil at different P and micronutrient leves[J].Mycorrhiza,2000(9):331-336. [16] Caris C,Hördt W,Hawkins H J,etal.Studies of iron transport by arbuscularmycorrhizal hyphae from soil peanut and sorghum plants[J].Mycorrhiza,1998,8:35-39. [17] Neumann E,Schmid B,Römheld V,etal.Extraradical development and contribution to plant performance of an arbuscularmycorrhizal symbiosis exposed to complete or partial rootzonedrying[J].Mycorrhiza,2009,20:21-23. [18] Toussaint J P,Smith F A,Smith S E.Arbuscularmycorrhizal fungi can induce the production of phytochemicals in sweet basil irrespective of phosphorus nutrition[J].Mycorrhiza,2007,17:291-297. [19] García I V,Mendoza R E.Arbuscularmycorrhizal fungi and plant symbiosis in a saline-sodicsoil[J].Mycorrhiza,2007,17(3):167-174. [20] Mena-Violante H G,Ocampo-Jiménez O,Dendooven L,etal.Arbuscularmyorrhizal fungi enhance fruit growth and quality of chileancho(CapsicumannuumL.cv San Luis) plants exposed to drought[J].Mycorrhiza,2006,16:261-267. [21] Yu Y,Zhang S Z,Huang H L.Behavior of mercury in a soil-plant syesten as affected by inoculuation with the arbuscularmycorrhizal fungus Glomusmosseae[J].Mycorrhiza,2010,20:407-414. [22] Kamińska M,Klamkowski K,Berniak H,etal.Response of mycorrhizal periwinkle plants to aster yellows phytoplasmainfection[J].Mycorrhiza,2010,20:161-166. [23] Elsen A,Gervacio D,Swennen R,etal.AMF-induced bio control against plant parasitic nematodes inMusasp.a systemic effect[J].Mycorrhiza,2008,18:251-256. [24] Koske R E,Gemma J N.A modified procedure for staining roots to detect VA mycorrhizas[J].Mycological Research,1989,92(4):486-488. [25] 任爱天,鲁为华,杨洁晶,等.不同磷水平下AM真菌对紫花苜蓿生长和磷利用的影响[J].中国草地学报,2014(6):72-78. [26] 冯远娇,陈卓娜,王建武,等.Bt玉米丛枝菌根真菌侵染率与养分含量的变化研究[J].中国生态农业学报,2010,18(3):486-491. [27] 张淑彬,王红菊,王幼珊,等.不同育苗基质对黄瓜生长及丛枝菌根真菌侵染的影响[J].中国农学通报,2011,27(10):275-279. [28] 马放,苏蒙,王立,等.丛枝菌根真菌对小麦生长的影响[J].生态学报,2014,34(21):6107-6114. [29] 王晓英,王冬梅,陈保冬,等.丛枝菌根真菌群落对白三叶草生长的影响[J].生态学报,2010,30(6):1456-1462. [30] 范燕山.丛枝菌根真菌对有机质栽培番茄生长的影响[D].长沙:湖南农业大学,2008. [31] 韦洁敏.丛枝菌根真菌对梨苗生长及抗旱耐热性的影响研究[D].重庆:西南大学,2012. [32] Johnson N C,Graham J H,Smith F A.Functioning of mycorrhzizal associations along the mutualism-parasitism continuum[J].New Phytologist,1997,135:575-585. [33] 郭修武,李坤,郭印山,等.丛枝菌根真菌对连作土壤中葡萄生长及根系分泌特性的影响[J].沈阳农业大学学报,2009,40(4):392-395. [34] 王小雪.接种丛枝菌根真菌对杨梅幼苗生长及铁素吸收效应的影响[D].重庆:西南大学,2013. [35] 韩冰,徐刚,郭世荣,等.丛枝菌根真菌对苗期黄瓜生长及生理特性的影响[J].江苏农业学报,2012,28(6):1392-1397. [36] 陈丹明,郭娜,郭绍霞.丛枝菌根真菌对牡丹生长及相关生理指标的影响[J].西北植物学报,2010,30(1):131-135. [37] 罗英,王怀玉.基质对VA菌根真菌的侵染及孢子产量的影响[J].四川师范大学(自然科学版),2003,26(3):302-305 [38] 董昌金,赵斌.影响丛枝菌根真菌孢子萌发的几种因素研究[J].植物营养与肥料学报,2003,9(4):489-494. [39] Vivas A,Marulada A,Ruiz-Lozano J M,etal.Influence of a Bacillus sp.on physiological activities of two arbuscularmycorrhizal fungi and on plant responses to PEG-induced drought stress[J].Mycorrhiza,2003,13(5):249-256. [40] Xavier L J C,Germida J J.Bacteria associated with Glomusclarum spores influence mycorrhizal activity[J].Soil Biology and Biochemistry,2003,35(3):471-478. [41] 秦芳玲,田中民.同时接种解磷细菌与丛枝菌根真菌对低磷土壤红三叶养分利用的影响[J].西北农林科技大学学报(自然科学版),2009,37(6):151-157. [42] 丁效东,张林,李淑仪,等.丛枝菌根真菌与根瘤菌接种对大豆根瘤分布及磷素吸收的影响[J].2012,18(3):662-669. [43] 马文彬,姚拓,张玉霞,等.植物促生菌剂对甘草生长影响的研究[J].草原与草坪,2014,34(1):13-16. [44] 荣良燕,姚拓,刘青海,等.复合菌肥代替部分化肥对玉米生长的影响[J].草原与草坪,2012,32(3):65-69. [45] 张瑞.盐碱土壤接种菌根对紫花苜蓿的综合效应[D].呼和浩特:内蒙古农业大学,2009. [46] 任爱天,鲁为华,杨洁晶,等.不同磷水平下AM真菌对紫花苜蓿生长和磷利用的影响[J].中国草地学报,2014,36(6):72-77. [47] 王茜,董梅,王强,等.不同丛枝菌根真菌对青藏高原高寒草原优良牧草垂穗披碱草生长的促生效应[J].云南农业大学学报,2014,29(6):840-846. [48] 吴雪霞,陈建林,查丁石.低温胁迫对茄子幼苗叶片光合特性的影响[J].华北农学报,2008,23(5):185-189. [49] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33(1):317-345. [50] 宋会兴,彭远英,钟章成.干旱生境中接种丛枝菌根真菌对三叶鬼针草(BidensPilosaL.)光合特征的影响[J].2008,28(8):3744-3751. [51] 孙秀秀,晋文娟,李衍素,等.两种捷克丛枝菌根真菌组合菌剂对设施黄瓜苗期生长的影响[J].中国蔬菜,2016(4):34-37. [52] 陈笑莹,宋凤斌,朱先灿,等.低温胁迫下丛枝菌根真菌对玉米幼苗形态、生长和光合的影响[J].华北农学院学报,2014,29(4):205-212. EffectofdifferentarbuscularmycorrhizalfungionthegrowthofMedicagosativa LIU Huan,YAO Tuo,LIU Ting,ZHANG Zhong-juan,MA Cong-yu (CollegeofPrataculturalScience,GansuAgriculturalUniversity/KeyLaboratoryofGrasslandEcosystem,MinistryofEducation/PrataculturalEngineeringLaboratoryofGansuProvince/Sino-U.S.CenterforGrazinglandEcosystemSustainability,GansuLanzhou730030,China) Five arbuscularmycorrhizal (AM) fungi (Glomusversiforme,G.monosporum,G.intraradices,AcaulosporalaevisandAcaulospora) were used to inoculate the pot alfalfa in order to study their effects on rate of colonization,shoot biomass,root biomass,height and photosynthesis of alfalfa.The results showed that alfalfa could be colonized by 5 AM fungi and form mycorrhiza,in which,the colonization percentages with GV,GM were 53.7%,50.2%.All tested fungi could promote the growth and photosynthesis of alfalfa and the difference among treatments was significant.G.versiformeandG.monosporumpromoted the root biomass,shoot biomass and photosynthesis.Compared with control,GV increased the height of alfalfa by 14.35%;GM increased the shoot biomass,root biomass and total biomass by 45.4%,40.2% and 43.1%;GM,GV enlarged stomatal conductance and reduced intercellular CO2concentration significantly,increased net photosynthesis rate by 159.6%,163.4%.However,the other AM fungi did not promote the growth of alfalfa.Different AM fungi showed varied promotion effects on alfalfa growth.The promising AM fungi wereG.versiformeandG.monosporum. arbuscularmycorrhizae;alfalfa;growth-promoting effect 2016-06-28; :2017-40-09 国家自然科学基金(41561006)项目“三江源区主要栽培牧草根际促生菌(PGPR)多样性及促生机理研究”资助 刘欢(1990- ),女,甘肃兰州人,在读硕士研究生。 E-mail:810827630@qq.com 姚拓为通讯作者。 S 541 :A :1009-5500(2017)04-0061-08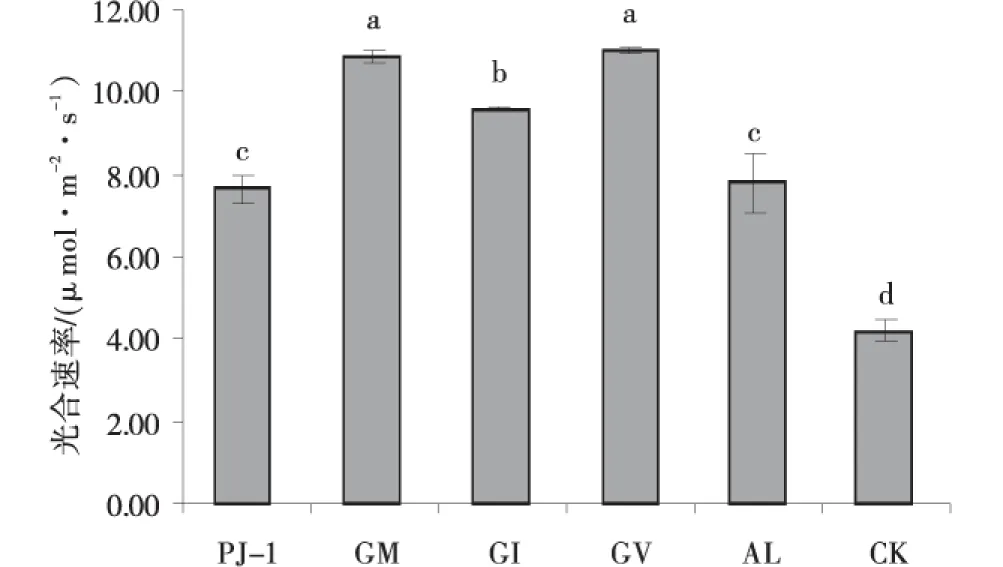



3 讨论
4 结论
