不同处理菜豆植株对西花蓟马体内解毒酶活性的影响
2017-09-16郅军锐岳文波
牟 峰,郅军锐,岳文波,曾 广
(贵州大学昆虫研究所,贵州省山地农业病虫害重点实验室,贵阳 550025)
不同处理菜豆植株对西花蓟马体内解毒酶活性的影响
牟 峰,郅军锐*,岳文波,曾 广
(贵州大学昆虫研究所,贵州省山地农业病虫害重点实验室,贵阳 550025)
为明确植物的诱导抗性对西花蓟马体内酶活性的影响,通过西花蓟马取食、机械损伤、外源茉莉酸和水杨酸甲酯诱导处理菜豆植株,研究西花蓟马2龄若虫取食不同处理菜豆植株后,其体内多功能氧化酶(MFO)、羧酸酯酶(CarE)、谷胱甘肽S-转移酶(GSTs)、乙酰胆碱酯酶(TChE)的变化。结果表明,西花蓟马2龄若虫取食不同处理的菜豆后,体内多功能氧化酶和乙酰胆碱酯酶活性变化相同,均是在24 h和36 h显著低于取食健康植株的。除取食茉莉酸处理的西花蓟马体内谷胱甘肽S-转移酶活性在24 h受到显著抑制外,西花蓟马若虫取食水杨酸甲酯、蓟马危害后体内的谷胱甘肽S-转移酶活性均高于对照,取食机械损伤处理的西花蓟马体内酶活性在不同取食时间下均与对照差异不显著。取食蓟马危害和茉莉酸处理的西花蓟马虫体羧酸酯酶活性在6 h和36 h均受到显著抑制,而取食机械损伤和水杨酸甲酯诱导的西花蓟马体内酶活性只有在36 h受到明显抑制。以上结果说明外源因子诱导的植物抗性能够干扰西花蓟马体内解毒机制,削弱了其对有毒次生物质的解毒代谢功能。
西花蓟马;菜豆;诱导抗性;解毒酶
植物在与昆虫的协同进化中,形成了一系列可相互适应机制,外源信号物质和昆虫取食均能够诱导植物产生抗性,进而对取食的害虫生长发育产生不利影响(桂连友等,2004;彭露等,2010)。面对植物的防御体系,植食性昆虫在选择压力的作用下演化出多种反防御机制以维持种群繁衍(Arimura and Kost, 2005)。其中解毒机制在昆虫的众多的反防御机制中发挥重要的作用。昆虫解毒酶是一类异质酶系,昆虫通过调节体内解毒酶活性对杀虫剂、致癌物以及植物产生的次生性代谢物等有毒物质进行解毒,以适应植物的变化,减弱或免受植物诱导抗性产生的有毒次生物质对昆虫的毒害(Kostaropoulosetal., 2001;邱立红和张文吉,2001)。解毒酶主要包括:多功能氧化酶(MFO)、羧酸酯酶(CarE)、谷胱甘肽S-转移酶(GSTs)、乙酰胆碱酯酶(TChE)等。Yu(1986)研究发现,植物次生性代谢物对昆虫体内解毒酶的诱导作用与杀虫剂的诱导解毒酶系统作用相似。已有报道,植物的次生性代谢物能够影响昆虫的解毒酶活性,从而使昆虫对杀虫剂的敏感性产生影响(Robertsonetal., 1990)。不同的寄主植物中含有的次生性代谢物种类和含量存在差异,因此对植食性昆虫体内酶活性的影响也不同(宋月芹等,2009),经虫害、茉莉酸和水杨酸及其化合物诱导寄主,虫体内解毒酶活性会产生不同的响应,其活性或升高或被抑制(吴国昭等,2012;胡海燕,2013)。
西花蓟马Frankliniellaoccidentalis是一种全球性的入侵害虫,自2003年发现后,在我国迅速扩散,给我国蔬菜和花卉产业造成严重的损失(吕要斌等,2011)。研究西花蓟马与植物之间的关系,有助于揭示寄主植物与西花蓟马的防御与反防御机制,为制定安全有效的防治策略奠定基础。很多学者研究了不同诱导因子对西花蓟马及其寄主菜豆的影响,如前人研究发现西花蓟马取食、机械损伤、外源水杨酸甲酯(MeSA)及茉莉酸(JA)诱导处理后会引起菜豆叶片营养物质和防御酶活性的变化(从春蕾等,2013,2014;田旭涛等,2013)。目前有关植物诱导抗性对西花蓟马的影响,多数集中在不同诱导处理对西花蓟马生长发育影响方面,如杨帆(2009)研究发现,西花蓟马取食茉莉酸甲酯、水杨酸等诱导处理的黄瓜叶片后,发育时间显着延长,蛹重下降,西花蓟马的生存适合度降低。牟峰等(2014)研究也表明西花蓟马取食外源茉莉酸、水杨酸甲酯、西花蓟马为害处理的菜豆植株后其未成熟期的发育时间均延长,存活率不同程度的下降。而有关其生理解毒机理的研究较少,只有陈锐芬(2011)报道寄主转换后不同寄主种群的西花蓟马虫体的羧酸酯酶、谷胱甘肽硫转移酶、乙酰胆碱酯酶等解毒酶的活性初期被激活、再被抑制,最后达到稳定。本试验以蓟马危害、机械损伤、水杨酸甲酯、茉莉酸处理菜豆植株,研究不同处理菜豆植株对西花蓟马2龄若虫体内解毒酶活性的影响,从生理生化角度分析植物诱导抗性对西花蓟马解毒机理的影响,为探讨利用植物诱导抗性防御害虫提供理论依据。
1 材料与方法
1.1 供试昆虫和供试植物
1.1.1供试昆虫
西花蓟马:实验室长期用菜豆PhaseolusvulgarisL.豆荚饲养的西花蓟马。
1.1.2供试植物
菜豆植株:在人工气候箱(温度25℃,湿度70%,光照14 L ∶10 D)内培养菜豆植株,以营养钵单株培育清洁苗,待菜豆长至2片真叶时作为供试寄主植物,生长期间不使用任何农药。
1.2 试验方法
1.2.1菜豆植株处理
(1)蓟马危害植株,记为DTF:在供试菜豆植株每片真叶接种10头虫龄一致的西花蓟马2龄若虫,每株共接20头作为西花蓟马为害后的植株,接虫1 d后剔除若虫。
(2)机械损伤植株,记为MD:在供试菜豆植株的每真叶用针头(直径0.29 mm)从正面刺透叶片10次,1 d后用于试验。
(3)茉莉酸诱导植株,记为JA:在供试菜豆植株叶片上,喷洒浓度为1.0 mmol/L茉莉酸,喷至叶片全湿为准,1 d后用于试验。
(4)水杨酸甲酯诱导植株,记为MeSA:在供试菜豆植株叶片上,喷洒浓度为5 mmol/L水杨酸甲酯,喷至叶片全湿为准,1 d后用于试验。
(5)健康植株,记为CK:健康菜豆植株作为对照。
1.2.2解毒酶活性测定
酶源制备参考刘建业等(2014)的方法,并稍作修改。虫源为西花蓟马2龄若虫50头,每组处理重复4次。多功能氧化酶(MFO)、羧酸酯酶(CarE)、谷胱甘肽-S-转移酶(GSTs)和乙酰胆碱酯酶(TChE)的测定均根据南京建成科技有限公司的试剂盒说明书的操作步骤进行。
1.3 数据统计与分析
以Excel 2003对数据进行统计,所获数据以平均值±标准误表示,采用SPSS 19.0软件进行分析,以Duncan氏新复极差法检验各处理间的差异显著性。
2 结果与分析
2.1 多功能氧化酶(MFO)活性的变化
取食不同处理的菜豆植株后西花蓟马若虫体内多功能氧化酶活性变化不同(图1)。6 h时,只有取食机械损伤的虫体酶活性与对照差异不显著,西花蓟马取食虫害菜豆、水杨酸甲酯以及茉莉酸处理的叶片,后其体内酶活性与对照相比均受到明显抑制,且蓟马危害和茉莉酸处理的抑制作用要强于水杨酸甲酯处理。24 h时,西花蓟马取食不同处理虫体酶活性均受到明显抑制,且以取食水杨酸甲酯处理的酶活性最低,与取食其它3种处理间差异显著。36 h时,西花蓟马虫体酶活性和上一个时间段相同,取食各处理的酶活性均显著低于对照,取食机械损伤处理的酶活性高于其它3个处理。
西花蓟马若虫体内的多功能氧化酶活性不论在哪一种处理下,随取食时间的延长均有一个先下降后上升的过程,均是24 h时多功能氧化酶活性最低,且与6 h和36 h之间差异明显。
2.2 乙酰胆碱酯酶(TChE)活性的变化
西花蓟马2龄若虫取食不同处理菜豆植株后,其体内乙酰胆碱酯酶活性变化也不同(图2)。在6 h时,取食蓟马危害与水杨酸甲酯处理的虫体酶活性均显著低于对照,但取食机械损伤与茉莉酸处理的虫体酶活性与对照差异不显著。24 h时,取食各处理的虫体酶活性均显著低于对照,但不同处理之间差异不同,以取食蓟马危害处理的酶活性下降最明显,取食水杨酸甲酯诱导的下降最少。36 h时,同样取食各处理的虫体酶活性均也显著低于对照,但各处理的酶活性变化与24 h相反,以取食水杨酸甲酯处理的酶活性最低,而取食蓟马危害、机械损伤和茉莉酸处理的酶活性之间差异不显著。
取食不同处理西花蓟马若虫体内乙酰胆碱酯酶活性在不同取食时间下,均呈现先升高再降低的变化过程。只有取食机械损伤处理的西花蓟马体内酶活性在24 h与6 h差异不显著,但显著高于36 h;取食其余处理的体内酶活性均是24 h显著高于6 h和36 h。

图2 不同处理诱导的菜豆植株对西花蓟马体内乙酰胆碱酯酶活性的影响Fig.2 Changes of TChE activity in Frankliniella occidentalis feeding kidney bean induced by different treatments
2.3谷胱甘肽S-转移酶(GSTs)活性的变化
不同处理菜豆植株对取食西花蓟马若虫体内谷胱甘肽S-转移酶活性有明显影响(图3)。6 h时,取食蓟马危害、水杨酸甲酯和茉莉酸处理的谷胱甘肽S-转移酶活性均显著高于对照处理,且取食水杨酸甲酯和茉莉酸处理的酶活性最高,但取食机械损伤处理的酶活性与对照差异不显著。24 h时,除取食蓟马危害处理的谷胱甘肽S-转移酶活性略有升高外,取食其它处理的酶活性均受到一定的抑制,其中取食茉莉酸处理的酶活性最低仅为对照的32.13%。36 h时,取食各处理下蓟马体内酶活性的变化和6 h类似。
取食蓟马危害和机械损伤的酶活性在不同时间下的变化相似,均是24 h酶活性最高,随着取食时间的延长,两种处理的酶活性都有一定程度下降,其中取食机械损伤处理下降幅度要大于取食蓟马危害处理。取食水杨酸甲酯处理的酶活性随着取食时间的延长而逐渐升高,至36 h时达到最高峰。取食茉莉酸处理的酶活性24 h时最低,但在36 h时酶活性最高。
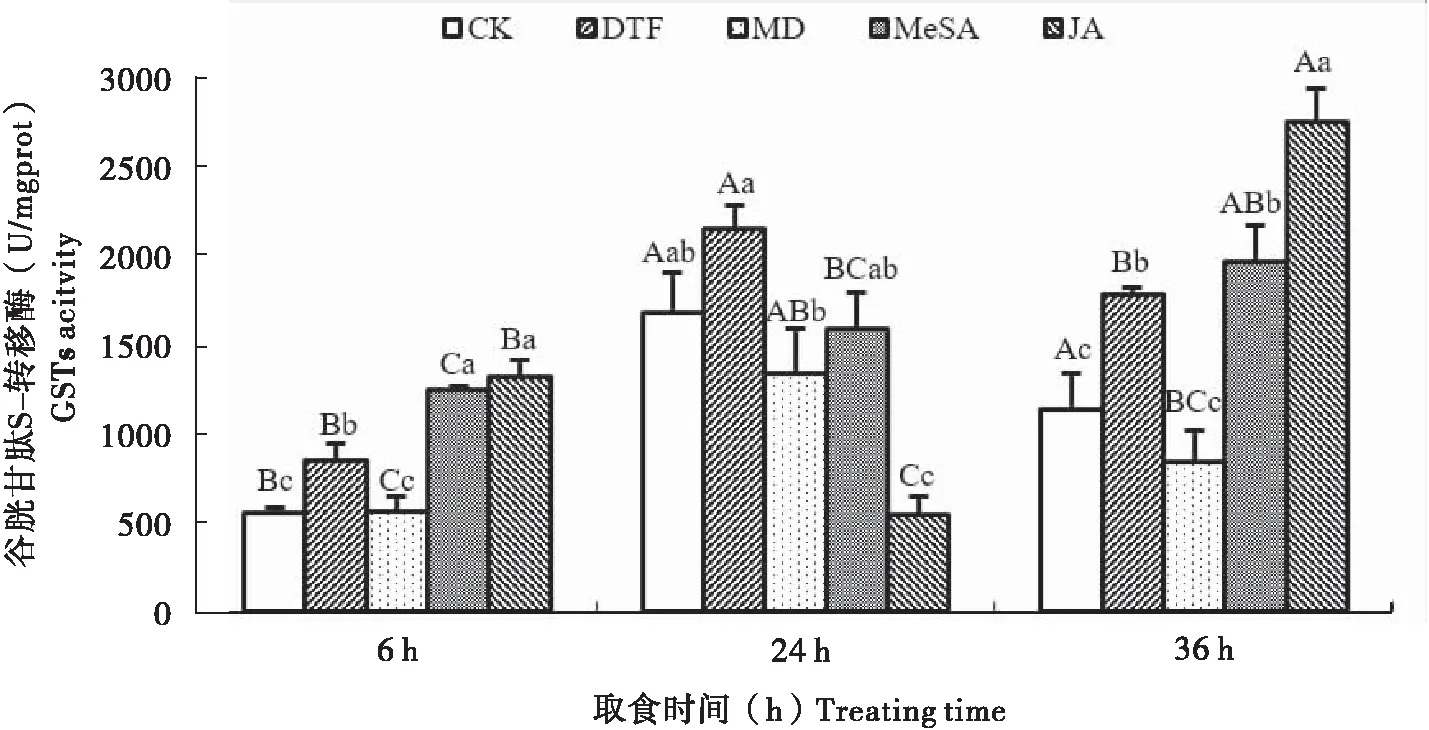
图3 不同处理诱导的菜豆植株对西花蓟马体内谷胱甘肽S-转移酶活性的影响Fig.3 Changes of GSTs activity in Frankliniella occidentalis feeding kidney bean induced by different treatments
2.4 羧酸酯酶(CarE)活性的变化
西花蓟马2龄若虫取食不同处理菜豆植株6 h时,取食蓟马危害和茉莉酸处理体内羧酸酯酶活性显著低于对照,分别为对照的59.16%和48.87%(图4),取食机械损伤、水杨酸甲酯与对照差异不显著。24 h时,取食茉莉酸处理虫体酶活性最高,为对照的2.12倍。取食蓟马危害与机械损伤处理的虫体酶活性均与对照差异不显著。取食水杨酸甲酯处理的虫体酶活性显著低于蓟马危害处理,但与对照差异不显著。36 h时,取食不同处理植株的虫体酶活性与对照相比均显著降低,分别只有对照的53.42%、65.22%、80.38%和69.45%。
西花蓟马取食机械损伤和水杨酸甲酯处理的菜豆其酶活性在不同取食时间下的变化趋势相似,均是24 h酶活性显著降低,随着取食时间的延长,36 h时酶活性又显著升高。取食蓟马危害和茉莉酸处理均是在24 h和36 h的酶活性显著高于6 h。

图4 不同处理诱导的菜豆植株对西花蓟马体内羧酸酯酶活性的影响Fig.4 Changes of CarE activity in Frankliniella occidentalis feeding kidney bean induced by different treatments
3 结论与讨论
昆虫面对植物产生的化学防御,将启动自身的解毒酶系统,将有毒物质转化成无毒或毒性较低的物质,从而免除外界不良条件的干扰。多功能氧化酶主要存在于昆虫消化道内细胞的内质网上,是一种广谱性的解毒代谢酶,外源物的进入可不同程度地诱导激活或抑制多功能氧化酶的活性(付昌斌和张兴,2000)。如菜青虫Pierisrapae取食川楝素后其幼虫体内微粒体多功能氧化酶活性明显被抑制(张兴和赵善欢,1992)。本研究发现,取食不同外源因子诱导处理菜豆植株24 h和36 h后,西花蓟马若虫体内的多功能氧化酶活性均显著受到抑制,且不同处理对西花蓟马多功能氧化酶的抑制效果不同,造成这种差异可能是因为不同外源因子诱导菜豆植株抗性产生的次生代谢物质种类或含量不同。
乙酰胆碱酯酶(TChE)作为有机磷和氨基甲酸酯类杀虫剂的作用靶标,在害虫抗药性形成过程中发挥着重要作用(曲明静等,2007)。有研究显示,次生物质对昆虫乙酰胆碱酯酶活性有影响且与取食时间存在一定的关联。刘少武和纪明山(2008)报道了辣椒碱在不同时间下对小菜蛾Plutellaxylostella体内的乙酰胆碱酯酶活性均有一定的抑制作用,与本研究取食不同外源因子诱导菜豆植株后,虫体乙酰胆碱酯酶活性均受到抑制一致,推测乙酰胆碱酯酶可能是西花蓟马适应不同诱导菜豆的作用位点之一。李春梅和郁建平(2013)发现血根碱对菜青虫乙酰胆碱酯酶具有强烈的抑制作用,其活性在36 h达到最低值,最大抑制率也高达95.12%,也推测乙酰胆碱酯酶可能是血根碱在菜青虫体内的作用靶标之一,本研究中蓟马取食机械损伤、水杨酸甲酯和茉莉酸处理诱导的菜豆植株,也发现在36 h虫体乙酰胆碱酯酶活性最低。双斑萤叶甲Monoleptahieroglyphica取食茉莉酸处理诱导的重瓣玫瑰后乙酰胆碱酯酶只是在1 d时具有显著抑制作用,取食水杨酸处理的乙酰胆碱酯酶活性在不同取食时间均与对照差异不显著(张鑫乾等,2014)。以上结果也表明不同的诱导处理对害虫体内乙酰胆碱酯酶活性抑制的时间效应不同。
谷胱甘肽S-转移酶是催化还原型谷胱甘肽与各种亲电子化合物进行亲核加成反应的一类酶能够使亲电化合物的亲水性增加,使其易于排出体外(Koide & Schreiner, 1992)。在昆虫体内,谷胱甘肽S-转移酶参与了多种植物次生物质的代谢,在昆虫的解毒代谢中起着非常重要的作用。本研究结果中,西花蓟马体内谷胱甘肽S-转移酶活性在取食蓟马危害处理后在6 h、24 h和36 h下均高于对照,而取食水杨酸甲酯和茉莉酸诱导处理后在6 h和36 h被诱导,24 h被抑制,取食机械损伤处理的酶活性在整个测定时间内与对照差异不显著,这可能与不同处理诱导菜豆的防御物质成分及含量有关,也和西花蓟马反防御机制的时间效应有关。不同的植物次生物质对不同植食性昆虫体内谷胱甘肽S-转移酶活性影响也不同,如董钧锋等(2002)研究发现烟碱和辣椒素显著诱导了烟青虫Prodenialitura体内谷胱甘肽S-转移酶活性,而番茄苷对烟青虫体内的谷胱甘肽S-转移酶具有抑制作用,烟碱、番茄苷、辣椒素和棉酚对棉铃虫谷胱甘肽S-转移酶均无显著影响。本研究结果表明,蓟马取食不同处理的菜豆植株后谷胱甘肽S-转移酶活性在不同取食时间下的变化不同,可能是不同诱导因子对植物次生物质诱导存在差异,导致昆虫取食后产生不同的生理生化反应。
羧酸酯酶(CarE)是昆虫体内重要的解毒酶,同时也可以被不同寄主植物或植物次生性物质所诱导,如棉蚜取食不同品种棉花对其羧酸酯酶活性影响较大,取食高抗品种后,总的羧酸酯酶活性被诱导,并且酶活力与单宁含量有一定的关系(姜永幸和郭予元,1996);用2-十三烷酮熏蒸处理48 h,棉蚜羧酸酯酶的活性达到最大值(吕敏等,2012)。本研究发现在6 h和36 h,蓟马危害的处理较机械损伤处理对虫体羧酸酯酶活性的抑制作用明显,但茉莉酸和水杨酸甲酯诱导表现不一致,这可能是因为茉莉酸和水杨酸甲酯介导的信号途径在植物防御机制中的作用不同,西花蓟马取食可以诱导菜豆启动茉莉酸和水杨酸介导的信号途径,从而使菜豆产生诱导防御机制(从春蕾等, 2014);机械损伤处理是人为模仿害虫危害处理,但蓟马取食两种处理后体内酶活性变化趋势却有差异,这可能是因为昆虫取食过程中口腔中的分泌物作用在受伤位点对植物防御基因表达产生了一定影响(殷海娣等,2006)。水杨酸及其衍生物水杨酸甲酯不仅可以诱导植物对多种病害产生抗性,还能诱导植物对害虫产生抗性,但水杨酸信号转导途径源自苯丙烷类代谢途径,而茉莉酸信号传导途径是亚麻酸通过脂氧合酶(1ipoxygenase, LOX)、丙二烯氧化合成酶(allele oxide sysnthase,AOS)等一系列酶促反应,因而在诱导植物抗性上存在着一定的差异(Kessler and Baldwin, 2002;胡留成,2008)。有关外源因子诱导植物抗性对虫体羧酸酯酶活性的影响,他人也有相似的研究报道,冯春富等(2011)的研究结果显示舞毒蛾Lymantriadispa幼虫取食经害虫危害、茉莉酸甲酯处理的落叶松后,其体内的羧酸酯酶活性在1-5 d时受到明显抑制。张鑫乾等(2014)也发现双斑萤叶甲取食茉莉酸甲酯处理的重瓣玫瑰后,其体内的羧酸酯酶活性在1 d和3 d时均受到显著抑制。
昆虫体内不同酶活性的变化不仅与取食不同的植物或次生物质存在差异,还与其他因素有关,如不同有效成分单宁酸和不同时间处理下,杨小舟蛾Micromelalophatroglodyte体内谷胱甘肽S-转移酶活性变化不同(张秀波等,2009)、低剂量的槲皮素可以诱导烟粉虱Bemisiatabaci谷胱甘肽S-转移酶的活性,高剂量的槲皮素对谷胱甘肽S-转移酶没有诱导作用,甚至还有抑制作用(牟少飞等,2006)等。本研究结果中,在6 h、24 h和36 h诱导处理下,乙酰胆碱酯酶活性在24 h高于其他取食阶段,这可能是因为植物在受到不同的诱导后,激发植物的防御反应,产生有毒次生物质,并随取食时间延长植物抗性水平提高,诱导增强昆虫相应的反防御系统,诱导相应酶系的活性,当不同酶底物浓度升高,反而抑制了酶得活性(牟少飞等,2006;彭露等,2010)。24 h时,虫体多功能氧化酶和羧酸酯酶酶活性低于6 h和36 h的,这可能是由于昆虫在取食初期产生了应急性适应,通过改变靶标的敏感性来避免次生物质的危害,使得在一段时间内酶活性较低。在短期内植物的某种次生物质对昆虫解毒酶有诱导作用,如牟少飞等(2006)发现槲皮素短时间酶可诱导羧酸酯酶酶和谷胱甘肽S-转移酶活性增加,随着植物防御反应中其他次生物质的产生,酶活性发生了变化。不同虫体酶在不同诱导因子和不同取食时间下的复杂变化,可能还与植物次生物质的种类、有效成分浓度、昆虫体内营养状态和靶标的敏感性等因素有关。
本研究结果表明西花蓟马若虫取食不同外源因子诱导处理的菜豆植株后,其体内的解毒酶活性被不同程度的抑制或激活,干扰了蓟马对有毒次生物质的解毒代谢功能。本试验只是选取西花蓟马2龄若虫,且取食时间只是设置3个时间梯度,不同虫态及其它不同时间还有待进一步的验证。另外,昆虫在受到有毒物质危害时,其体内生理代谢的响应必定是多方面的或综合性的,不仅仅是解毒酶发挥相应的解毒作用,保护酶也会通过自身酶活性调节,起到一定保护作用。因而,植物诱导抗性对昆虫生理生化机理的影响还需进一步的探讨。
References)
Arimura G, Kost C, Boland W. Herbivore-induced, indirect plant defences [J].BiochimicaetBiophysicaActa(BBA)-MolecularandCellBiologyofLipids, 2005, 1734 (2): 91-111.
Chen RF. Studies on Host-adaptation ofFrankliniellaoccidentalis[D]. Fuzhou: Fujian Agriculture and Forestry Universary, 2011. [陈锐芬. 西花蓟马对寄主植物的适应特性研究[D]. 福州: 福建农林大学, 2011]
Cong CL, Zhi JR, Liao QR,etal. Effects of thrips feeding, mechanical wounding, and exogenous methyl salicylate and jasmonic acid on defense enzyme activities in kidney bean leaf [J].ActaEntomologicaSinica, 2014, 57 (5): 564-571. [从春蕾, 郅军锐, 廖启荣, 等. 蓟马取食、机械损伤以及外源水杨酸甲酯和茉莉酸对菜豆叶片防御酶活性的影响[J]. 昆虫学报, 2014, 57 (5): 564-571]
Cong CL, Zhi JR, Xie LF,etal. Effects ofFrankliniellaoccidentalisfeeding on the chlorophyll and nutrients in the leaves ofPhaseolusvulgaris[J].PlantProtection, 2013, 39 (2): 20-24. [从春蕾, 郅军锐, 谢路飞, 等. 西花蓟马为害对菜豆叶绿素及营养物质含量的影响[J]. 植物保护, 2013, 39 (2): 20-24]
Dong JF, Zhang JH, Wang CZ. Effects of plant allelochemicals on nutritional utilization and detoxication enzyme activities in twoHelicoverpaspecies [J].ActaEntomologicaSinica, 2002, 45 (3): 296-300. [董钧锋, 张继红, 王琛柱. 植物次生物质对烟青虫和棉铃虫食物利用及中肠解毒酶活性的影响[J]. 昆虫学报, 2002, 45 (3): 296-300]
Feng CF, Yan SC, Lu YF,etal. Effects of induced resistance ofLarixgmeliniion the activities of detoxifying enzymes inLymantriadispar[J].ScientiaSilvaeSinicae, 2011, 47 (8): 102-107. [冯春富, 严善春, 鲁艺芳, 等. 兴安落叶松诱导抗性对舞毒蛾幼虫解毒酶活性的影响[J]. 林业科学, 2011, 47 (8): 102-107]
Fu CB, Zhang X. Effects ofSabinavulgarisextracts on the activities of several enzymes inMythimnasepartaLarvae [J].JournalofPlantProtection, 2000, 27 (1): 75-78. [付昌斌, 张兴. 砂地柏提取物对粘虫幼虫体内几种酶系活性的影响[J]. 植物保护学报, 2000, 27 (1): 75-78]
Gui LY, Liu SS, Chen ZM. Plant resistance to insects induced by application of exogenous jasmonic acid and methyl jasmonate [J].ActaEntomologicaSinica, 2004, 47 (4): 507-514. [桂连友, 刘树生, 陈宗懋. 外源茉莉酸和茉莉酸甲酯诱导植物抗虫作用及其机理[J]. 昆虫学报, 2004, 47 (4): 507-514]
Hu HY. Effects of Tobacco Infested byBemisiatabaciB-biotype on Interspecific Competition and Detoxification Metabolism betweenB.tabaciandTrialeurodesvaporariorum[D]. Taian: Shandong Agricultural University, 2003. [胡海燕. B型烟粉虱诱导的烟草防御对B型烟粉虱和温室白粉虱种间竞争及解毒代谢的影响[D]. 泰安: 山东农业大学, 2013]
Hu LC, Cui W, Wang X,etal. Herbivore resistallce induced bySpodopteralitura(Fabricius)(Lepidoptera: Noctuidae)and its relation to the JA signaling pathway in Chinese cabbage (BrassicacampestrisL.)[J].ActaEntomologicaSinica, 2010, 53 (9): 1001-1008. [胡留成, 崔巍, 汪霞, 等. 斜纹夜蛾幼虫诱导的油菜抗虫性及其与茉莉酸信号途径的关系[J]. 昆虫学报, 2010, 53 (9): 1001-1008]
Jiang YX, Guo YY. Studies on carboxylesterase ofAphisgossypiion different cotton varieties [J].ActaGossypiiSinica, 1996, 8 (4): 215-218. [姜永幸, 郭予元. 不同棉花品种对棉蚜羧酸酯酶活性影响的研究[J]. 棉花学报, 1996, 8 (4): 215-218]
Kessler A, Baldwin IT. Plant responses to insect herbivory [J].AnnualReviewofPlantBiology, 2002, 53 (53): 299-328.
Koide RT, Schreiner RP. Regulation of the Vesicular-ArbuscularMycorrhizalsymbiosis[J].AnnualReviewofPlantPhysiologyandPlantMolecularBiology, 1992, 43 (1): 557-581.
Kostaropoulos I, Papadopoulos AI, Metaxakis A,etal. Glutathione-S-transferase in the defence against pyrethroids in insects [J].InsectBiochemistryandMolecularBiology, 2001, 31 (4): 313-319.
Li CM, Yu JP. Effect of sanguinarine on metabolic enzymes ofPierisrapae[J].ChineseJournalofBiologicalControl, 2013, 29 (3): 463-468. [李春梅, 郁建平. 血根碱对菜青虫几种代谢酶活性的影响[J]. 中国生物防治学报, 2013, 29 (3): 463-468]
Liu JY, Qian L, Jiang XC,etal. Effects of elevated CO2concentration on the activities of detoxifying enzymes and protective enzymes in adults ofFrankliniellaoccidentalisandF.intonsa(Thysanoptera: Thripidae)[J].ActaEntomologicaSinica, 2014, 57 (7): 754-761. [刘建业, 钱蕾, 蒋兴川, 等. CO2浓度升高对西花蓟马和花蓟马成虫体内解毒酶和保护酶活性的影响[J]. 昆虫学报, 2014, 57 (7): 754-761]
Liu SW, Ji MS. Effects of capsaicin on the activities of acetylcholinesterase and carboxylesterase inPlutellaxylostella[J].ChinaPlantProtection, 2008, 28 (4): 8-9, 18. [刘少武, 纪明山. 辣椒碱对小菜蛾体内乙酰胆碱酯酶和羧酸酯酶的影响[J]. 中国植保导刊, 2008, 28 (4): 8-9, 18]
Lv M, Sun HH, Wang LH,etal. Effects of secondary metabolites on activities of glutathione S-transferases, carboxylesterase in aphid [J].ChineseAgriculturalScienceBulletin, 2012, 28 (3): 253-256. [吕敏, 孙婳婳, 王丽红, 等. 植物次生物质对棉蚜谷胱甘肽S-转移酶和羧酸酯酶活性的诱导作用[J]. 中国农学通报, 2012, 28 (3): 253-256]
Lv YB, Zhang ZJ, Wu QJ,etal. Research progress of the monitoring, forecast and sustainable management of invasive alien pestFrankliniellaoccidentalisin China [J].ChineseJournalofAppliedEntomology, 2011, 48 (3): 488-496. [吕要斌, 张治军, 吴青君, 等. 外来入侵害虫西花蓟马防控技术研究与示范[J]. 应用昆虫学报, 2011, 48 (3): 488-496]
Mou F, Zhi JR, Cong CL,etal. Effects of exogenous jasmonic acid and methyl salicylate on developmental duration and population dynamics ofFrankliniellaoccidentalis[J].PlantProtection, 2014, 40 (6): 70-74. [牟峰, 郅军锐, 从春蕾, 等. 外源茉莉酸和水杨酸甲酯对西花蓟马生长发育和种群动态的影响[J]. 植物保护, 2014, 40 (6): 70-74]
Mu SF, Liang P, Gao XW. Effects of quercetin on specific activity of carboxylesterase and glutathione S-transferases inBemisiatabaci[J].ChineseBulletinofEntomology, 2006, 43 (4): 491-495. [牟少飞, 梁沛, 高希武. 槲皮素对B型烟粉虱羧酸酯酶和谷胱甘肽S-转移酶活性的影响[J]. 昆虫知识, 2006, 43 (4): 491-495]
Peng L, Yan Y, Liu WX,etal. Counter-defense mechanisms of phytophagous insects towards plant defense [J].ActaEntomologicaSinica, 2010, 53 (5): 572-580. [彭露, 严盈, 刘万学, 等. 植食性昆虫对植物的反防御机制[J]. 昆虫学报, 2010, 53 (5): 572-580]
Qiu LH, Zhang WJ. Relationship between mixed-function oxidases and the resistance to fenvalerate inHelicoverpaarmigera[J].ActaEntomologicaSinica, 2001, 44 (4): 447-453. [邱立红, 张文吉. 微粒体多功能氧化酶系与棉铃虫对氰戊菊酯抗药性的关系[J]. 昆虫学报, 2001, 44 (4): 447-453]
Qu MJ, Xu XJ, Han ZJ,etal. Advances in studies of acetylcholinesterase gene variation assoiated with insect resistance [J].ChineseBulletinofEntomology, 2007, 44 (2): 191-194. [曲明静, 许新军, 韩召军, 等. 昆虫乙酰胆碱酯酶基因变异抗药性机制研究[J]. 昆虫知识, 2007, 44 (2): 191-194]
Robertson JL, Armstrong KF, Suckling DM,etal. Effects of host plants on the toxicity of azinphosmethyl to susceptible and resistant light brown apple moth (Lepidoptera: Tortricidae)[J].JournalofEconomicEntomology, 1990, 83 (6): 2124-2129.
Song YQ, Sun HZ, Li T,etal. Effect of the host plants on activity of 4 detoxifying enzymes in beet armyworm,Spodopteraexigua[J].ChineseAgriculturalScienceBulletin, 2009, 25 (19): 203-205. [宋月芹, 孙会忠, 李涛, 等. 不同寄主植物对甜菜夜蛾解毒酶活性的影响[J]. 中国农学通报, 2009, 25 (19): 203-205]
Tian XT, Li Dan, Zhang J,etal. Inductive effects of exogenous jasmonic acid on three defensive enzymes in leaves of kidney bean and tomato seedlings [J].ActaAgriculturaeBoreali-occidentalisSinica, 2013, 22 (9): 136-140. [田旭涛, 李丹, 张箭, 等. 外源茉莉酸对菜豆和番茄幼苗3种防御酶的诱导效应[J]. 西北农业学报, 2013, 22 (9): 136-140]
Wu GZ, Hu L, Ye M,etal. Effects of soybean trypsinase inhibitor and defense signaling compounds on detoxification enzymes inSpodopteralitura(F.)larvae [J].ChineseJournalofAppliedEcology, 2012, 23 (7): 1952-1958. [吴国昭, 胡林, 叶茂, 等. 大豆胰蛋白酶抑制剂和防御信号物质对斜纹夜蛾解毒酶的影响[J]. 应用生态学报, 2012, 23 (7): 1952-1958]
Yang F. The Resistance of Cucumber Induced by Exogenous Salicylicacid and Methyl Jasmonate Respectively toFrankliniellaoccidentalis(Pergande)(Thysanoptera, Thripidae)[D]. Wuhan: Huazhong Agricultural University, 2009. [杨帆. 水杨酸、茉莉酸甲酯诱导黄瓜对西花蓟马(缨翅目:蓟马科)的抗性[D]. 武汉: 华中农业大学, 2009]
Yin HD, Huang CH, Xue K,etal. Roles of insect salivary components in insect-plant interactions [J].ActaEntomologicaSinica, 2006, 49 (5): 843-849. [殷海娣, 黄翠虹, 薛堃, 等. 昆虫唾液成分在昆虫与植物关系中的作用[J]. 昆虫学报, 2006, 49 (5): 843-849]
Yu SJ. Consequences of Induction of Foreign Compound-metabolizing Enzymes in Insects Molecular Aspects of Insect-plant Associations [M]. New York: Springer US, 1986: 153-174.
Zhang X, Zhao SH. Effects of toosndanin on the activities of several enzymes in the imported cabbage wormPierisrapaeL. [J].ActaEntomologicaSinica, 1992, 35 (2): 171-177. [张兴, 赵善欢. 川楝素对菜青虫体内几种酶系活性的影响[J]. 昆虫学报, 1992, 35 (2): 171-177]
Zhang XB, Tang F, Liu YS,etal. Induction of glutathione S-tranderases by tannic acid inMicromelalophatroglodyta[J].ChineseBulletinofEntomology, 2009, 46 (4): 579-584. [张秀波, 汤方, 刘玉升, 等. 单宁酸对杨小舟蛾谷胱甘肽S-转移酶活性的诱导[J]. 昆虫知识, 2009, 46 (4): 579-584]
Zhang XQ, Yan JX, Yang JY,etal. Effects of induced resistance ofRosarugosa‘plena’ on the activities of detoxifying enzymes inMonoleptahieroglyphica(Motschulsky)[J].JournalofNortheastForestryUniversity, 2014, 5: 125-128, 148. [张鑫乾, 严俊鑫, 杨杰莹, 等. 重瓣玫瑰诱导抗性对双斑萤叶甲成虫解毒酶的影响[J]. 东北林业大学学报, 2014, 5: 125-128,148]
EffectofdifferenttreatmentsofkidneybeanontheactivitiesofdetoxificationenzymesinFrankliniellaoccidentalis
MU Feng, ZHI Jun-Rui*, YUE Wen-Bo, ZENG Guang
(Institute of Entomology, Guizhou University; Guizhou Provincial Key Laboratory for Agricultural Pest Management of the Mountainous Region, Guiyang 550025, China)
To assess the influence of induced resistance of plants on the physiological and biochemical substances inFrankliniellaoccidentalis. The kidney beans were treated by thrips damage, mechanical damage, exogenous jasmonic acid (JA)and methyl salicylate (MeSA)induction respectively. The changes of activities of multi-function oxidase (MFO), carboxylesterase (CarE), glutathione S-transferase (GSTs), cholinesterase (TChE)in 2ndlarvae ofF.occidentalisfeeding different induced kidney bean were evaluated. The results showed that the activities of MFO and TChE in 2ndlarvae had similar results, they were significantly lower than that of feeding healthy bean at 24 h and 36 h afterF.occidentalisfeeding kidney bean leaves of different treatments. The GSTs activity inF.occidentalisincreased only at 24 h after thrips feeding kidney bean induced by JA, and the activities of GSTs induced by thrips damage, exogenous MeSA induction were higher than control respectively. GSTs activity inF.occidentalisfeeding mechanical damaged plants wasn’t significantly different compared with the control under the different feeding time. The CarE activity was significantly inhibited at 6 h and 36 h after feeding the kidney bean plant by thrips damage and JA induced, while it was significantly lower only at 36 h than the control at mechanical damage and MeSA induction. The above results indicated that the induced resistance by exogenous factors interfered withF.occidentalisdetoxification mechanisms in the body and reduced its detoxification metabolic function of toxic secondary substance.
Frankliniellaoccidentalis; kidney bean; induced resistance; detoxifying enzymes
国家自然科学基金(31660516);贵州省农业攻关项目(黔科合NY[2015]3014-1号);贵州省国际科技合作基地(黔科合平台人才[2016]5802)
牟峰,男,1989年生,吉林桦甸人,硕士研究生,研究方向为农业昆虫与害虫防治,E-mail: 15285138563@163.com
*通讯作者Author for correspondence,E-mail: zhijunrui@126.com
Received: 2016-08-19;接受日期Accepted: 2016-11-06
Q965;S433.89
:A
1674-0858(2017)04-0862-08
牟峰,郅军锐,岳文波,等.不同处理菜豆植株对西花蓟马体内解毒酶活性的影响[J].环境昆虫学报,2017,39(4):862-869.
