淮河上游南湾湖麦穗鱼年龄与生长的研究
2017-08-16李红敬王雅平冷秋丽李晓杰李晓凤于同雷
李红敬 王雅平 冷秋丽 李晓杰 李晓凤 于同雷 黄 斌
(信阳师范学院生命科学学院, 信阳 464000)
淮河上游南湾湖麦穗鱼年龄与生长的研究
李红敬 王雅平 冷秋丽 李晓杰 李晓凤 于同雷 黄 斌
(信阳师范学院生命科学学院, 信阳 464000)
2014年的3月、5月、7月和12月在淮河上游南湾湖采集麦穗鱼(Pseudorasbora parva)样本532尾, 对麦穗鱼的年龄组成与生长进行分析。结果表明样本的体长分布范围为35.82—88.28 mm, 平均体长为(61.61±11.8) mm,体重的分布范围为3.07—59.17 g, 平均体重为(19.23±10.73) g。雄性个体比雌性个体大, 雌雄性比为0.64鲶1。群体的年龄组成为1—3龄, 其中3龄样本数量占优势为57.38%。体长与体重的关系是雌性W=9.602E–5L2.928(R2=0.883); 雄性W=4.487E–5L3.116(R2=0.889), 雌雄样本间存在显著性差异(F=5.241, P<0.05)。麦穗鱼的鳞径与体长之间呈线性关系, 并且雌雄样本的鳞径与体长之间的关系差异性显著(F=78.405, P<0.05)。生长参数分别是雌性: L∞=107.005, K=0.246, t0= –0.76; 雄性: L∞=145.254, K=0.181, t0= –0.66。生长拐点是雌性3.607龄对应的体长和体重分别为70.46 mm和24.72 g, 雄性5.619龄对应的体长和体重分别为98.64 mm和73.53 g。研究结果表明雌性为匀速生长, 雄性为异速生长; 雄性麦穗鱼比雌性麦穗鱼的生长速度快。
南湾湖; 麦穗鱼; 年龄; 生长
麦穗鱼(Pseudorasbora parva)隶属于鲤形目(Cypriniformes), 鲤科(Cyprinidae), 逗亚科(Gobioninae), 麦穗鱼属(Pseudorasbora)。栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等水域[1,2], 对不同栖息环境有着较强的适应性; 食杂性[3], 以浮游动物、底栖动物为主要食物[4]。世代周期短、性成熟早、繁殖力强[5]。属于东亚地区的土著物种, 广泛分布于中国大陆[6]、日本[7]、朝鲜[8]、中国台湾[9]等国家和地区。20世纪50年代从黑龙江流域入侵至苏联[10], 此后快速的入侵至欧洲的其他国家, 目前其分布已遍及全球[11]。
在东亚地区分布的麦穗鱼在生态系统中占有重要地位, 不仅能为肉食性鱼类提供食物资源, 而且能够维持浮游动植物种群的稳定[4,12], 而在其他入侵地, 麦穗鱼的入侵给当地鱼类生态带来了巨大影响[13], 甚至威胁到一些土著种的生存, 如云南的滇池、洱海和抚仙湖就有多种土著鱼类因麦穗鱼等入侵种而日趋减少或濒临灭绝[14]。在英国湖区麦穗鱼在入侵后4年繁殖成为当地数量最多的鱼类,而威胁到其他本土鱼类的生存[15]。对入侵麦穗鱼的管理和控制是目前亟待解决的问题。
鱼类年龄与生长的研究, 是分析和评价鱼类种群数量变动的基本依据之一。目前在亚洲地区对于麦穗鱼生物学方面的研究主要集中于体长与体重关系[16]以及生长参数的比较[17]等方面, 对于麦穗鱼生长规律等方面的研究还较为缺乏。而在麦穗鱼的入侵地对其生物学方面的研究, 主要是通过比较种群分布以及个体生长的变化情况来确定环境对其生长的影响[18—21]。目前对于入侵麦穗鱼的治理主要是采取生物防治[22]。因此, 我们以分布于东亚大陆淮河上游区域的南湾湖及其周边溪流中的麦穗鱼为研究对象, 对麦穗鱼种群的生长特性开展研究。旨在了解亚热带向暖温带过渡区麦穗鱼的自然生长规律, 为入侵麦穗鱼的防治和管理以及麦穗鱼的应用研究提供参考资料, 对淮河流域鱼类资源的资料进行补充。
1 材料与方法
1.1 样本采集
南湾湖(32°08′N, 113°58′E)位于淮河上游, 受东亚季风气候的影响, 具有北亚热带向暖温带过渡的季风气候和山地气候特征, 与同纬度的其他地区相比, 冬夏气温均偏低, 年均温15.2℃, 年均降雨量约1140 mm, 年平均蒸发量约935 m3。
2014年3月、5月、7月、10月和12月, 在信阳市南湾湖的三个较大入水河口附近(谭家河、董家河以及浉河港), 使用成套刺网在采样区内对鱼类进行随机采样。每套刺网由8种网目规格各异的浮性刺网(网目大小依次为20、25、33、40、50、60、70和80 mm, 均高1.5 m、长20 m) 串联而成, 用以捕获较大体长范围内的鱼类个体。日落之前, 在采样区下两套共16条刺网, 刺网上端浮子浮力大于底端铅锤质量; 第2天早晨天亮后收网, 上岸摘鱼,在整个采样期间, 这两套刺网按同样采样程序用于各个采样点, 以保障渔获物数据之间的可比性。样本采集后在新鲜状态下运送至实验室, 用纱布和吸水纸将样本表面多余的水分擦拭干净, 使每尾样本在测量时都保持一样的状态, 分别进行编号, 用游标卡尺测量体长和全长(精确至0.01 mm)。使用FA2104A型电子天平称量体重(精确至0.01 g)。取每尾样本的背鳍起点下方至侧线上方的鳞片5—10枚, 用于年龄鉴定。
1.2 数据处理
体长与体重的关系采用Keys公式W=aLb对雌雄样本分别进行拟合。其中W表示体重(g)、L表示体长(mm)、a是常数、b是幂指数。以体长的自然对数值作为协变量进行方差分析, 检验雌、雄个体间体长与体重的关系是否存在差异。体长体重方程中指数b值能反映鱼类在不同发育阶段和不同生境中的生长发育情况, 当鱼类体长体重方程的b值等于3或接近3时, 体长与体重的生长为等速生长。
体长与鳞径的关系采用不同的鳞径与体长的关系方程, 分别为: Einar Lea、Rosa Lee和蒙纳斯蒂尔斯基方程Ln=Lrn/R、L=a+bR、以及L=aRb对麦穗鱼体长与鳞径的关系进行拟合, 选择判定系数(R2)最大者为其最佳回归方程; 采用Rosa Lee方程对雌雄样本的体长进行退算。协方差分析雌雄样本的鳞径与体长的关系。
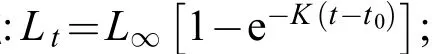
数据的统计整理使用Microsoft excel2007, 实验结果的分析和图表的制作使用Statistica 8.0。
2 结果
2.1 样本组成与性比
使用鳞片能够成功鉴定出427尾麦穗鱼样本的年龄, 其余105尾样本由于鳞片破损或者其他原因未能成功鉴定出年龄。对427尾麦穗鱼样本的年龄进行统计。其中1龄样本较少, 占总样本的6.32%, 2龄样本占36.3%, 3龄样本较多占57.38%。
通过对427尾麦穗鱼体长、体重的测量和统计(图 1、图 2), 其中雌性167尾, 雄性260尾。性比为0.64鲶1, 用χ2分析比较性比与1鲶1之间存在显著性差异(P<0.05)。 427尾麦穗鱼总样本的体长范围为35.82—88.28 mm, 平均体长为(61.61±11.8) mm。雌性样本的体长为35.82—87.76 mm, 平均体长为(55.99±10.52) mm。雄性样本的体长为36.83—88.28 mm, 平均体长为(65.22±11.17) mm。雌性样本和雄性样本的体长分布存在显著性差异(Kolmogorov-Smirnov检验D=4.635, P<0.05)。
总样本的体重范围为3.07—59.17 g, 平均体重为(19.23±10.73) g。雌性样本的体重范围为3.07—49.54 g, 平均体重为(14.08±8.06) g。雄性样本的体重范围为3.08—59.17 g, 平均体重为(22.54±10.94) g。雌性样本和雄性样本的体重分布存在显著性差异(Kolmogorov-smirnov检验D=4.597, P<0.05)。
2.2 体长与体重的关系

图 1 麦穗鱼总样本群体及雌、雄样本体长组成Fig.1 The distribution patter of body length of the total samples, male and female Pseudorasbora parva
对麦穗鱼的体长和体重的关系进行拟合(图 3),其相关式为: 雌性W=9.602E–5L2.928R2=0.883; 雄性W=4.487E–5L3.116R2=0.889; 将雌雄样本的体长、体重进行对数处理, 使其成线性关系, 用协方差分析(ANCOVA)检验雌雄之间体长、体重关系存在显著性差异(F=5.241, P<0.05)。分别将雌雄样本以及总样本的长重关系指数与匀速生长鱼类的b=3进行比较, 雌性(b=2.928)与3之间无显著性差异(t=–1.787, P>0.05), 雄性(b=3.116)与3之间存在显著性差异(t=9.278, P<0.05), 即雌性为匀速生长, 雄性为异速生长。可以用Von Bertalanffy生长方程(VBGF)来描述其生长特性。
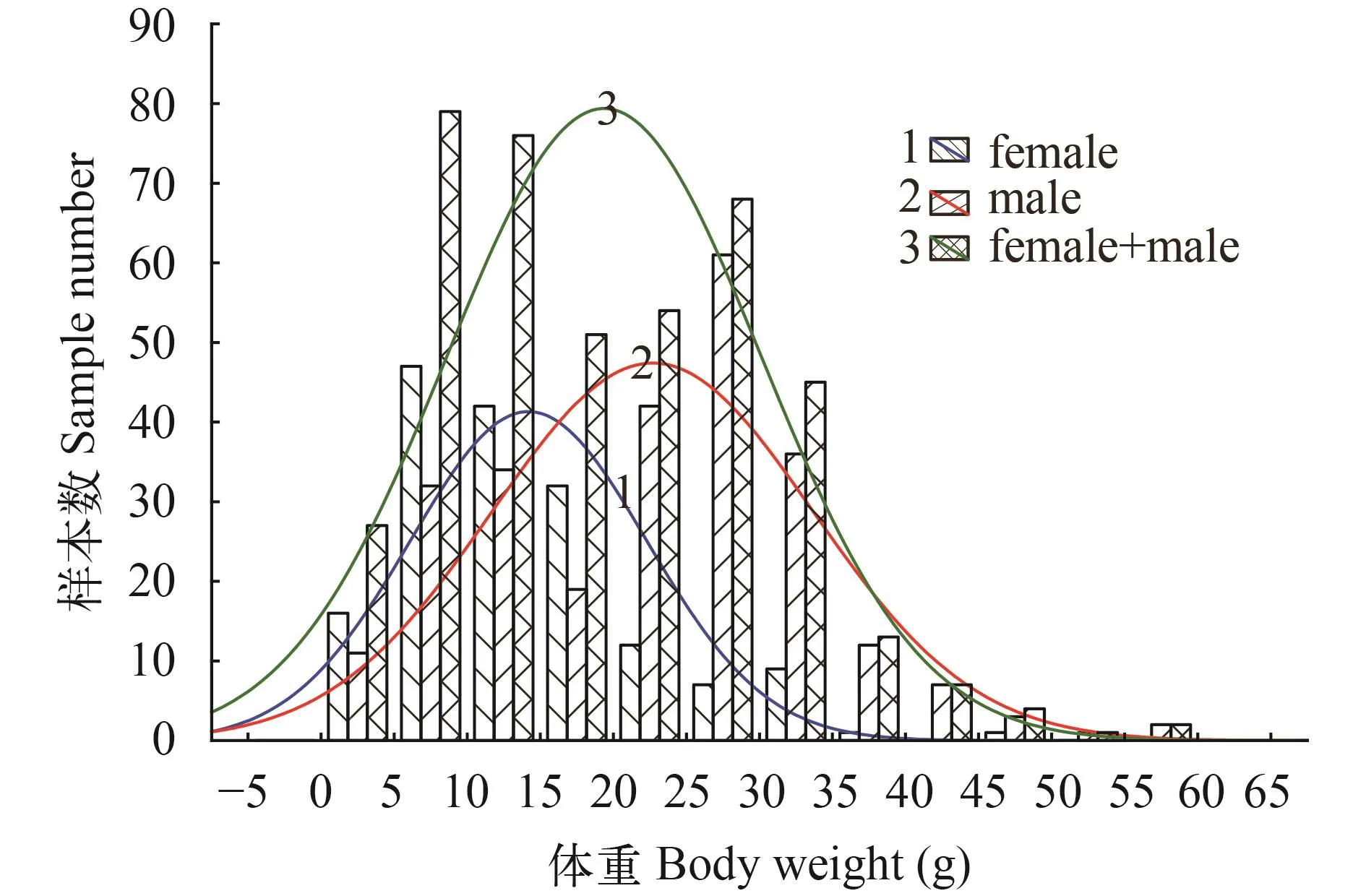
图 2 麦穗鱼总样本群体及雌、雄样本体重组成Fig.2 The distribution pattern of body weight of the total samples, male and female Pseudorasbora parva

图 3 麦穗鱼体长与体重的关系Fig.3 The relationship between body length and body weight of Pseudorasbora parva
2.3 体长与鳞径的关系
采用Einar Lea、Rosa Lee和蒙纳斯蒂尔斯基这3个方程对鳞径与体长的关系进行拟合, 并根据拟合出的方程退算出雌雄样本的体长(表 1)。通过对3种方法的退算体长与实测体长的比较, 结果显示Rosa Lee公式退算出的体长值与实测体长的差异性较小, 因此确定麦穗鱼样本的鳞径与体长的关系式为, 雌性: L=16.399+0.053R; 雄性: L=23.501+ 0.0414R。
协方差分析显示雌雄样本的鳞径与体长的关系存在显著性差异(ANCOVA分析F=78.405, P<0.05)。
2.4 体长的退算
根据鳞径与体长的关系, 对麦穗鱼雌雄样本各龄的体长进行退算(表 2)。对雌雄实测体长与退算体长分别进行χ2检验结果显示, 雌性P>0.05 (χ2=0.079, df=2, P=0.961), 雄性P>0.05 (χ2=0.108, df=2, P=0.948), 即雌雄样本的实测体长与退算体长之间不存在显著性差异。表明选用Rosa Lee公式作为鳞径与体长的关系式是可靠的。
2.5 生长方程
根据平均退算体长以及体长与体重的关系, 计算出各龄的退算体重。根据ford方程求得雌雄群体的生长参数, 根据Beverton法从ln(L∞–Lt)=lnL∞+ Kt0–Kt求得理论体长或体重等于0时的年龄t0。各生长参数如下, 雌性: L∞=107.005, W∞=84.034, K=0.246, t0=–0.76; 雄性: L∞=145.254, W∞=244.989, K=0.181, t0=–0.66。将计算出的生长参数代入Von Bertalanffy生长方程, 得出麦穗鱼雌雄群体体长、体重的生长方程分别为, 雌性: Lt=107.005[1–e–0.246(t+0.76)], Wt=84.034[1–e–0.246(t+0.76)]2.928; 雄性: Lt=145.254 [1–e–0.181(t+0.66)] , Wt=244.989[1–e–0.181(t+0.66)]3.116
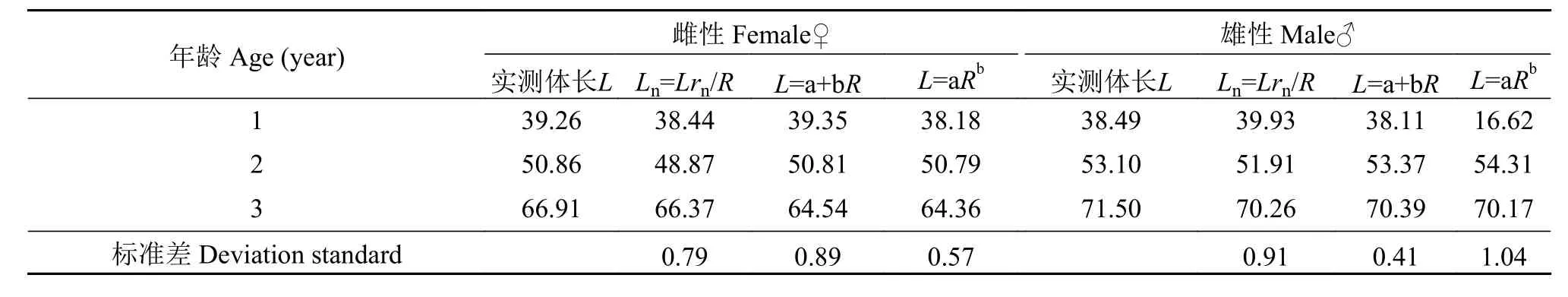
表 1 不同方法退算麦穗鱼体长的比较Tab.1 The comparison of backed-calculated length of different methods to Pseudorasbora parva
F检验结果显示雌雄的体长生长方程回归极显著(P<0.01)。将理论体长与退算体长进行χ2检验,结果显示雌性P>0.05, 雄性P>0.05, 差异性不显著,说明体长的生长曲线拟合程度可靠。从生长曲线(图 4)可以看出, 雌雄群体体长的生长都不具拐点,开始生长很快, 但是随着年龄的增长, 其生长逐渐接近于渐进值, 而体重的生长则近似于“S”形生长。

表 2 麦穗鱼各龄退算体长和平均实测体长Tab.2 Backed-calculated Length (BL) and mean observed BL of Pseudorasbora parva
分别对体长和体重生长方程求一阶导数, 即得出其生长速度的方程:
雌性: dL/dt=26.323e–0.246(t+0.76); dW/dt=60.529e–0.246(t+0.76)[1–e–0.246(t+0.76)]1.928
雄性: dL/dt=26.291e–0.181(t+0.66); dW/dt=138.173e–0.181(t+0.66)[1–e–0.181(t+0.66)]2.116
对生长方程求二阶导数可得其生长加速度的方程:
雌性: d2L/dt2= –6.476e–0.246(t+0.76)
d2W/dt2=14.89e–0.246(t+0.76)[1–e–0.246(t+0.76)]0.928[2.928e–0.246(t+0.76)–1]
雄性: d2L/dt2= –4.759e–0.181(t+0.66)
d2W/dt2=25.009e–0.181(t+0.66)[1–e–0.181(t+0.66)]1.116[3.116e–0.181(t+0.66)–1]
体长和体重的生长速度和加速度曲线如图 5—8所示。计算出麦穗鱼生长的拐点年龄分别为雌性3.607龄对应的体长和体重分别为70.46 mm和24.72 g,雄性5.619龄对应的体长和体重分别为98.64 mm和73.53 g。

图 4 麦穗鱼体长、体重生长曲线Fig.4 The growth curve of Pseudorasbora parva
3 讨论

图 5 雄性体长生长速度与加速度曲线Fig.5 Growth in body length as dL/dt and d2L/dt2of male Pseudorasbora parva
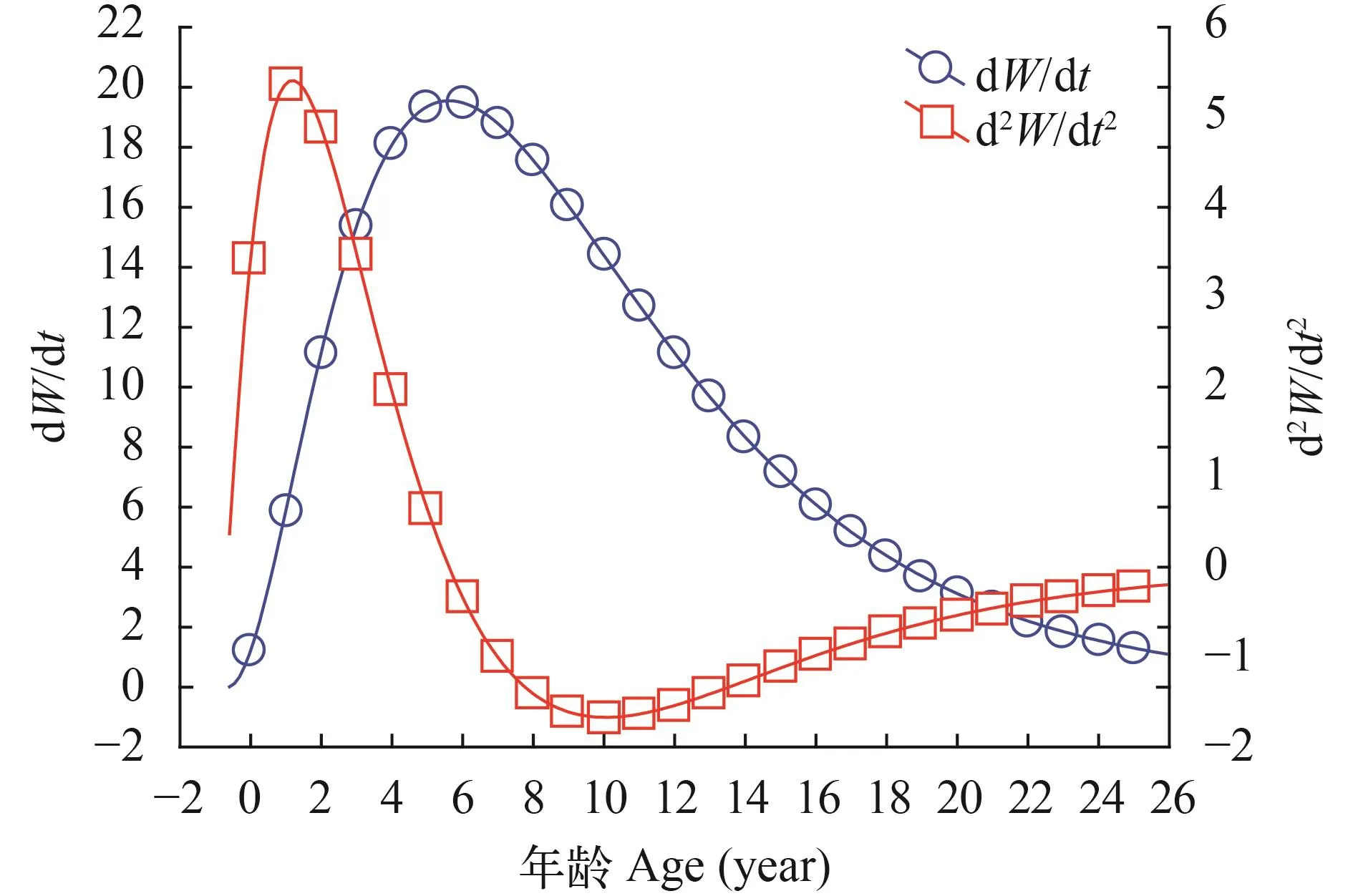
图 6 雄性体重生长速度与加速度方程Fig.6 Growth in body length as dW/dt and d2W/dt2cruves of male Pseudorasbora parva
目前, 对麦穗鱼的年龄鉴定, 通常使用长度频度法[17]和鳞片鉴定法[16,19]。我们选取鳞片作为麦穗鱼的年龄鉴定材料, 主要是因为麦穗鱼的鳞片易处理轮纹清晰。并且多数小型鲤科鱼类都用鳞片鉴定其种群的年龄[23,24]。
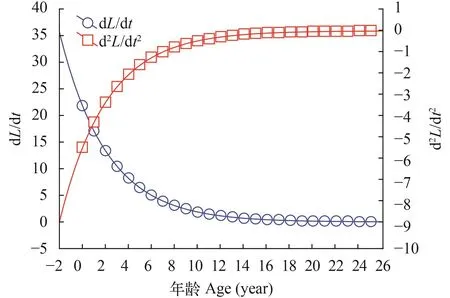
图 7 雌性体长生长速度与加速度Fig.7 Growth in body weight as dW/dt and d2W/dt2cruves of female Pseudorasbora parva
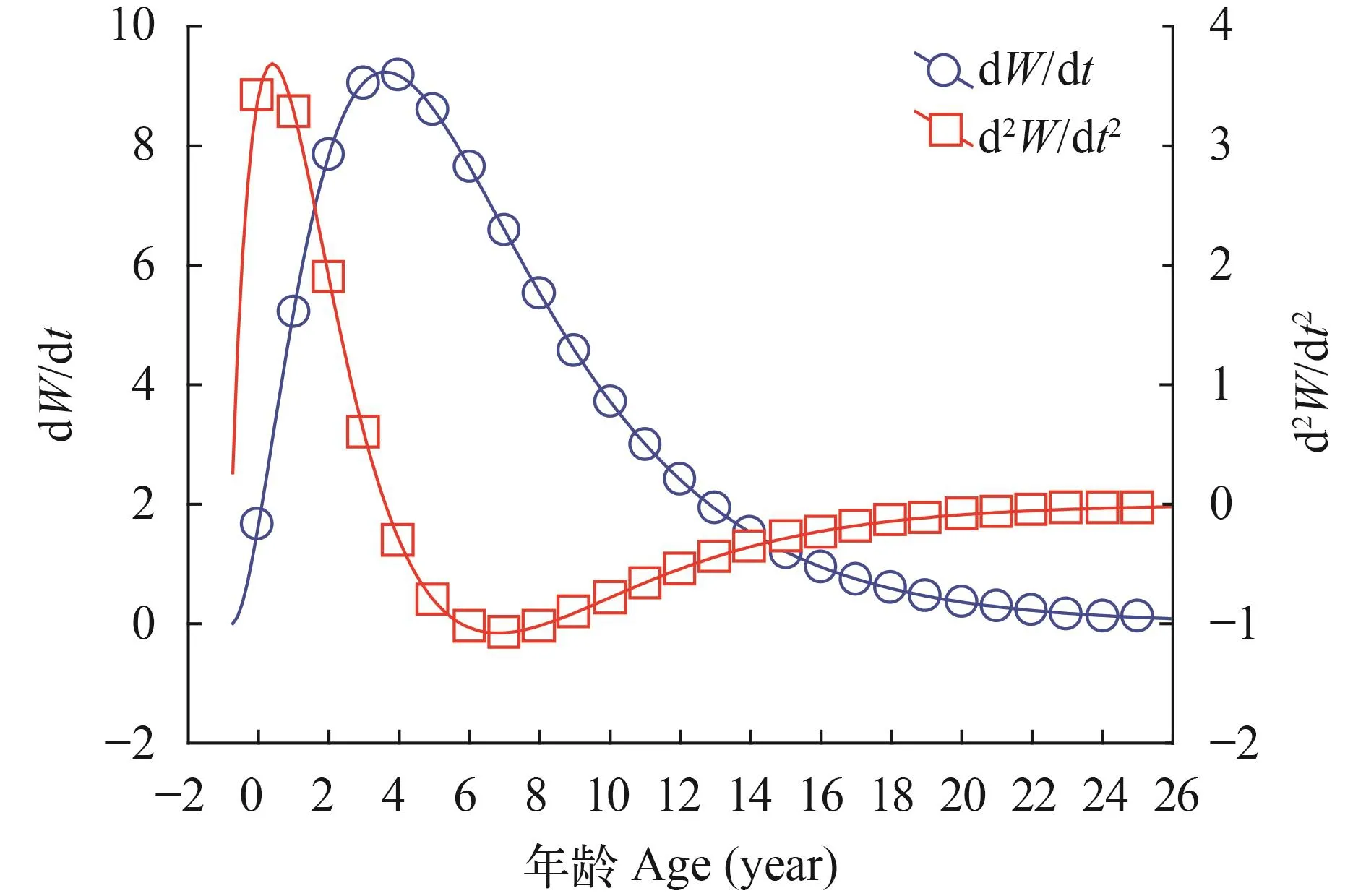
图 8 雌性体重生长速度与加速度Fig.8 Growth in body weight as dW/dt and d2W/dt2of female Pseudorasbora parva
鉴定结果显示南湾湖麦穗鱼的最大年龄为3龄,最大体长是88.28 mm, 小于保安湖麦穗鱼的最大体长100.5 mm[16]。大于日本九州岛麦穗鱼的最大体长60 mm[17]。这种差异出现的原因主要是环境不同导致的。九州岛麦穗鱼样本采集自有明海下游的一条人工灌溉沟渠, 沟渠中水生生物数量较少,不能为鱼类的生长提供足够的食物, 从而影响其生长。而保安湖纬度较低, 水体温度较高更加适合鱼类的生长。而在入侵地布拉迪斯拉发的麦穗鱼最大年龄为5龄[19], 普雷斯帕湖为4龄[18]。在英国2个不同池塘中, 麦穗鱼的最大体长分别是76和81 mm[20]。在入侵地麦穗鱼的寿命有所延长, 体长相对较小,说明麦穗鱼对环境的适应能力强, 入侵后并未对种群的生长造成影响。
研究结果显示麦穗鱼的雌雄群体的生长方式存在差异, 雌性群体为匀速生长(b=2.928);雄性群体为异速生长(b=3.116)。张堂林等[16]研究保安湖麦穗鱼的体长与体重的关系, 雄性麦穗鱼样本在4月、5月和10月属于匀速生长方式, 其他月份则表现为异速生长的特征; 雌性麦穗鱼样本在4月、5月表现为匀速生长, 其他月份表现为异速生长的特征。而其他地区的麦穗鱼种群的生长表现为异速生长方式[18]。这主要是因为鱼类体长与体重的关系会受到性别、性腺发育以及季节的影响[25]。
我们确定麦穗鱼体长与鳞径的关系是通过比较三种不同的鳞径与体长的关系而筛选出来的最适合的方法。Einar Lea公式认为鳞片与体长之间呈正比, Rosa Lee公式认为鳞片与体长呈线性关系,并且多数鱼类的鳞径与体长都符合这一关系[26,27]。也有鱼类的鳞径与体长呈曲线关系[28]我们通过比较不同方法对体长的退算, 从而确定麦穗鱼的鳞径与体长之间呈线性关系, 并根据这一关系对雌雄群体的体长进行了退算, 退算结果略小于实测体长,出现轻微的李氏现象。但是退算值与实测值之间并不存在显著性差异(P>0.05)说明用线性关系拟合麦穗鱼鳞径与体长的关系是可取的, 而其他地区麦穗鱼的研究结果也证实了其鳞径与体长之间是线性关系[16,19]。
鱼类的生长通常会有一个适合其生长参数的数学模型来分析其生长趋势[29]。von Bertalanffy生长方程是一个主要用来描述年龄与体长之间非线性关系的鱼类生长模型[30]。我们通过von Bertalanffy生长方程计算出麦穗鱼的渐进体长雄性145.254 mm大于雌性107.005 mm, 但是都要大于所采集麦穗鱼样本的最大实测体长(雄性88.28 mm,雌性87.76 mm), 说明本研究对于麦穗鱼的生长参数的估算是可靠的。K值表示鱼类个体的生长接近渐进值的速度, 在本研究中雌性麦穗鱼的K值(0.246)大于雄性(0.181), 即雌性个体生长到渐进体长的速度比雄性快, 也就符合了雌性的渐进体长小于雄性的现象。将本研究的麦穗鱼的各项生长指标与其他地区的麦穗鱼的生长指标进行比较, 俄罗斯阿穆尔河麦穗鱼的K值0.41相对较大, 渐进体长是109 mm较小[31], 说明其生长到接近于渐进体长的速度更快, 这可能与俄罗斯的气候有关, 俄罗斯地处北温带, 气候较寒冷, 因此鱼类的生长也受到限制。而希腊普雷斯帕湖[18]的麦穗鱼属于入侵种,但是当地麦穗鱼的K值0.25和渐进体长118 mm与本研究的结果并没有太大的差异性, 这也说明了麦穗鱼具有较强的适应能力, 同时这也可能与希腊的气候有关, 希腊属于地中海型气候, 全年气温变化不大, 更加适合鱼类的生长。
由于目前渔获物的主要组成为2龄和3龄个体,缺少小个体、低年龄的个体, 并且渔获物中高龄鱼个体极少, 这种现象的出现, 捕捞的选择性可能是导致渔获物1龄个体样本偏少的主要原因。在采集小型鱼类时, 依靠刺网采集样本, 容易造成渔获物组成的选择性, 应配合其他网具采集。本研究样本采集主要在入湖的3个较大河口处采集, 在南湾湖的中心及其他区域未进行采集, 也会造成样本的选择性。
麦穗鱼属于小型鲤科鱼类, 其分布范围广, 对环境适应能力强, 虽然麦穗鱼不属于经济鱼类, 但在水体食物网中起重要作用, 对水体变化敏感, 可以作为水体生态系统变化的指示生物, 对生态系统的影响较大, 麦穗鱼对维持水生生态系统的稳定具有重要作用, 不仅能够减缓水藻带来的水华的危害,而且对浮游动物的生存动态具有调控作用。
我们计算出麦穗鱼的生长拐点雌性3.607龄, 雄性5.619龄。在拐点年龄之前, 麦穗鱼个体快速生长, 拐点年龄之后生长开始减慢。而我们所采集的麦穗鱼样本年龄都处于拐点年龄之前, 出现这种状况可能是过度捕捞所致。这表明南湾湖麦穗鱼资源利用是不合理的, 过度捕捞使种群结构遭到破坏,资源量日趋减少, 种群出现了明显的小型化趋势,严重影响了麦穗鱼野生资源的可持续性。目前, 应采取限制电、炸、毒捕捞和捕捞渔具规格, 以及在繁殖季节加强保护禁止捕捞等措施, 以利于麦穗鱼资源的恢复和增殖。
[1]Asaeda T, Manatunge J.Foraging of a small planktivore (Pseudorasbora parva: Cyprinidae) and its behavioral flexibility in an artificial stream [J].Hydrobiologia, 2005, 549(1): 155—166
[2]Asaeda T, Manatunge J.Physiological responses of topmouth gudgeon, Pseudorasbora parva, to predator cues and variation of current velocity [J].Aquatic Ecology, 2007, 41(1): 111—118
[3]Nagata T, Ha J Y, Hanazato T.The predation impact of larval Pseudorasbora parva (Cyprinidae) on zooplankton: a mesocosm experiment [J].Journal of Freshwater Ecology, 2005, 20(4): 757—763
[4]Zhang T, Cui Y, Li Z, et al.The ecology of topmouth gudgeon, Pseudorasbora parva, in Bao’an Lake–III.Food habits [J].Acta Hydrobiologica Sinica, 1998, 22(Suppl): 157—167 [张堂林, 崔奕波, 方榕乐, 等.保安湖麦穗鱼种群生物学Ⅲ食性.水生生物学报, 1998, 22(增刊): 157—167]
[5]Britton J R, Davies G D, Brazier M, et al.A case study on the population ecology of a topmouth gudgeon (Pseudorasbora parva) population in the UK and the implications for native fish communities [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2007, 17(7): 749—759
[6]Hu M, Wu Z Q, Liu Y.The fish fauna of mountain streams in the Guanshan National Nature Reserve, Jiangxi, China [J].Environmental Biology of Fishes, 2009, 86(1): 23—27
[7]Onikura N, Nakajima J.Age, growth and habitat use of the topmouth gudgeon, Pseudorasbora parva in irrigation ditches on northwestern Kyushu Island, Japan [J].Journal of Applied Ichthyology, 2013, 29(1): 186—192
[8]Jang M H, Yoon J D, Shin J H, et al.Status of freshwater fish around the Korean demilitarized zone and its implications for conservation [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2008, 18(6): 819—828
[9]Chen I S, Weng C J, Chen Y R, et al.The checklist of inland-water and mangrove fish fauna of Kinmen Island, Fujian Province, Taiwan with comments on ecological conservation of native fishes [J].Journal of Marine Science and Technology, 2013, 21: 316—319
[10]Britton J R, Davies G D, Brazier M, et al.A case study on the population ecology of a topmouth gudgeon (Pseudorasbora parva) population in the UK and the implications for native fish communities [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2007, 17(7): 749—759
[11]Gozlan R E, Andreou D, Asaeda T, et al.Pan-continental invasion of Pseudorasbora parva: towards a better understanding of freshwater fish invasions [J].Fish and Fisheries, 2010, 11(4): 315—340
[12]Chen Y, Li Z, Lin L S.Effects of a small planktivore (Pseudorasbora parva: Cyprinidae) on eutrophication of a shallow eutrophic lake in central China [J].Aquatic Ecosystem Health & Management, 2010, 13(3): 328—334
[13]Fan L Q, Liu H P, Lin J, et al.Non-native fishes: distribution and assemblage structure in the Ihasa River basin, Tibet, China [J].Acta Hydrobiologica Sinica, 2016, 40(5): 958—967 [范丽卿, 刘海平, 林进, 等.拉萨河流域外来鱼类的分布、群落结构及其与环境的关系.水生生物学报, 2016, 40(5): 958—967]
[14]Yan Y, Chen Y.Variations in reproductive strategies between one invasive population and two native populations of Pseudorasbora parva [J].Current Zoology, 2009,55(1): 56—60
[15]Britton J R, Davies G D, Brazier M, et al.A case study on the population ecology of a topmouth gudgeon (Pseudorasbora parva) population in the UK and the implications for native fish communities [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2007, 17(7): 749—759
[16]Zhang T L, Cui Y B, Fang, R L, et al.Population biology of topmouth gudgeon (Pseudorasbora parva) in Bao’an Lake-I Age and growth [J].Acta Hydrobiologica Sinica, 1998, 22(Suppl): 139—145 [张堂林, 崔奕波, 方榕乐,等.保安湖麦穗鱼种群生物学Ⅰ年龄与生长.水生生物学报, 1998, 22(增刊): 139—145]
[17]Onikura N, Nakajima J.Age, growth and habitat use of the topmouth gudgeon, Pseudorasbora parva in irrigation ditches on northwestern Kyushu Island, Japan [J].Journal of Applied Ichthyology, 2013, 29(1): 186—192
[18]Rosecchi E, Crivelli A J, Catsadorakis G.The establishment and impact of Pseudorasbora parva, an exotic fish species introduced into Lake Mikri Prespa (north-western Greece) [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 1993, 3(3): 223—231
[19]Záhorská E, Kováč V, Katina S.Age and growth in a newly-established invasive population of topmouth gudgeon [J].Central European Journal of Biology, 2010, 5(2): 256—261
[20]Davies G D, Britton J R.Influences of population density, temperature and latitude on the growth of invasive topmouth gudgeon Pseudorasbora parva [J].Ecology of Freshwater Fish, 2015, 24(1): 91—98
[21]Lemmens P, Mergeay J, Vanhove T, et al.Suppression of invasive topmouth gudgeon Pseudorasbora parva by native pike Esox lucius in ponds [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2015, 25(1): 41—48
[22]Maceina M J, Boxrucker J, Buckmeier D L, et al.Current status and review of freshwater fish aging procedures used by state and provincial fisheries agencies with recommendations for future directions [J].Fisheries, 2007, 32(7): 329—340
[23]Quist M C, Jackson Z J, Bower M R, et al.Precision of hard structures used to estimate age of riverine catostomids and cypri-nids in the upper Colorado River basin [J].North American Journal of Fisheries Management, 2007, 27(2): 643—649
[24]Yan Y Z, Xu Y S, Chu L, et al.Age, growth and reproduction of Sarcocheilichthys nigripinnis from the Qingyi Stream in the Huangshan Mountains [J].Zoological Research, 2012, 33(E1-2): 25—31
[25]Pauly D.A preliminary compilation of fish length growth parameters [J].Berichte des Instituts für Meereskunde an der Christian-Albrechts Universität, Kiel, 1978, 55: 1—200
[26]Tarkan A S, Copp G H, Zięba G, et al.Growth and reproduction of threatened native crucian carp Carassius carassius in small ponds of Epping Forest, south-east England [J].Aquatic Conservation: Marine and Freshwater Ecosystems, 2009, 19(7): 797—805
[27]Kumar R S, Sarkar U K, Gusain O, et al.Age, growth, population structure and reproductive potential of a vulnerable freshwater mullet, Rhinomugil corsula (Hamilton,1822) from a tropical river betwa in central India [J].Proceedings of the National Academy of Sciences, India Section B: Biological Sciences, 2014, 84(2): 275—286
[28]Zhou C, Zhu Q, Liu H Z.Analysis of the growth function of the largemouth bronze gudgeon Coreius guichenoti in the Upper Reaches of the Yangtze River [J].Sichuan Journal of Zoology, 2010, 29(4): 510—516 [周灿, 祝茜,刘焕章.长江上游圆口铜鱼生长方程的分析.四川动物, 2010, 29(4): 510—516]
[29]Quince C, Abrams P A, Shuter B J, et al.Biphasic growth in fish I: theoretical foundations [J].Journal of Theoretical Biology, 2008, 254(2): 197—206
[30]Cope J M, Punt A E.Admitting ageing error when fitting growth curves: an example using the von Bertalanffy growth function with random effects [J].Canadian Journal of Fisheries and Aquatic Sciences, 2007, 64(2): 205—218
[31]Ivancheva E Y, Ivanchev VP, Sarychev V S.Distribution of stone moroko (Pseudorasbora parva) in the Upper Don basin [J].Russian Journal of Biological Invasions, 2014, 5(3): 151—155
STUDY ON THE AGE AND GROWTH OF PSEUDORASBORA PARVA FROM NANWAN LAKE UPSTREAM THE HUAIHE RIVER
LI Hong-Jing, WANG Ya-Ping, LENG Qiu-Li, LI Xiao-Jie, LI Xiao-Feng, YU Tong-Lei and HUANG Bin
(College of Life Sciences, Xinyang Normal University, Xinyang 464000, China)
Pseudorasbora parva is a native small cyprinid fish widely distributes in Asia with a considerable population.To investigate the growth, age composition and growth characteristics of Pseudorasbora parva, 532 individuals were collected from three different water inlets of Nanwan Lake in March, May, July and December.The results showed that the body length were ranged from 35.82 to 88.28 mm with the mean of (61.61±11.8) mm, and that the body weight were ranged from 3.07 to 59.17 g with the mean of (19.23±10.73) g.The age were ranged from 1 to 3.3 years and the dominant population with the mean of 57.38% were individuals with 3 years.The relationship between body length and body weight were W=9.602E–5L2.928(R2=0.883) for female and W=4.487E–5L3.116(R2=0.889) for male, which showed a significant difference between male and female (F=5.241, P<0.05).A linear relationship was found between the scale radius and body length, which was significantly affected by gender (F=78.405, P<0.05).Growth parameters were L∞=107.005, K=0.246 and t0= –0.76 for female and L∞=145.254, K=0.181 and t0= –0.66 for male, respectively.The growth inflexion of female (3.607 years) were 70.46 mm for body length and 24.72 g for body weight, and the growth inflexion of male (5.619 years) were 98.64 mm for body length and 73.53 g for body weight.These results suggest that the female was isauxesis while the male was allometric, and that the male had a faster growth rate than female.
Lake Nanwan; Pseudorasbora parva; Age; Growth
Q178.1
A
1000-3207(2017)04-0835-08
10.7541/2017.104
2016-04-05;
2016-12-14
国家自然科学基金(1204306)资助 [Supported by the National Natural Science Foundation of China (1204306)]
李红敬(1966—), 男, 河南新野人; 博士; 研究方向为动物生态学。E-mail: lhj@xynu.edu.cn
