高产絮凝物蝉花菌株筛选和絮凝条件初探
2017-08-09孟凡丽孙嘉龙
李 娟,邹 晓,孟凡丽,孙嘉龙
(1.贵州大学 生命科学学院/真菌资源研究所,贵州 贵阳 550025;2.贵州省环境科学研究设计院,贵州 贵阳 550081;3.贵州理工学院资源与环境工程学院,贵州 贵阳 550081)
高产絮凝物蝉花菌株筛选和絮凝条件初探
李 娟1,邹 晓1,孟凡丽2,孙嘉龙3*
(1.贵州大学 生命科学学院/真菌资源研究所,贵州 贵阳 550025;2.贵州省环境科学研究设计院,贵州 贵阳 550081;3.贵州理工学院资源与环境工程学院,贵州 贵阳 550081)
为探究虫生真菌蝉花不同菌株之间产絮凝剂的差异及絮凝物的絮凝特性,以高岭土法对36株蝉花菌株进行了高产絮凝物的筛选及高产菌株的生长特性和絮凝性质的研究。实验筛选出稳定、高产絮凝物的蝉花菌株GZUIFR-6722,絮凝物活性物质是其胞外聚合物。该菌株在生长过程中生物量的积累和产絮凝物能力呈正相关。当菌株6 722絮凝物投加量为1.2 mL每50 mL时,絮凝效果较好,絮凝率可达91.47%。1%的CaCl2能显著促进絮凝效果,絮凝率可达94.89%。颠倒次数、静置时间、温度以及pH对絮凝效果影响不显著。
微生物絮凝物;蝉花;絮凝特性
蝉花(蝉棒束孢Isariacicadae)是一种虫草类真菌,对蚜虫、小菜蛾、白粉虱等害虫的防治效果较好[1,2],同时也是一味传统名贵中药,在糖尿病、抗肾纤维化进程、延缓肾衰竭等的临床及实验研究中,取得了较好效果[3,4]。迄今为止,对蝉棒束孢透明质酸、多糖及糖蛋白等活性物质[5-7]和抗衰老、改善免疫功能等[8,9]在医疗和保健上已有大量的研究报道,因此,蝉花是一种极具应用前景的高安全性的真菌资源。除此之外,也有研究发现蝉花能产生絮凝物,郭旭辉等[10]率先发现蝉花发酵液具有絮凝活性,采用pH为7.0的马铃薯液体培养基150 r/min、30℃摇床培养84 h后絮凝率可达85.1%。孙嘉龙等[11]从土壤中分离到一株具高效絮凝活性的菌株3 716,并对其培养条件与絮凝活性进行了研究,由此可见,蝉花的絮凝能力也可用于微生物絮凝物的开发。
微生物絮凝物被称为继无机絮凝物和合成有机高分子絮凝物之后的第三代絮凝物[12],是由微生物产生、经分离和提纯获得的具有絮凝活性的次生代谢产物[13],其有效成分一般为多糖、蛋白质、脂类和核酸等生物大分子[14,15]。相对于传统的无机及合成高分子絮凝物而言,微生物絮凝物具有易降解、安全无毒以及无二次污染等优点。与微生物絮凝物相比,无机和合成有机高分子絮凝物对环境不可避免的破坏限制了它们的应用[16]。因此,从环境安全角度考虑,微生物絮凝物是目前絮凝物产品中的研究热点,是一类极具发展前途的水处理剂[17],可广泛应用于饮用水、食品废水、医药废水及重金属废水等领域[18-20]。微生物絮凝物生产菌来源广泛,目前已报道了50余种絮凝物生产菌[21],分别属于细菌、真菌、放线菌及藻类,其中以细菌和真菌占绝大部分。筛选更安全有效的絮凝物生产菌、降低菌株产絮凝物成本、探究絮凝机理是目前微生物絮凝物领域的研究热点。为丰富真菌絮凝物菌株资源库,本文着眼于安全性更高、更具特色的虫草类真菌絮凝物进行研究。
虫生真菌蝉花具有优越产絮凝物性质,但考虑到研究所用的菌株不同,菌株之间的絮凝能力也会存在差异。本文对来自贵州大学真菌资源研究所的36株蝉花菌株产絮凝物能力和絮凝条件进行了探索。
1 材料与方法
1.1 材料
1.1.1 菌株 用于实验的36株蝉花菌株(表1),现保存于贵州大学真菌资源研究所(GZUIFR)。菌株来自全国各地及韩国济州岛(JZD)。
1.1.2 培养基 a.PDA(g/L):土豆200 g,葡萄糖20 g,琼脂15~18 g;b.液体培养基(g/L):PDA中去除琼脂。
1.2 蝉花高产絮凝物菌株的筛选
1.2.1 菌株培养 将蝉花菌株活化,于25℃恒温培养箱中倒置培养5~7 d后用无菌水洗下平板上的孢子,调整浓度为1×106个/mL,接种到液体培养基。每株3个平行,放置于恒温振荡培养箱中,140 r/min、25℃条件下培养,7 d后取发酵液用于絮凝试验。
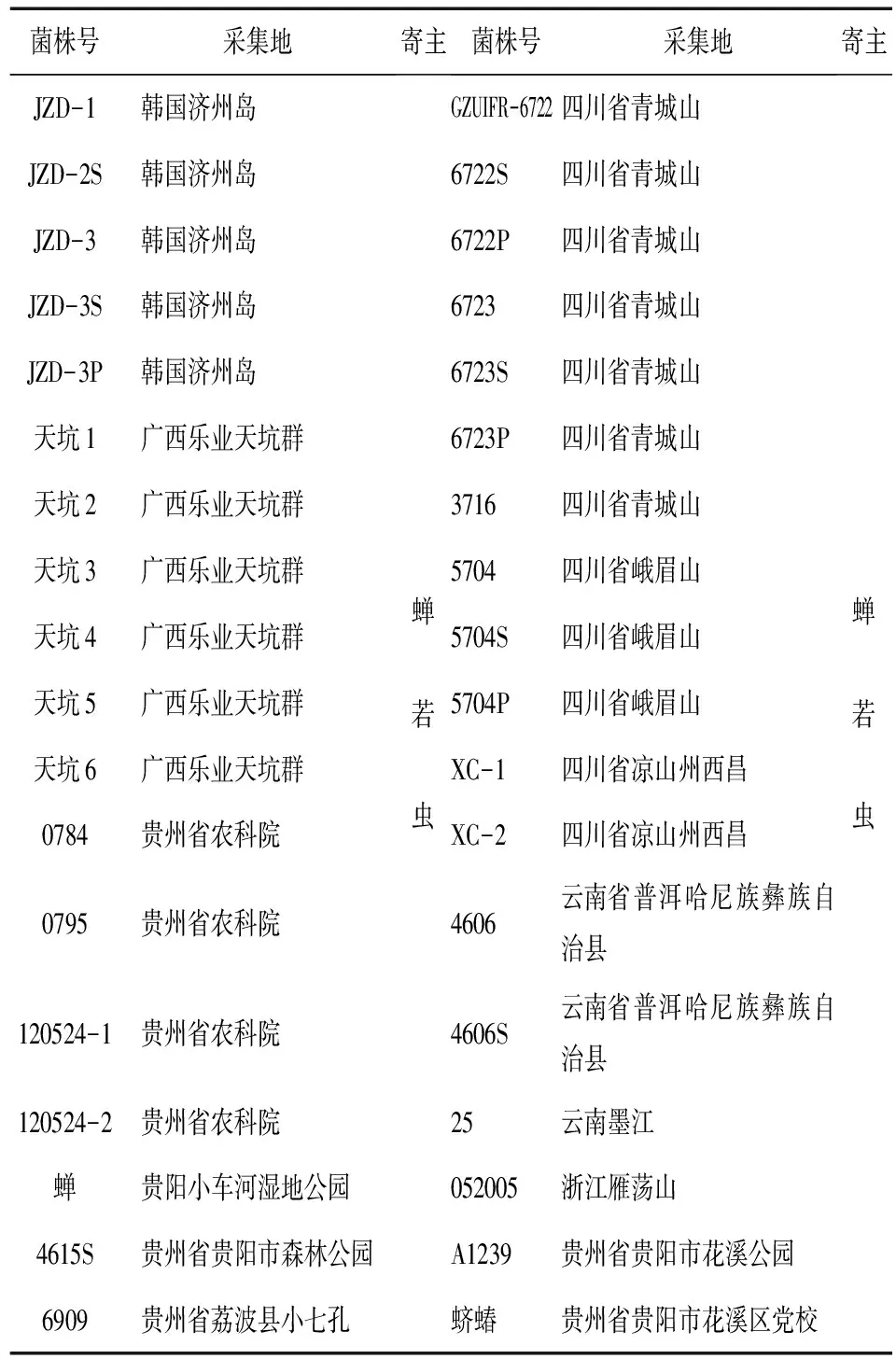
表1 贵州大学真菌资源研究所36株蝉花菌株
1.2.2 絮凝率的测定 在50 mL的比色管中加入49 mL 4 g/L的高岭土悬浊液、1 mL蝉花发酵液,将比色管颠倒10次,每次以气泡上升完毕为准,静置20 min,取液面1 cm以下液体,在550 nm处测定吸光值(B),以添加蒸馏水的吸光值(A)为对照,每个处理3个平行样;确定发酵液的絮凝活性,用絮凝率E(%)来表示[22]。絮凝率计算公式:絮凝率E(%)=(A-B)/A×100%。
1.3 菌株GZUIFR-6722活性物质的分布测定
取培养7 d后的成熟发酵液,分别测定发酵原液、6 000 r/min离心10 min后的上清液、悬浮在等量蒸馏水中的菌丝段、蒸馏水洗涤3~5次后悬浮在等量蒸馏水中的菌丝段以及洗涤液共5种组分的絮凝率[23]。
1.4 菌株GZUIFR-6722生长特性
1.4.1 液体发酵的生长曲线测定 对液体发酵培养的菌株6 722,每24 h取20 mL菌液离心获得菌体,蒸馏水洗涤后,将菌体80℃烘干至恒重,称其质量,设置三个平行。同时进行称重法补水[24]。
1.4.2 培养时间对絮凝物絮凝活性的影响 将菌株6 722接种到PDA液体培养基中进行发酵培养,每1 d取样测定上清液对高岭土的絮凝活性。
1.4.3 培养培养过程中发酵液pH的变化 测定发酵液的pH的变化,每1 d取样测定一次。
1.5 絮凝物的分离纯化及絮凝特性、温度和pH对蝉花絮凝物絮凝效果的影响1.5.1 絮凝活性物质的分离纯化 取培养7 d后的成熟发酵液,6 000 r/min离心10 min,收集上清液。使用旋转蒸发仪,将温度控制在50℃左右,浓缩发酵上清液至原体积的1/3左右。加入2倍体积预冷至4℃的无水乙醇,轻微振荡混匀,4℃冰箱中静置18 h。6 000 r/min离心15 min,得到沉淀物用95%乙醇清洗两次,冷冻干燥得到蝉花絮凝物[25],该产品用于絮凝特性和絮凝物稳定性的研究。1.5.2 蝉花絮凝物的投加量对絮凝物絮凝活性的影响 将蝉花絮凝物配制成1 g/L的絮凝物。设置絮凝物的投加量分别为0、0.4、0.8、1.2、1.6、2.0、2.4、2.8、3.2、3.6、4.0、4.4 mL,以蒸馏水作为对照,测定絮凝率,探究絮凝物投加量对絮凝效果的影响。
1.5.3 不同阳离子及阳离子的浓度对蝉花絮凝物絮凝活性的影响 不同阳离子类型:配制质量浓度为1%的KCl、NaCl、MnCl2、CaCl2、MgSO4五种阳离子,以蒸馏水作为对照,测定絮凝率,探究不同阳离子类型对絮凝物絮凝活性的影响。不同阳离子浓度:配制质量浓度为0.5、1.0、1.5、2.0、2.5、3.0、3.5%的CaCl2,以蒸馏水作为对照,测定絮凝率,探究不同阳离子浓度对絮凝物絮凝活性的影响。
1.5.4 颠倒次数及静置时间 颠倒次数:设置颠倒次数梯度:5、10、15、20、25、30、35、40次,以蒸馏水作为对照,测定絮凝率,探究不同颠倒次数对絮凝物絮凝活性的影响。静置时间:设置静置时间梯度:5、10、15、20、25、30、35、40 min,以蒸馏水作为对照,测定絮凝率,探究不同静置时间对絮凝物絮凝活性的影响。
1.5.5 温度和pH对蝉花絮凝物絮凝效果的影响 悬浮液温度:通过制冷和加热调节高岭土悬浊液的温度为10、20、30、40、50、60、70、80、90℃,以蒸馏水作为对照,测定絮凝率,探究高岭土悬浊液温度对絮凝效果的影响。悬浮液pH:测定高岭土悬浊液的初始pH,用1 mol/L的HCl和1 mol/L NaOH调节高岭土悬浊液的pH为2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0,以蒸馏水作为对照,测定絮凝率,探究高岭土悬浊液pH对絮凝效果的影响。
2 结果与分析
2.1 蝉花高产絮凝物菌株的筛选
对贵州大学真菌资源研究所保存的36株蝉花进行了絮凝能力的测定(表2),36株蝉花都具有一定的絮凝能力,但不同菌株之间存在着絮凝能力与产絮凝物稳定性之间的差异,以絮凝率为标准,比较多次重复结果的标准误差,以标准误差来衡量菌株产絮凝物的稳定性。筛选出其中产絮凝物较为稳定且絮凝能力较强的菌株为GZUIFR-6722,后续絮凝性质研究以GZUIFR-6722为实验对象。
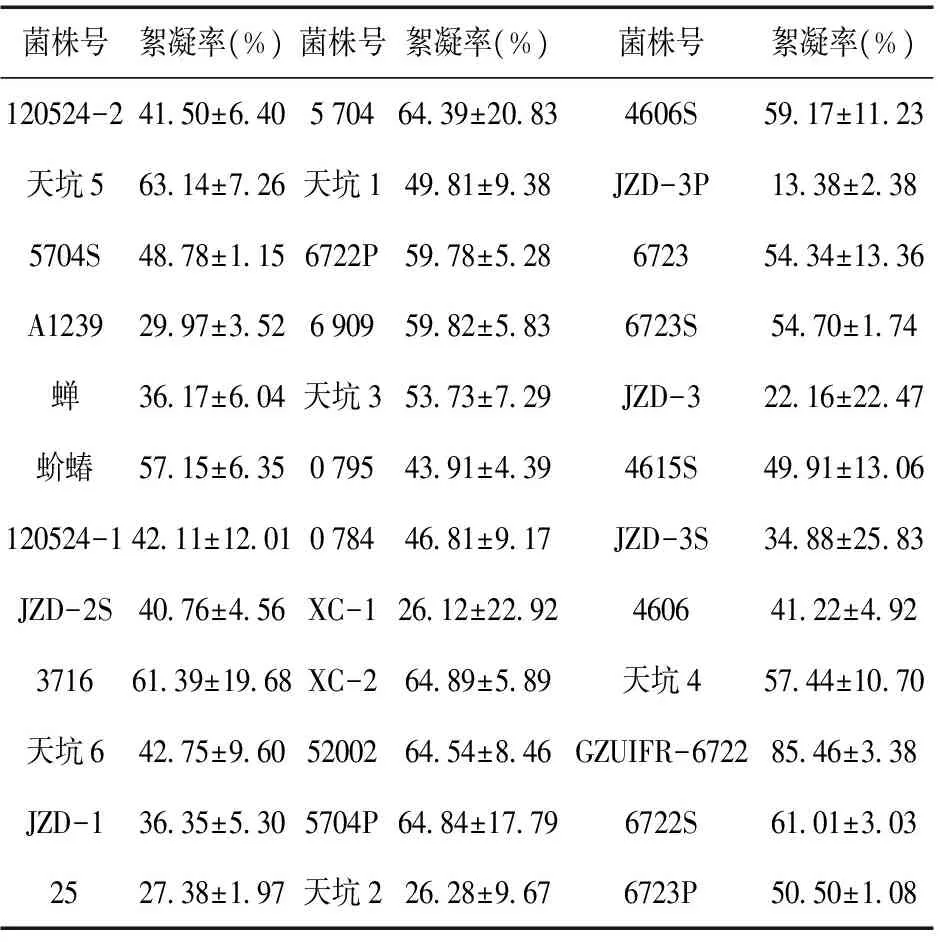
表2 贵州大学真菌资源研究所36株蝉花菌株絮凝效率
2.2 絮凝活性物的检测
絮凝活性物质分布测定结果如图1所示,GZUIFR-6722的5种组分絮凝率依次为:上清液(87.78±0.60)%>发酵原液(85.02±2.36)%>洗涤液(32.11±0.50)%>离心后菌体悬浮液(24.37±2.08)%>清洗后菌体悬浮液(16.45±1.45)%。离心后,上清液絮凝率较发酵原液稍高;离心后的菌丝体具有一定的絮凝能力,但经过清洗后絮凝率降低,而洗涤液具有了絮凝能力且絮凝率高于菌体悬浮液,说明附着在菌体上的絮凝活性物质被洗脱到蒸馏水中,上清液的高絮凝率表明GZUIFR-6722产生的絮凝物主要存在于发酵上清液中,是由菌体细胞分泌到胞外的。
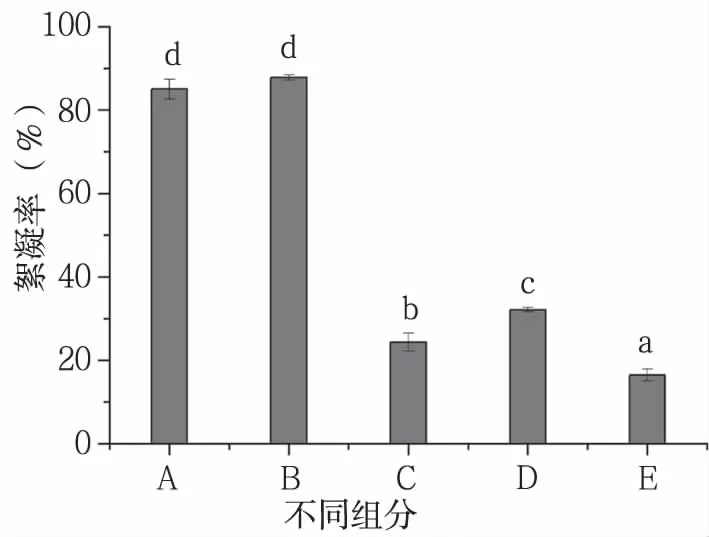
图1 GZUIFR-6722产生絮凝物絮凝效率
A:发酵原液;B:上清液;C:离心后菌体悬浮液;D:洗涤液;E:清洗后菌体悬浮液
注:图中不同字母代表差异显著(P<0.05),下同。2.3 菌株GZUIFR-6722的生长特性
2.3.1 菌株GZUIFR-6722培养过程中的生物量与产絮凝物相关性 测定了GZUIFR-6722在培养的8 d过程中,生长曲线和产絮凝物动力学曲线(图2)。GZUIFR-6722培养到第3 d到第4 d中生物量只有少量增加,第5 d,菌株进入稳定期后不再生长,开始累积代谢产物,第6 d后菌株进入衰老期,生物量开始下降;在培养过程中,随着菌株的生长,菌株的絮凝率随生物量的累积而增加,最大絮凝率(79.62±0.34)%出现在菌株生长稳定后期至衰老期的第7 d,比生物量最大值的时间延迟,然后急剧下降,可能是菌株由于营养物质的消耗而开始分解利用胞外代谢产物导致。总的来说,GZUIFR-6722的生物积累和产絮凝物能力成正相关。
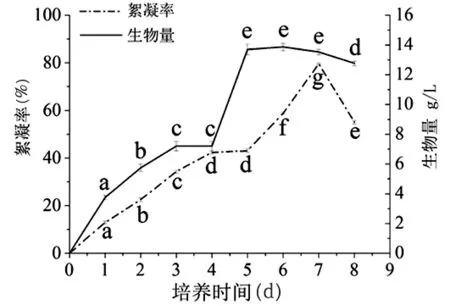
图2 GZUIFR-6722的生长与产絮凝曲线
2.3.2 菌株GZUIFR-6722培养过程中pH的变化 GZUIFR-6722在生长过程中,pH呈现出一个先下降后趋于稳定(图3),可能是菌株在生长期内产生了酸性物质而使pH降低[26],培养前4天pH变化呈显著性降低,在第4天发酵液的pH最低,为(4.16±0.32)。
2.4 蝉花絮凝物的絮凝特性
2.4.1 蝉花絮凝物投加量对絮凝效果的影响 蝉花絮凝物的不同投加量对其絮凝效率的影响不同(图4)。当絮凝物的投加量为0.4~1.2 mL/50 mL时,絮凝率随投加量的增加而增加,并在1.2 mL时达到最高值(91.47±2.43)%;但在1.2~4.4 mL/50 mL时,絮凝率随投加量的增加反而降低,可能是当絮凝物的添加量增加后,大颗粒被吸附的聚合物包围,悬浮物之间达到了一种新的相互排斥的电荷平衡,导致絮凝率下降[27]。

图3 GZUIFR-6722生长过程中的pH变化
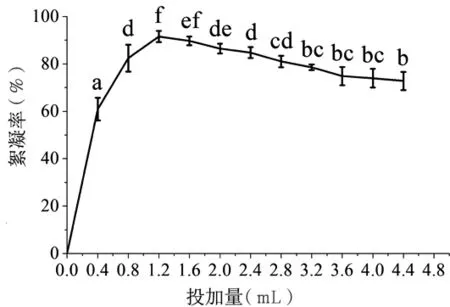
图4 絮凝物投加量对其絮凝效果的影响
2.4.2 不同阳离子及阳离子的浓度对絮凝效果的影响
2.4.2.1 不同阳离子类型 Junichi T等[28]研究表明,絮凝体系中,不同的阳离子类型对微生物絮凝物的絮凝效果影响不同。图5显示了5种不同阳离子对蝉花絮凝物的影响。结果显示:与对照相比,5种阳离子对蝉花絮凝物都有一定促进作用,其中CaCl2的效果最为显著,絮凝率高于对照18.39%,后续以CaCl2作为蝉花絮凝物的阳离子助剂进行研究。
2.4.2.2 不同CaCl2浓度 不同浓度的CaCl2对蝉花絮凝物的影响效果不同(图6),添加CaCl2后于对照相比差异显著,其中CaCl2浓度为1%时促进效果最为明显,絮凝率达(94.89±2.58)%。后续的研究阳离子助剂CaCl2的浓度为1%。
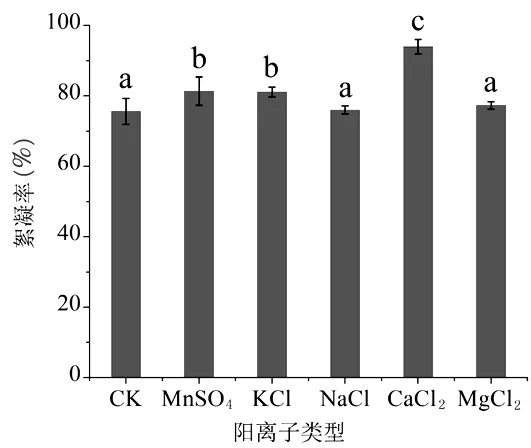
图5 阳离子类型对絮凝物效果的影响
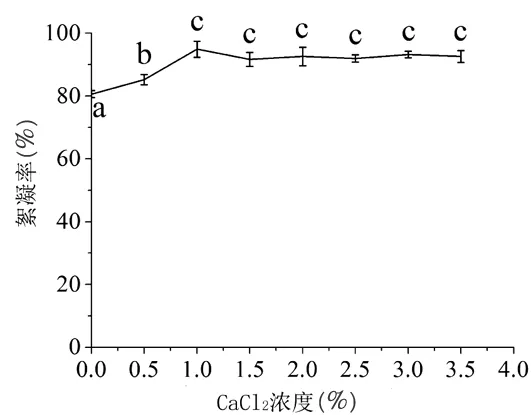
图6 CaCl2浓度对絮凝物效果的影响
2.4.3 颠倒次数及静置时间对絮凝效果的影响
2.4.3.1 静置时间 微生物絮凝物与高岭土作用时,会形成大量的絮体,这些絮体的沉降需要一定的时间。在一定的静置时间内,蝉花絮凝物与高岭土形成的絮凝体随着静置时间的延长而相继沉降,10 min后絮凝率最大且不同处理之间没有显著差异。因此后续的实验静置时间为10 min。
2.4.3.2 颠倒次数 颠倒次数就是对整个絮凝体系混匀程度的研究。颠倒次数从5~40次,蝉花絮凝物的絮凝率均大于92%,不同颠倒次数之间没有显著差异,因此之后的实验颠倒次数为5次。
2.4.4 温度和pH对蝉花絮凝物絮凝效果的影响
2.4.4.1 悬浮液温度 设置不同的高岭土悬浮液温度,探究蝉花絮凝物在应用过程中对悬浮液温度的稳定性(图7)。蝉花絮凝物对悬浮液温度较为稳定,悬浮液温度在10~90℃之间,蝉花絮凝物的絮凝率维持在93%以上,方差分析显示各个温度之间差异不显著,蝉花絮凝物对温度较为稳定。在实际水处理中,可以不考虑温度对蝉花絮凝物效果的影响。
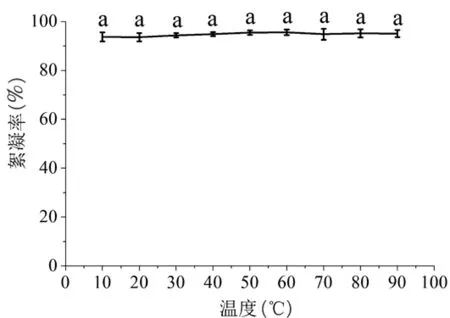
图7 温度对絮凝物活性的影响
2.4.4.2 悬浮液pH 实际水处理中,由于处理不同的原料以及处理工序的不同,产生的水会有不同的酸碱度,会影响絮凝物的使用效果。设置不同的悬浮液的pH,探究蝉花絮凝物在使用过程中对悬浮液pH的稳定性(图8)。结果显示:pH在2~10范围之间,絮凝效果均维持在90%以上。相较于碱性环境下,弱酸性环境下的絮凝效果较优,调节pH在2~9后的高岭土悬浮液与对照相比,絮凝率差异不显,蝉花絮凝物对悬浮液pH较为稳定。在实际的水处理中,当pH在2~10之间时,这种较小的差异可以忽略不计,可以不考虑pH对蝉花絮凝物效果的影响。

图8 pH对絮凝物效果的影响
3 结论与讨论
本研究筛选出一株能稳定高效产絮凝物的蝉花菌株GZUIFR-6722,分析了菌株产生絮凝物活性物质的分布,发现活性物质是其胞外产物,与多数真菌生产絮凝物一致[13,25]。方差分析显示温度和pH对蝉花絮凝物效果的影响差异不显著,猜测该絮凝物可能是受温度和pH影响较小的胞外多糖,对胞外多糖的组成及理化性质的研究将有助于了解絮凝物的絮凝机理,胞外多糖的提取方法不同会导致提取量也不同[13],后续研究有待建立起蝉花胞外多糖的温和、高效、操作简单的提取方法。在生长过程中,GZUIFR-6722生物量的积累和产絮凝物能力成大致的正相关,该性质有利于在蝉花絮凝物工业生产过程中调控絮凝物的生产技术,在低细胞死亡率的前提下,最大程度地提取出蝉花絮凝物。CaCl2作为阳离子助剂能显著促进蝉花絮凝物的效果,与前人研究一致[11]。颠倒次数和静置时间对絮凝效果的影响差异不显著,颠倒5次,静置10 min的絮凝率达(94.15±2.08)%,在水处理过程中可以根据实际情况进行调节,进一步达到节约人力物力的效果,且蝉花絮凝物对温度和pH不敏感,所以在实际的水处理过程中不用考虑温度和pH对该絮凝物的影响,减少对温度和pH调节,可极大地节约资源成本和使处理程序简约化。
目前,微生物絮凝物由于生产成本较高,絮凝物产量较低等因素在根本上限制了其在实际工业中的应用[29],后续研究应集中在GZUIFR-6722高产絮凝物的快速、低成本培养方法,使其有望进一步应用水处理领域。
[1] 刘爱英,李 忠,邹 晓,等.中国蝉花资源研究应用[M].贵阳:贵州科技出版社,2012.
[2] 邹 晓,刘爱英,梁宗琦.虫生真菌蝉拟青霉诱变株对温室白粉虱的致死因素分析[J].植物保护,2007(2):73-76.
[3] 刘伟敬,唐阳敏,谢淑华,等.蝉花菌丝对糖尿病肾病大鼠的肾脏保护作用机制研究[J].中国中西医结合肾病杂志,2014,15(8):665-668.
[4] 任颍芳,朱 戎.中药蝉花药物考证及其防治肾病的研究进展[J].河南中药,2013,33(5):781-783.
[5] 谭爱娟,武立琨,余晓培.蝉拟青霉产透明质酸最佳液体培养基的筛选[J].食品科学,2007,28(8):294-296.
[6] 刘 芸,谭爱娟,吕世明.酶法提取蝉拟青霉多糖的研究[J].天津农业科学,2010,16(2):36-38.
[7] 谭爱娟,梁宗琦,刘爱英.蝉拟青霉菌丝体糖蛋白的分离提取及组分测定[J].食品科学,2007,28(1):159-160.
[8] 杨介钻,金丽琴,吕建新,等.蝉拟青霉多糖抗衰老作用的实验研究[J].中国老年学杂志,2004,4(24):343-344.
[9] 杨介钻,卓 佳,陈柏坤.蝉拟青霉多糖对老年大鼠免疫功能的调节作用[J].中国中药杂志,2008,33(3):292-295.
[10] 郭旭辉,周文霞,梁宗琦,等.一株蝉拟青霉菌产絮凝活性的研究[J].武汉大学学报(理学版),2004,50(S2):031-033.
[11] 孙嘉龙,李 忠.微生物絮凝剂产生菌的选育及条件优化[J].贵州农业科学,2010,38(11):169-171.
[12] 谢玉清,茆 军,张志东,等.一株产絮凝剂的类芽孢杆菌新种及絮凝特性研究[J].环境科学与技术,2014,37(4):7-10.
[13] 时翠翠,黄 强,方 迪,等.生物絮凝丝状真菌Talaromycessp.CC-1胞外多聚物的提取及组分分析[J].环境化学,2014,33(4):624-629.
[14] 马 放,段妹悦,孔祥震,等.微生物絮凝剂的研究现状及其发展趋势[J].中国给水排水,2012,28(2):14-17.
[15] 王 琴,杨劲峰,赵继红.微生物絮凝剂在废水处理中的应用研究[J].广州化工,2013,1(12):46-48.
[16] 刘洪艳,王红玉.潮间带污泥产絮凝细菌W17分离与絮凝性质分析[J].环境科学与技术,2015,38(8):127-130.
[17] 常玉广,马 放,夏四清,等.絮凝微生物的功能定位及其絮凝成分分析[J].同济大学学报(自然科学版),2009,37(1):84-86,96.
[18] Zhang Zhi-qiang,Lin Bo,Xia Si-qing,etal.Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source[J].JournalofEnvironmentalSciences,2007,19(6):667-673.
[19] 张云波,李 政,刘其友,等.微生物絮凝剂产生菌KJ-10的絮凝性能及絮凝成分分析[J].环境工程学报,2011,5(4):951-955.
[20] 姚敏杰,连 宾.微生物絮凝剂对高浓度重金属离子废水絮凝作用研究[J].环境科学与技术,2009,32(11):1-4.
[21] 曾 苏,陈晓平,李南华,等.微生物絮凝剂生产菌T1的鉴定及其对生活污水絮凝特性[J].环境化工,2015,34(3):578-583.
[22] Kurane R,Hatamochi K,Kakuno T,etal.Production of a bioflocculant by Rhodococcus erythropolis S-1 grown on alcohols[J].BiosciBiotechBioch,1994,58(2):428-429.
[23] 刘立凡,阮彩群.糖蜜废液培养微生物絮凝剂及絮凝特性研究[J].环境科学与技术,2009,32(8):152-156.
[24] 胡子全,赵海泉.一株有机解磷菌的筛选及其最佳生长条件的研究[J].中国给水排水,2007,23(17):66-71.
[25] 黄 强.黑曲霉A80产生物絮凝剂的培养条件及其絮凝特性研究[D].青岛:中国海洋大学,2012.
[26] 杨生玉,王 刚,沈永红.微生物生理学[M].北京:化学工业出版社,2007.
[27] 屹 立,赵 吉,刘 君,等.高效微生物絮凝剂生产菌的筛选及其絮凝特性分析[J].中国给排水,2007,23(21):62-66.
[28] Junichi T,Hiroaki T.Manufacture of flocculant PF-102 with the fungus Paecilomyces[J].Gar.Offen.DE,1986,86(134):704-799.
[29] 王馨蔚,莫创荣,梁 敏,等.木薯淀粉废水培养复合微生物絮凝剂产生菌的营养条件优化[J].食品工业科技,2016(04):211-216.
Screening of High Bioflocculability Strain ofIsariacicadaeMigand the Flocculating Conditions
LIJuan1,ZOUXiao1,MENGFan-li2,SUNJia-long3*
(1.SchoolofLifeSciences,GuizhouUniversity,Guiyang,Guizhou550025,China;2.ZuizhouInstituteofEnvironmentalScienceResearchandDesignGuiyang,Guizhou550081,China;3.SchoolofResourcesandEnvironmentalEngineering,GuizhouInstituteofTechnology,Guiyang,Guizhou550081,Chian)
The aim of this paper is to study the bioflocculability and characteristics of different strains ofIsariacicadaeby screening all 36 strains with kaolin.The results showed that GZUIFR-6722 strain can perform steadily with high bioflocculability.The active substrate was identified as some kind of extracellular metabolite.During the growth,the production of active substrate was related positively with the biomass.The flocculating rate was 91.47% after addition of 1.2 mL/50 mL bioflocculants.Moreover,the addition of 1% CaCl2could promote the performance of the bioflocculants,and increased the flocculating rate to 94.89%.Furthermore,the results also indicated that inversion,standing time,temperature and pH had no obvious effect on flocculation.The findings suggest for application of the GZUIFR-6722 strain in the treatment of waste water.
microbial flocculants;Isariacicadae;flocculation characteristics
2017-02-20;
2017-03-24
国家自然科学基金(41563010;31360031);贵州理工学院高层次人才科研项目(XJGC20140605);贵州省社发攻关项目(黔科合SY字[2013]3143号);贵州省教育厅创新群体重大研究项目(黔教合KY字[2016]045)。
X172
A
1008-0457(2017)03-0027-06 国际
10.15958/j.cnki.sdnyswxb.2017.03.005
*通讯作者:孙嘉龙(1978-),男,博士,教授,主要研究方向:真菌资源与生态应用;E-mail:danielsjl@163.com。
