不同家系马尾松对持续干旱的响应及抗旱性*
2017-07-18杜明凤丁贵杰赵熙州
杜明凤 丁贵杰 赵熙州
(1.贵州大学林学院 贵州省森林资源与环境研究中心 贵阳 550025; 2.贵州师范大学喀斯特研究院 贵阳 550001)
不同家系马尾松对持续干旱的响应及抗旱性*
杜明凤1,2丁贵杰1赵熙州1
(1.贵州大学林学院 贵州省森林资源与环境研究中心 贵阳 550025; 2.贵州师范大学喀斯特研究院 贵阳 550001)
【目的】 了解持续干旱对不同家系马尾松幼苗生长和生理特征的影响,初步揭示马尾松的抗旱机制及抗旱遗传效应,比较不同家系的抗旱能力,为马尾松抗旱优良家系选育提供理论依据。【方法】 以3个来自广西和贵州的2年生马尾松家系(GX01、GX02和GZ01)幼苗为试验材料,采用随机区组设计,温室土培盆栽模拟干旱环境,研究持续干旱胁迫对马尾松幼苗生长、渗透调节和抗氧化系统的影响。【结果】 随干旱胁迫程度的加重,3个家系马尾松的SOD活性均呈先升后降趋势,各家系在胁迫0~10天存在显著差异(P<0.05); POD活性亦呈先升后降趋势,但家系间差异不显著(P>0.05); MDA含量变化中GX01呈先升后降趋势,GX02和GZ01呈先降后升趋势,各家系在胁迫5~20天以及第30天存在显著差异(P<0.05); 脯氨酸含量变化中GZ01呈持续上升趋势,GX02和GX01呈先升后降又再升高的趋势,各家系在胁迫20~30天存在显著差异(P<0.05)。连续干旱限制马尾松苗期的生长,3个家系马尾松的苗高和地径的生长以及干物质积累均受到抑制,家系间的苗高生长、地径生长、干物质积累等均存在显著差异(P<0.05); 连续干旱导致GX01和GX02的根冠比显著高于对照,GZ01亦高于对照,但差异不显著。隶属函数分析得出 3 个马尾松家系幼苗抗旱性GX01>GX02>GZ01; 灰色关联分析发现,根冠比、苗高与抗旱性关联度最大,其次为MDA和脯氨酸含量,可作为评价抗旱性的主要指标。【结论】 马尾松幼苗在轻度干旱胁迫下通过增加保护酶活性抵御干旱; 在中、重度干旱胁迫下通过增加渗透调节物质抵御干旱逆境伤害; 极重度干旱胁迫下,保护酶活性显著下降,MDA含量显著上升,膜质过氧化伤害严重。干旱胁迫下,马尾松幼苗通过增加地下部分生长、减少地上部分生长以适应干旱逆境。马尾松抗旱性受遗传因素控制,不同家系间抗旱性的差异可能与其不同地理环境长期驯化所形成的遗传差异有关。
马尾松; 干旱胁迫; 生长特性; 生理响应; 抗旱性; 遗传特性
随着全球气候变暖加剧,干旱问题日趋严重,植物对干旱胁迫的适应性及抗旱机制,成为干旱、半干旱区域植物遗传、生态等众多领域的研究热点。干旱胁迫下,植物在形态、结构、生长、光合、生理及代谢等方面发生变化以适应干旱(Fangetal., 2015; Amritaetal., 2004; Shulaevetal., 2008),主要表现为系统抗旱和细胞抗旱(康宗利等, 2006)。系统抗旱指植物体通过减慢生长、改变形态和结构、降低生物量等方式来降低植物失水的伤害,如油蒿(Artemisiaordosica)(王林龙等, 2015)、欧洲云杉(Piceaabies)(Majdietal., 2005)。细胞抗旱则通过调节细胞的生理变化来抵御干旱,如: 水分缺损时,调节细胞渗透物质含量以维持细胞膨压,增强细胞亲水性; 或启动内源保护系统,增强酶类和非酶类抗氧化剂活性,维护体内活性氧代谢的平衡,以抵御过氧化膜质损伤等,如柔毛栎(Quercuspubescens)(Antoninoetal., 2011)、麻疯树(Jatrophacurcas)(Silvaetal., 2010)。
马尾松(Pinusmassoniana)是我国南方主要的针叶树种,具有速生丰产、适应性强、综合利用程度高等优点,在我国林产化工业、森林资源发展及森林生态服务功能中具有重要地位(丁贵杰等, 2006)。贵州、广西是我国重要的马尾松商品用材林产区,这些区域虽年降水量充足,但雨水分布不均且伴有多发季节性干旱,严重制约马尾松用材林的发展。近年,关于马尾松适应干旱的研究主要集中在种源水平上苗木生长和生物量(胡晓健等, 2010),或菌根化苗木的生理响应及抗旱性(王艺等, 2013)等方面,上述研究对马尾松抗旱机制做了有益探索,但指标选择上存在一定局限,评价马尾松抗旱性时不够客观全面; 同时,各种生理响应的内在联系研究较少,哪种生理反应最先响应,引发哪些后续生理变化等研究尚待深入; 此外,对马尾松抗旱机制与其遗传特性的揭示尚属空白,探究马尾松对干旱响应的遗传差异,区分环境效应或遗传效应,是了解其适应干旱机制和策略的重要基础。目前,在广西、贵州马尾松优良遗传型筛选的基础上,进一步研究其优良家系的抗旱特性,是评价其干旱适应性,确定将南带种源引入中带的重要依据。基于此,本研究通过盆栽试验,研究了持续干旱胁迫对来自广西和贵州的3个马尾松优良家系的形态、生长、生物量分配及生理等多方面的影响,采用隶属函数法评价马尾松的抗旱性,灰色关联法筛选出抗旱指标,为进一步探寻马尾松幼苗对干旱胁迫响应特性及抗旱机制提供科学的理论依据; 此外,初步分析了马尾松抗旱性与其遗传特性之间的关系,对马尾松优良抗旱家系的筛选和培育及干旱地区马尾松种质资源的发展与保护具有重要的实践意义。
1 材料与方法
1.1 试验材料与设计
试材GX01和GX02两个家系是由广西壮族自治区林业科学研究院选育,并经过广西自治区品种审定委员会审定的优良家系,其速生性和适生性很好,且适合向中亚热带引进。GZ01是产自贵州都匀马尾松国家良种基地优良家系的混种(文中一并简称家系)。2014年4月,将3个家系的1年生马尾松苗移栽至贵州大学林学院苗圃花盆内,花盆上口径300 mm,下口径200 mm,高250 mm; 土壤取马尾松林下黄壤(pH5.0~5.5),每盆土量一致,土壤田间持水量48.23%; 期间正常生长与管理1年,最后各选择长势一致的2年生幼苗(60 cm ± 5 cm),于2015年5月正式试验。采取随机区组设计,设对照组和干旱组,每组30盆,每盆3株。试验前给苗木浇透水,对照组每3天正常浇水,干旱组持续自然耗水30天,直至枯死。分别在试验的0,5,10,15, 20,25,30天进行生长测定,并取相同部位针叶测定生理指标。各取样时间点对应的大气温度分别为27,24,25,26,23,27,28 ℃; 土壤相对含水量分别为81.9%,71.4%,57.8%,46.6%,38.7%,30.1%, 20.6%,分属于湿润(75%~80%)、轻微干旱(55%~60%)、中度干旱(45%~50%)、重度干旱(30%~35%)和极端干旱(20%~25%)(王艺等, 2013)。
1.2 指标测定
1.2.1 生长指标 试验开始时(0天),每个家系各选10株苗木(每盆选1株),全株取样,烘干称重法测定苗木干质量,获得起始干质量; 干旱胁迫30天,每个家系对照组和干旱组各选10株苗木,相同方法获得最终干质量。干物质积累=最终干质量-起始干质量,根冠比=苗木地下干质量/苗木地上干质量。苗高: 皮尺测定从地径至顶芽基部的苗干长度,精确到0.1 cm,每10天测量1次,6次重复。地径: 电子游标卡尺测定土痕处的苗干直径,精确到0.01 mm,每10天测量1次,6次重复(每盆选1株为1次重复)。
1.2.2 生理指标 超氧化物歧化酶(SOD) 采用黄嘌呤氧化酶法,以每g组织在1 mL反应液中SOD抑制率达50%时对应的SOD量为1个酶活性单位,550 nm吸光值计算酶活性。过氧化物酶(POD)采用愈创木酚法,37 ℃下每mg蛋白每min催化1 μg愈创木酚的酶量为1个酶活性单位,420 nm吸光值变化计算酶活性。脯氨酸(PRO)含量用酸性茚三酮比色法,520 nm吸光值计算PRO含量。丙二醛(MDA) 含量采用硫代巴比妥酸法,532 nm吸光值计算MDA含量。上述指标均采用南京建成生物有限公司的试剂盒测定,测定过程及计算公式均参照试剂盒说明书的具体操作步骤进行。所有吸光值均由T6新世纪紫外可见分光光度计测定。每个指标3次重复(每盆选1株为1次重复)。
1.3 数据分析
采用SPSS20.0统计软件进行相关分析、方差分析和多重比较; 抗旱遗传特性分析参考周志春等(2004)、续九如(2006)的方法,抗旱效率=干旱胁迫干物质积累量 / 供水对照干物质积累量,家系遗传力h2=1-1/F(F为方差值); 模糊隶属函数(王艺等, 2013)评价3个家系抗旱能力,隶属函数值越高则表示抗旱性越强; 灰色关联法(黄承玲等, 2011)分析各指标与抗旱隶属函数均值的关联度,关联度越大则表示该指标与抗旱性关系越紧密。
2 结果与分析
2.1 马尾松生长指标
2.1.1 干旱胁迫对苗高的影响 正常水分下,3个家系苗高均随时间递增,家系间差异显著; 干旱胁迫下苗高显著低于正常水分下的苗高, 表明干旱胁迫抑制了苗高生长,且10天后抑制逐渐增强, 20天后苗高生长趋于停滞; 相同水分条件下,苗高为GX02>GX01>GZ01,GX02与GX01差异不显著,二者均显著高于GZ01,表明GX02的苗高生长受干旱抑制程度最小,GX01其次,GZ01最大(图1A)。
2.1.2 干旱胁迫对地径的影响 正常水分下,3个家系地径均随时间递增,家系间差异显著; 干旱胁迫下地径生长随时间先升后降, 20天后均低于正常水分下的地径,表明干旱胁迫抑制了地径生长,且10天后抑制逐渐增强; 相同水分条件下,地径为GZ01>GX01>GX02,GX01与GX02差异不显著,二者均显著低于GZ01,表明GZ01地径受干旱抑制的程度最小,GX01其次,GX02最大(图1B)。
2.1.3 干旱胁迫对干物质积累的影响 正常水分下,3个家系干物质积累量GX02(0.89 g)>GX01(0.83 g)>GZ01(0.75 g),家系间差异显著; 持续干旱30天后,各家系干物质积累GX01(0.35 g)>GX02(0.29 g)>GZ01(0.20 g),呈显著差异,分别比供水对照显著下降了57.83%,67.42%,73.33%,表明干旱胁迫下各家系干物质积累明显受到抑制,其中GZ01受影响程度最大,GX02次之,GX01最小(图2A)。
2.1.4 干旱胁迫对根冠比的影响 正常水分下,3个家系根冠比介于0.190~0.196之间,无显著差异; 持续干旱30天后,GX01、GX02显著大于供水对照,GZ01高于供水对照,但差异不显著,表明干旱胁迫引起生物量重新分配,生物量更多流向根部导致根冠比增大;干旱胁迫后的根冠比为GX01(0.298)>GX02(0.276)>GZ01(0.215),分别比供水对照显著提高了52.04%,42.27%,13.16%,GX01略高于GX02,但差异不显著,二者均显著高于GZ01, 表明GX01在干旱胁迫下具有较强的生物量分配能力(图2B)。

图1 持续干旱胁迫下马尾松苗高和地径变化Fig.1 The changes of height and collar diameter of Masson pine seedlings under continuous drought stress 不同大写字母表示各家系差异显著( P < 0.05)。下同。Different capital letters mean significant difference at 0.05 level in 3 Masson pine families. The same below.
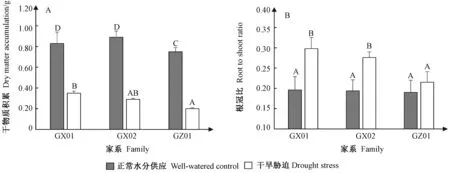
图2 持续干旱胁迫下马尾松苗干物质积累和根冠比变化Fig.2 The changes of dry matter accumulation and root to shoot ratio of Masson pine seedlings under continuous drought stress
2.1.5 生长指标与抗旱效率的关系 正常水分下,抗旱效率与苗高表现出极显著正相关,表明家系水平上马尾松可通过苗高初步判断其抗旱性状。持续干旱30天后,根干质量、根冠比、苗高、干物质积累较高的家系,其抗旱性相应较高; 而地径、茎干质量、总干质量等与其抗旱性的相关性不显著(表1)。
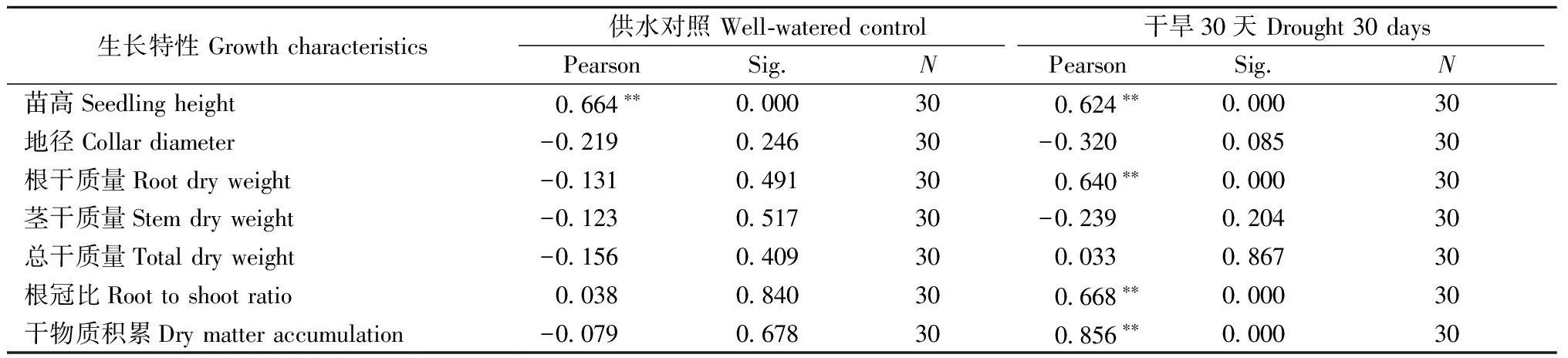
表1 马尾松家系抗旱效率与生长特性的相关分析①Tab.1 Correlation between drought resisting efficiency and its related growth traits of three Masson pine families
①**,在 0.01水平上显著相关Significant correlation at 0.01 level.
2.1.6 抗旱性相关指标的家系遗传力 正常水分下,苗高和干物质积累的家系遗传力估算值为0.997和0.882,表明其受较高水平的遗传控制; 而根干质量、根冠比的家系遗传力相对较低,表明正常水分下该指标在家系间的差异不显著,受遗传控制较小,更多由环境因素决定。持续干旱30天,根干质量、根冠比、苗高、干物质积累的家系遗传力估算值在0.937~0.998,均高于正常水分下各生长指标的家系遗传力,表明干旱胁迫下上述生长指标的抗旱性均受较高水平的遗传控制(表2)。
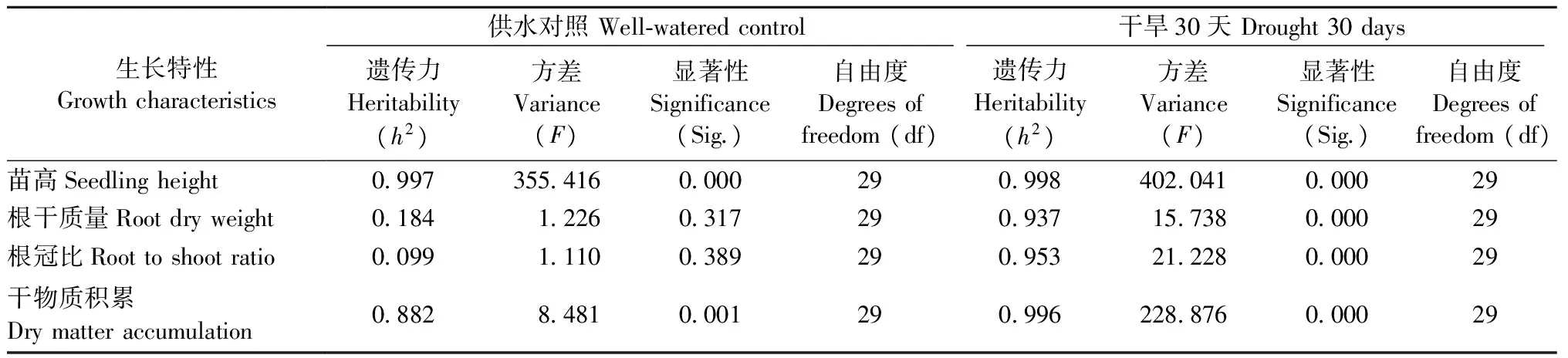
表2 不同干旱程度下马尾松抗旱相关生长指标的家系遗传力Tab.2 Family heritabilities of drought resisting growth traits of Masson pine families under different water levels
2.2 马尾松生理指标
2.2.1 干旱胁迫对MDA含量的影响 持续干旱下,3 个家系MDA含量整体呈增大趋势; 0~10天增速缓慢,差异不显著,10天后均显著上升,并于25~30天升达峰值,表明中度干旱胁迫下,细胞膜开始表现过氧化伤害,随干旱的持续伤害程度逐渐增大。其中,MDA含量增幅为GZ01>GX02>GX01,表明GZ01的膜质过氧化最严重。相同时间点,各家系间MDA含量差异显著; 5天时GZ01显著高于GX01和GX02,10~20天时 GX01均显著高于GX02和GZ01,25天各家系无显著差异,30天时GX01则明显下降,并显著低于GZ01和GX02,表明持续干旱胁迫后期,GX01抵御膜质过氧化伤害的能力大于GZ01和GX02(图3A)。
2.2.2 干旱胁迫对PRO含量的影响 持续干旱下,3 个家系的PRO含量均有明显积累; 0~10天时积累缓慢,无显著差异,15天时均迅速增大直至30天,该阶段各家系变化趋势不同: 15~30天时,GZ01的PRO含量维持在219.67~267.63 μg·g-1间,变幅不显著,GX02呈现先降后升的显著变化趋势,GX01则出现先升后降再升高的显著变化过程。其中,PRO含量增幅大小为GX01>GZ01>GX02,分别是胁迫0天值的14.4倍、11.3倍、10.6倍,表明GX01应对干旱胁迫的渗透调节能力强于GZ01和GX02。此外,各家系PRO含量达到峰值的时间不同,分别出现在20天(GX01的356.39 μg·g-1)、25天(GZ01的267.6 μg·g-1)、30天(GX02的281.23 μg·g-1),上述时间点家系间差异显著; 表明GX01的PRO对干旱胁迫的响应较GX02和GZ01迅速(图3B)。
2.2.3 干旱胁迫对SOD活性的影响 持续干旱下,3 个家系的SOD活性均呈现先升后降趋势; 其中,GZ01在干旱初期较敏感,5天时显著上升,10天时达峰值707.07 U·g-1(FW),随后持续下降,30天时SOD活性低于胁迫0天值; GX01和GX02在干旱初期变化不显著,10天时分别显著上升至峰值781.90 U·g-1(FW)和780.21 U·g-1(FW),显著高于GZ01峰值,15~25天呈显著下降,但仍表现出较高酶活性,30天时SOD活性亦高于胁迫0天值。整个胁迫过程中,增幅大小为GX01>GX02>GZ01。 0~10天,各家系间的SOD活性差异显著; GZ01由0天显著低于GX01和GX02上升为5天时显著高于GX01和GX02,表明GZ01的SOD活性对轻度干旱的响应比GX01和GX02迅速; 10~30天时GZ01持续低于GX01和GX02,表明中度、重度干旱时GX01和GX02的SOD活性相对较高(图3C)。
2.2.4 干旱胁迫对POD活性的影响 持续干旱下,3 个家系的POD活性呈现先升后降趋势,与SOD变化趋势一致; 其中,GX01的POD活性对干旱胁迫较敏感,5天时显著上升至309.67 U·g-1min-1,10天达峰值326.00 U·g-1min-1,表现出较高的酶活性,之后逐渐降低,25天仍高于胁迫0天值; GX02和GZ01的POD活性在10天时分别达到峰值296.25 U·g-1min-1和293.67 U·g-1min-1,但均低于GX01在轻度干旱(5~10天)时的POD活性,之后逐渐降低,25天时已低于胁迫0天值; 表明GX01在轻度、中度干旱下(5~15天)保持较高的POD活性,重度干旱下(20~30天)各材料表现一致。整个胁迫过程中,POD增幅为GX01>GX02>GZ01,但家系间的差异不显著(图3D)。
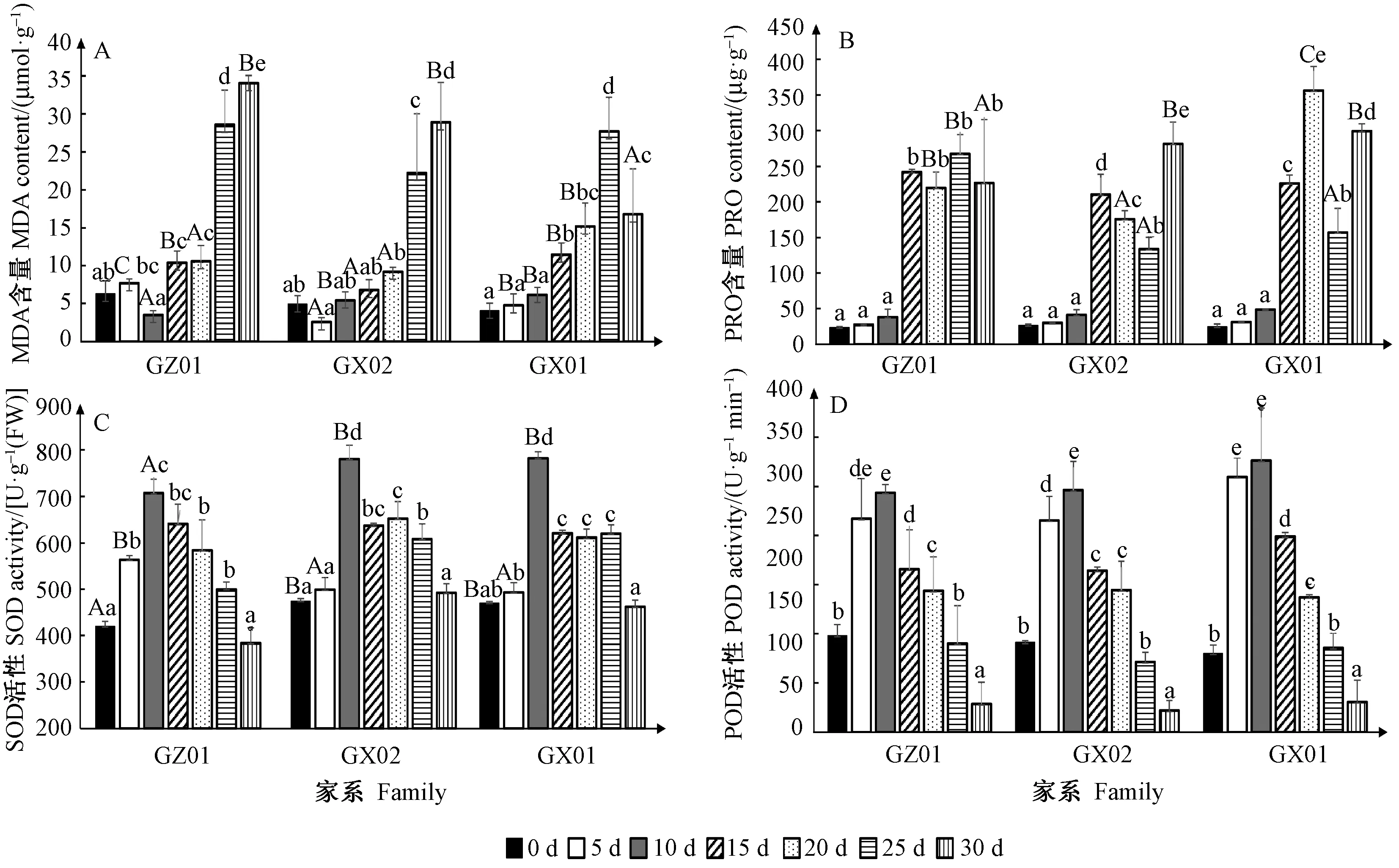
图3 持续干旱胁迫对MDA、PRO含量以及SOD、POD活性的影响Fig.3 The effects of drought stress on content of MDA and PRO, and activities of SOD and POD of three Masson pine families 不同大写字母表示同时间点不同家系间差异显著,不同小写字母表示同家系不同时间点差异显著(P< 0.05)。 Different capital letters mean significant difference at 0.05 level between different families in the same water level; and different lowercase letters mean significant difference at 0.05 level between different water levels in the same family.
2.2.5 各生理指标间的相关分析 生理指标间相关分析(表3)显示,POD与SOD极显著正相关(P<0.01),表明干旱胁迫下,抗氧化系统最快响应,SOD、POD均迅速升高; MDA、PRO均与POD极显著负相关(P<0.01),表明POD活性的增强可明显减缓膜质过氧化,膜质过氧化程度的加深反过来可抑制POD活性; 随着POD活性降低,机体启动后续防御,通过提高PRO含量,以增强渗透调节的方式继续抵御逆境伤害。可见,干旱胁迫下各生理指标间存在密切关联,马尾松的干旱应答是多种生理生化作用持续变化、协调互作的过程。
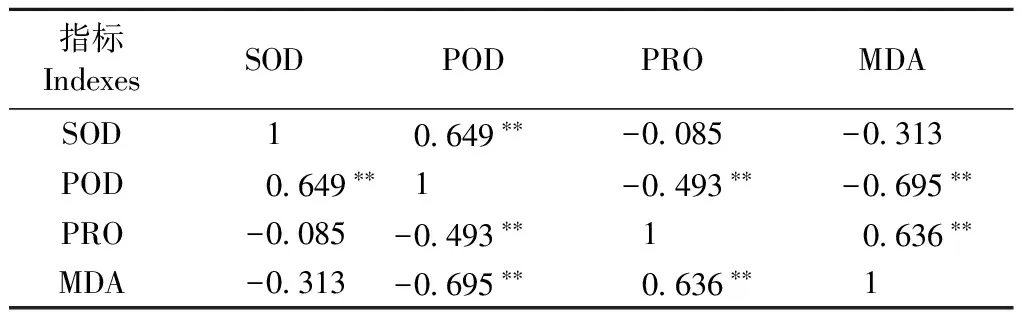
表3 马尾松各生理指标间的相关系数①Tab.3 Correlation between different physiological indexes of Masson pine
①**,在 0.01水平显著相关 Significant correlation at 0.01 level.
2.3 抗旱性综合评价
2.3.1 模糊隶属函数分析 对3个家系在干旱胁迫下形态、生物量、生理指标进行模糊隶属函数处理,各指标的隶属平均值越大,表示抗旱能力越强。表4显示,抗旱性大小为GX01>GX02>GZ01,表明3个家系对干旱的适应能力不同,并表现出不同抗旱特征。GX01和GX02抗旱能力远高于GZ01,但二者抗旱途径有差异,GX01在渗透调节能力及根生长上表现较好,而GX02则在保护酶系统及膜质抗氧化上表现较好。

表4 各家系马尾松抗旱指标的平均隶属函数值Tab.4 Synthetically subordinate function value of Masson pine seedlings
2.3.2 灰色关联分析 对各抗旱指标均值与抗旱系数间进行灰色关联分析,关联度越大表明该指标与抗旱性的关系越密切。表5显示,关联度大小为根冠比>苗高>根干质量>MDA>PRO>POD>SOD>干物质积累>地径。表明根冠比、苗高、根干质量等生长指标与抗旱性关联最大,其次为MDA、PRO,而POD、SOD、干物质积累、地径等对抗旱性影响最小。

表5 马尾松抗旱指标的灰色关联分析Tab.5 Grey relational analyses of drought resistance indexes of Masson pine seedlings
3 讨论
3.1 马尾松对干旱的响应机制及抗旱性评价
逆境下,植物活性氧(ROS)加速积累,超过一定阈值将引起膜质过氧化伤害,MDA作为膜质过氧化终产物,能反映膜系统损伤程度(Liuetal., 2011; Dounavietal., 2016)。本研究中,MDA含量在轻度、中度、重度干旱(0~20天)时上升缓慢,极重度干旱(25~30天)时大幅度上升,与王艺等(2013)的研究结果相似,表明马尾松在轻度、中度、重度干旱时具有较高的耐受性,在极重度干旱时其膜质过氧化严重而造成不可逆的膜系统损伤。相关分析表明,马尾松抗旱防御机制与启动氧清除系统、调节渗透物质密切相关,具有几种生理作用关联互作、协同抵御干旱逆境的抗旱特征。轻度干旱时(0~10天),SOD、POD均显著升高,MDA、PRO变化平稳; 说明干旱胁迫初期,苗木的保护酶系统最先响应,SOD、 POD活性大幅增加,体内活性氧得以清除,有效减少干旱引起的细胞过氧化损伤,因而MDA含量相应较低; 同时水分损失不明显,渗透调节物质PRO含量变化平缓; 该阶段几种生理过程共同响应,但氧清除系统占据主导作用。中度、重度干旱时(15~20天),SOD、POD下降显著,但仍维持较高水平,MDA、PRO显著上升,且POD、MDA与PRO之间存在显著的相关性; 说明干旱胁迫中期,MDA积累加速,反而制约SOD、POD活性,保护酶系统由积极保护转为消极抵御; 同时,水分明显亏缺,苗木迅速调整抗旱适应机制,通过加速积累PRO含量,主动降低体内渗透势,以促使细胞从外界吸收水分,维持细胞膨压,减轻植物的脱水伤害,提高自身的抗旱能力; 且PRO积累量越高,抗旱性越强 (Silvaetal., 2010)。极重度干旱时(25~30天),SOD、POD进一步下降,且低于胁迫0天的水平,MDA、PRO则进一步升高,说明干旱胁迫后期,水分严重亏缺,PRO持续大幅上升以维持机体活动; 同时,活性氧物质大量积累导致MDA急速骤升,SOD、POD活性受严重抑制而明显下降,最终膜结构破坏,抗旱能力减弱。此外,GX01和GX02在胁迫25天时其PRO随MDA的升高而降低,与水曲柳(Fraxinusmandshurica)(卫星等, 2009)的研究结果相似; 25天时,MDA骤然升高,膜结构破坏,引起机体代谢紊乱,修复能力及活力明显减弱,渗透调节作用相应受到抑制; 30天时PRO有所升高,表明GX01和GX02在极重度干旱胁迫下仍具有一定的生理响应及活动特性。可见,马尾松干旱早期主要以提高保护酶活性减少过氧化伤害,干旱中、后期主要以增强渗透调节减轻膜透性伤害,通过调节一系列适应性生理变化,抵御和适应干旱逆境。
生物量是植物生长发育的物质基础,干旱胁迫下,植物通常会调整生长方向和生物量分配策略应对逆境伤害; 根系作为最先感应土壤水分亏缺的器官,在干旱胁迫时常表现出适应性变化(Finéretal., 2011; 卫星等, 2009)。研究发现,随着干旱胁迫加深,3个家系根冠比均高于对照,高径生长和干物质积累均不同程度受到抑制; 与种源水平马尾松(胡晓健等, 2010)、喜树(Camptothecaacuminata)(应叶青等, 2012)等的研究结论相似。干旱胁迫下,GX01和GX02的根冠比显著高于对照,苗高、地径均显著低于对照,表明二者均选择暂缓地上部分生长,将生物量更多分配给地下根系,促进根生长以吸收更多水分,抵抗土壤干旱引起的水分缺失,表现出积极的抗旱性; GZ01根冠比显著低于GX01、GX02,虽略高于其供水对照,但差异不显著,反映出GZ01生物量重新分配的能力较弱,相比GX01和GX02,其生物量更多流向地上部分,即选择茎叶生长以避开干旱逆境带来的生长停滞,表现出消极的避旱反应。整体而言,GX01干旱胁迫下的干物质积累最多(0.35 g),随水分增加干物质积累的增幅最小(0.48 g),属于对干旱敏感度较低的耐旱型家系; 虽然GZ01干物质积累最少(0.20 g),但正常水分下增益最大,其干物质积累是干旱下的3.75倍,高于GX01(2.37倍)和GX02(3.07倍),应属于对水分敏感的干旱敏感型家系。
上述研究显示,马尾松在形态、生长及生理方面均对干旱胁迫表现出明显响应。灰色关联分析得出,各指标抗旱关联度大小为根冠比>苗高>根干质量>MDA>PRO>POD>SOD>干物质积累>地径,即生长指标是衡量3个家系抗旱性的首要指标,其次为生理指标; 与杨敏生等(2002)等的观点相似。水分是幼苗生长的限制因子,苗高、生物量等对水分变化非常敏感,特别在每年5—6月的幼苗快速生长期,这种敏感程度愈加凸显,更能直观反映水分缺失对马尾松的影响; 与生长指标相比,生理指标的遗传变异较小(杨敏生等, 1997),尽管在某些胁迫点上家系间的生理指标差异显著,但更多表现为植物内在的瞬时变化,同一物种生物学特性决定了家系间的SOD、 POD、MDA、 PRO的变化趋势大体一致; 因此评价马尾松抗旱性,生长指标优于生理指标。此外,本研究得出SOD与马尾松抗旱性的关联度较低,与王艺等(2013)的研究结论不同; 本研究中,SOD对干旱最先响应,在抵御干旱过程中发挥重要的先导作用,但试验的3个家系SOD活性均保持较高水平,且总体上家系间差异不显著,基于SOD难以判断各家系抗旱性大小; 而王艺等(2013)研究接菌与不接菌马尾松的抗旱性,外源菌作用容易引起马尾松SOD的显著变化,其抗旱表现亦随之不同。因此,试验材料及评价系统的差异可能是研究结论不同的主要原因。此外,MDA作为膜损伤的监测指标,与马尾松抗旱关联度最大,可作为评价马尾松抗旱性的重要指标之一。综上所述,植物应对干旱逆境过程是一个复杂的网络调控过程,受形态、生理、细胞、分子等复合遗传性状的综合影响,单一指标难以真实反映植物的抗旱能力; 基于隶属函数法,本研究得出3个家系抗旱性大小为GX01> GX02> GZ01。
3.2 马尾松抗旱性与遗传特性的关系
植物的抗旱性是遗传变异和自然选择的结果,与植物的形态、结构、生长等密切相关(吴丽君等, 2015; 任磊等, 2015)。从马尾松抗旱性状的家系遗传力来看,苗高、干物质积累量、根干质量及根冠比都受较高水平的遗传控制,说明遗传因素对抗旱性状的影响占主导地位。干旱胁迫下,GX02和GX01的高径生长、根冠比变化相似,均与GZ01差异显著; 可见,GX01和GX02在应答干旱时其形态、生长、生物量的变化趋势相似,且表现出较强的抗旱性,与抗旱性较差的GZ01存在明显差异。因此,3个家系抗旱性的差异是由遗传差异所决定的。研究组发现,受地理距离差异的影响,广西的GX01与GX02遗传距离较近,均与贵州的GZ01遗传距离较远(杜明凤等, 2016)。形态观测发现,贵州的GZ01具有明显的生长停顿,而广西的GX01和GX02没有明显生长停顿; 苗高均为20~25 cm的1年生幼苗长至2年时,GX02(62~67 cm)和GX01(60~65 cm)苗高接近,明显高于GZ01(50~58 cm),符合马尾松随纬度增加苗高生长渐降的规律(秦国峰等, 2012),表明苗高生长差异与纬度相关,印证了马尾松遗传差异与地理差异有关(杜明凤等, 2016)。由此推断,马尾松抗旱性受遗传因素控制,家系间显著的抗旱差异由其遗传差异决定,是在不同地理环境长期“驯化”下所形成的不同生存或适应机制。
4 结论
持续干旱胁迫下,马尾松通过减缓地上苗木生长、增加地下根部生长来抵御干旱逆境; 干旱早期,马尾松增加保护酶的活性以延缓过氧化伤害,中后期则增加渗透调节物质减缓膜质损伤。基于形态、生长及生理等各项指标的分析,发现3 个马尾松家系的抗旱性差异明显,依次为GX01(广西)>GX02(广西)>GZ01(贵州); 其抗旱差异主要受遗传因素影响,可能与其不同的地理环境长期驯化有关。
丁贵杰,周志春,王章荣,等. 2006.马尾松纸浆用材树种培育与利用.北京:中国林业出版社,1-34.
(Ding G J, Zhou Z C, Wang Z R,etal. 2006. Cultivation and utilization of pulpwood stand forPinusmassoniana. Beijing: China Forestry Publishing House,1-34. [in Chinese])
杜明凤,丁贵杰. 2016.不同种源马尾松ISSR遗传结构及影响因素分析.广西植物,36(9):1068-1075.
(Du M F, Ding G J. 2016. Analysis of genetic structure and its related factors ofPinusmassonianafrom different populations by ISSR marker. Guihaia, 36(9):1068-1075. [in Chinese])
胡晓健,欧阳献,喻方圆. 2010.干旱胁迫对不同种源马尾松苗木生长及生物量的影响.江西农业大学学报,32(3): 510-516.
(Hu X J, Ouyang X, Yu F Y. 2010. Effects of drought stress on the growth and biomass ofPinusmassonianaseedlings in different provenances. Acta Agriculturae Universitatis Jiangxiensis, 32(3): 510-516. [in Chinese])
黄承玲,陈 训,高贵龙. 2011.3种高山杜鹃对持续干旱的生理响应及抗旱性评价.林业科学, 47(6):48-55.
(Huang C L, Chen X, Gao G L.2011. Physiological response of seedlings of three Azalea species of drought stress and evaluation of drought resistance. Scientia Silvae Sinicae, 47(6):48-55.[in Chinese])
康宗利,杨玉红,张立军. 2006.植物响应干旱胁迫的分子机制.玉米科学, 14(2): 96-100.
(Kang Z L, Yang Y H, Zhang L J. 2006. Molecular mechanism of responding to drought stress in plants. Journal of Maize Sciences, 14(2): 96-100. [in Chinese])
秦国峰,周志春.2012.中国马尾松优良种质资源.北京:中国林业出版社,1-41.
(Qin G F, Zhou Z C. 2012. Germplasm Resources of Chinese Masson Pine. Beijing: Chinese Forestry Publishing House, 1-41. [in Chinese])
任 磊,赵夏陆,许 靖, 等. 2015.4种茶菊对干旱胁迫的形态和生理响应.生态学报,35(15):5131-5139.
(Ren L, Zhao X L, Xu J,etal.2015.Varied morphological and physiological responses to drought stress among four teaChrysanthemumcultivars. Acta Ecologica Sinica, 35(15):5131-5139. [in Chinese])
王林龙,李清河,徐 军,等. 2015. 不同种源油蒿形态与生理特征对干旱胁迫的响应.林业科学, 51(2): 37-43.
(Wang L L, Li Q H, Xu J,etal. 2015. Morphology and physiology characteristic responses of different provenances ofArtemisiaordosicato drought stress. Scientia Silvae Sinicae, 51(2): 37-43. [in Chinese])
王 艺,丁贵杰. 2013.马尾松菌根化苗木对干旱的生理响应及抗旱性评价.应用生态学报, 24(3):639-645.
(Wang Y, Ding G J. 2013. Physiological responses of mycorrhizalPinusmassonianaseedlings to drought stress and drought resistance evaluation. Chinese Journal of Applied Ecology, 24(3):639-645. [in Chinese])
卫 星,王政权,张国珍,等. 2009.水曲柳苗木不同根序对干旱胁迫的生理生化反应. 林业科学,45(6):16-21.
(Wei X, Wang Z Q, Zhang G Z,etal. 2009. Physiological and biochemical responses of different order roots inFraxinusmandshuricaseedlings to drought stress. Scientia Silvae Sinicae, 45(6): 16-21.[in Chinese])
吴丽君,李志辉,杨模华,等. 2015.赤皮青冈幼苗叶片解剖结构对干旱胁迫的响应.应用生态学报, 26(12):3619-3626.
(Wu L J, Li Z H, Yang M H,etal.2015. Response of leaf anatomical characteristics ofCyclobalanopsisgilvaseedlings to drought stress. Chinese Journal of Applied Ecology, 26(12):3619-3626. [in Chinese])
续九如. 2006.林木数量遗传学.北京:高等教育出版社, 1-117.
(Xu R J. 2006. Trees quantitative genetics. Beijing: Higher Education Press, 1-117. [in Chinese])
杨敏生,裴保华,朱之悌. 1997.水分胁迫下白杨无性系生理和生长的数量遗传分析.北京林业大学学报,19(2):50-56.
(Yang M S, Pei B H, Zhu Z T. 1997. Quantity genetic analysis of the physiological and growth properties for white poplar hybrid clones. Journal of Beijing Forestry University, 19(2):50-56 [in Chinese])
杨敏生,裴保华,朱之悌. 2002. 白杨双交杂种无性系抗旱性鉴定指标分析.林业科学,38(6):36-42.
(Yang M S, Pei B H, Zhu Z T. 2002. Index analysis on comprehensive judgement of drought resistance ability of white poplar bybrid clones. Scientia Silvae Sinicae, 38(6):36-42. [in Chinese])
应叶青,刘 鹏,王兴华,等. 2012.干旱对不同种源喜树苗木生长及叶片喜树碱产量的影响. 林业科学, 48(11):30-35.
(Ye Y Q, Liu P, Wang X H,etal. 2012. Effect of drought stress on the seedling growth from different provenances and camptothecin yield in leaves ofCamptothecaacuminata. Scientia Silvae Sinicae, 48(11):30-35. [in Chinese])
周志春,谢钰容,金国庆,等. 2004.马尾松磷效率及相关性状的家系遗传和变异.北京林业大学学报,26(6):1-5.
(Zhou Z C, Xie Y R, Jin G Q,etal.2004.Inheritance and variation of phosphorus efficiency and its related traits in families ofPinusmassoniana. Journal of Beijing Forestry University, 26(6):1-5. [in Chinese])
Amrita G D, Keith T K, Walter G W. 2004. Plant water relations and photosynthesis during and after drought in a Chihuahuan desert arroyo. Journal of Arid Environments, 59(1):27-39.
Antonino D I, Antonio M, Gabriella S S,etal. 2011.Fine root growth ofQuercuspubescensseedlings after drought stress and fire disturbance. Environmental and Experimental Botany, 74(2):272-279.
Dounavi A, Netzer F, Celepirovic N,etal. 2016. Genetic and physiological differences of European beech provenances (F.sylvaticaL.) exposed to drought stress. Forest Ecology and Management, 361(2):226-236.
Fang Y J, Xiong L Z.2015. General mechanisms of drought response and their application in drought resistance improvement in plants. Cellular & Molecular Life Sciences, 72(4):673-689.
Finér L, Ohashi M, Noguchi K,etal. 2011. Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management, 261(2):265-277.
Liu C C, Liu Y G, Guo K,etal. 2011. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in Karst habitats of southwestern China. Environmental and Experimental Botany, 71(6):174-183.
Majdi H, Andersson P. 2005. Fine root production and turnover in a Norway spruce stand in northern Sweden: effects of nitrogen and water manipulation. Ecosystems, 8(2): 191-199.
Shulaev V, Cortes D, Miller G,etal. 2008. Metabolomics for plant stress response. Physiologia Plantarum, 132(2):199-208.
Silva E N,Ferreira-Silva S L, Viégas R A,etal. 2010. The role of organic and inorganic solutes in the osmotic adjustment of drought-stressedJatrophacurcasplants. Environmental and Experimental Botany, 69(3): 279-285.
(责任编辑 徐 红)
Responses to Continuous Drought Stress and Drought Resistance ofDifferent Masson Pine Families
Du Mingfeng1,2Ding Guijie1Zhao Xizhou1
(1.ResearchCentreforForestResourcesandEnvironmentofGuizhouProvinceCollegeofForestry,GuizhouUniversityGuiyang550025; 2.SchoolofKarstScience,GuizhouNormalUniversityGuiyang550001)
【Objective】In this study, we investigated effects of continuous drought stress on growth and physiological characteristics of Masson pine (Pinusmassoniana) and compared drought-resistances of 3 Masson pine families, to elucidate the mechanisms of drought resistance and their genetic effects, and to provide a theoretical basis for breeding drought-resistant families.【Method】Pot experiment with random block design in greenhouse was conducted to investigate the effect of drought stress on growth, osmoregulation and antioxidants of 2-year-old Masson pine seedlings, respectively from Guizhou and Guangxi provinces.【Result】With the increase of drought stress intensity, the activities of protective enzyme SOD of the three Masson pine families displayed an increase in the beginning and followed by a decrease, and significant difference in SOD activity among the three families was found (P<0. 05) during 0-10 days of stress; the activities of protective enzyme POD was found in the same tendency of SOD activities, but the difference in POD activity among the three families was not significant during the whole period of drought treatment (P>0. 05); MDA content displayed a trend of increase followed by a decrease in GX01 but a reversed trend in GX02 and GZ01, and significant differences were found in MDA content among the three families (P<0. 05) from the 5thto the 20thday of drought stress and on the 30thday of drought stress; the proline content of GZ01 was increased constantly, while the proline contents of GX02 and GX01 displayed a pattern of increase-decrease-increase, and proline contents were found significantly differences among the three families (P<0. 05) from the 20thto the 30thday of drought stress. Moreover, continuous drought restricts the growth of seedlings, the growth of height,and ground diameter and the accumulation of dry matters of the 3 families of Masson pine were all inhibited by the drought stress and varied significantly (P<0. 05 ) among the three families. Continuous drought led to a significantly larger root to shoot ratio of GX01 and GX02 compared to the control. The ratio of GZ01 was also larger than that of the control, but not to a significant level. In addition, subordinate function was used to rank their drought resistance and the order was GX01>GX02>GZ01. Root to shoot ratio, height, MDA and Pro were as the key indicators to evaluate the drought resistance of Masson pine.【Conclusion】 These results indicated that activities of antioxidant enzymes of Masson pine seedlings was increased to resist the damage of mild drought stress; and then, the content of osmotic substance was increased to prevent the damage of dehydration from moderate and severe drought stresses; at last, the membrane was seriously damaged with increasing of MDA contents under aggravation drought stress. Obviously, Masson pine seedlings adapted to drought stress by increasing underground growth and decreasing aboveground growth. We conclude that drought resistance of Masson pine was mainly controlled by genetic factors, and differences of its drought resistance maybe caused by the genetic difference which was formed by long-term domestication under different geographical and ecological environment.
Pinusmassoniana; drought stress; growth characteristics; physiological response; drought resistance; hereditary
10.11707/j.1001-7488.20170603
2016-07-28;
修回日期: 2016-08-26。
基金项目: 国家自然科学基金项目(31260183); 国家科技支撑课题(2015BAD09B0102); 贵州省重大专项课题 (黔科合重大专项字[2012]6001号)。
S718.43
A
1001-7488(2017)06-0021-09
*丁贵杰为通讯作者。
