植物组织培养技术在中药资源中的应用
2017-07-13王娟李金鑫李建丽高文远
王娟+李金鑫+李建丽+高文远


[摘要] 植物组织培养技术以其独特的优势被广泛的应用于中药资源领域,在中药资源保护方面发挥了重要的作用。该文综述了近年来植物组织培养的一些应用,包括植物组织培养生产药用植物活性化合物、遗传多样性分析、道地药材、诱导子应用、生物合成及转基因植物等。通过以上研究将进一步促进植物组织培养技术的发展,使其在中药资源领域发挥更大的作用。
[关键词] 植物组织培养; 中药资源; 诱导子; 生物合成
[Abstract] Plant tissue culture technology has been widely used in the field of traditional Chinese medicine(TCM) resources with its unique advantages, playing an important role in the protection of TCM resources. In this review, some applications of plant tissue culture were summarized, including production of active compounds by using plant tissue culture, genetic diversity analysis, Dao-di herbs, elicitor application, biosynthesis and transgenic plants. Through the above researches will promote the further development of plant tissue culture technology, making it play a greater role in the field of TCM resources.
[Key words] plant tissue culture; Chinese medicine resources; elicitor; biosynthesis
中药资源是我国中药产业的基石,随着我国中药工艺的快速发展,大型和超大型企业不断涌现、形成和发展,对我国的中药资源形成了很大的压力,近些年中药材濒危资源的不断增加,中药材价格的不断增长,足以表明我国资源保护和可持续利用工作的紧迫性和重要性。通过发展植物组织培养技术来解决中药材的资源问题,对我国具有特殊而且重要的意义。植物组织培养技术的应用可以体现在以下几个方面:①通过植物组织和器官培养生产活性成分,保护濒危和珍稀药用植物资源;②通过遗传多样性分析,对植物组织培养材料进行评价;③通过植物组织培养技术研究道地药材遗传机制和环境机制;④通过诱导子的添加提高培养物中活性成分含量;⑤利用植物组织培养物进行基因功能筛选、验证及遗传转化研究。本文将从以上几个方面综述近年来植物组织培养技术在中药资源领域中的应用。
1 植物组织培养生产药用植物活性化合物
药用植物细胞,毛状根和不定根培养技术是生产药用植物次级代谢产物的一种非常有价值的工具。因此,通过细胞培养生产次级代谢产物的植物将近1 000种,超过600种的活性成分直接由植物细胞培养提供。目前,100多种毛状根已经成功的被发根土壤农杆菌诱导[1],主要通过优化培养基和关键生理因素去增加毛状根培养的次级代谢产物的生成。目前,人参,三七,柴胡,甘草,丹参,雷公藤和许多种药用植物已经制定了不定根培养体系。目前,为了大量的生产次级代谢产物,细胞、不定根、毛状根已经成功地应用于大规模培养。例如,人参不定根培养已经达到3万L。金丝桃不定根已经达到500 L,紫锥菊不定根已经达到1 000 L[2]。韩国,CBN生物科技公司,每年生产大约40~45 t人参不定根,这是一个利用植物组织培养生产药品、食品和化妆品的成功范例。主要研究了培养条件的优化和诱导子的使用来提高次级代谢产物的含量。由WHO资助的半合成青蒿素已经被赛诺菲公司研制成功,其用发酵方法由单糖生产的青蒿酸在2013年已形成60 t左右产能。Phyton为成为世界领先的高品质紫杉醇生产商已进行了巨额投资。我国具有通过植物细胞发酵生产紫杉醇的巨大產能,杜绝了对紫杉树种植的依赖,也避免了他们与环境、可持续性、可靠性和质量稳定性相关的固有问题。近年来药用植物细胞,毛状根和不定根次级代谢产物的积累情况见表1。
近年来,药用植物细胞,毛状根和不定根培养受到了广泛关注,因为它提供了许多植物次级代谢产物。总的来说,药用植物的细胞,毛状根和不定根是逐年增加的,并且它的培养规模已经逐渐扩大。
2 遗传多样性分析
植物组织培养是植物快速大量增值的一种潜在工具,并且有利于控制重要次级代谢产物的生成。然而再生植株的遗传基因会发生一定程度的变异,从而造成植株化学成分的改变及其临床疗效的差异,影响组织培养的优点[40]。因此,必须保证组培植株基因的一致性,以确保植株的质量。用于监测遗传基因变化的方法有简单重复序列,随机扩增多态性DNA和相关序列扩增多态性。
简单重复序列通常形成具有具体基因,少许具体标记物的种族。随机扩增多态性DNA方法是一种PCR技术,随机扩增DNA片段,使用低于最低退火温度的核苷酸序列的单一引物。这个技术已经广泛用于种族分类,遗传作图和种系发生[41-42]。在吊灯花属快繁的研究中,在植株的直接芽器官发生,间接芽器官发生和母本植物之间比较基因稳定性,结果显示通过简单重复序列和随机扩增多态性DNA方法,直接芽器官发生的基因和母本植物是相似的,间接芽器官发生的随机扩增多态性DNA指纹图谱显示出低的变异率[40]。相关序列扩增多态性是简单的有效地生产高再生性和多功能性的全组基因片段。在人参悬浮细胞培养中,通过随机扩增多态性DNA方法在悬浮细胞与幼苗之间比较基因稳定性,结果显示悬浮细胞的基因和幼苗是相似的[43]。在拟南芥悬浮细胞培养中,通过简单重复序列方法来比较悬浮细胞和拟南芥植株的基因稳定性,发现悬浮细胞的基因和幼苗是相似的[44]。
3 道地药材
道地性归根结底是道地药材所拥有的基因型受到特定生境(道地产区)中环境因子诱导后表达的产物。因此,基因表达是道地药材研究的重要环节,而功能基因表达的调控是道地药材研究的目标之一[45]。揭示道地药材和非道地药材在基因表达方面的差异是道地药材功能基因研究的重要内容。由于基因表达对研究所用RNA材料的要求较高,且通常需要通过对比实验来完成,造成道地药材基因表达及调控实验目前主要集中在实验室中。由于细胞培养和组织培养周期较短,材料易于获得,均匀性较好,因此道地药材功能基因表达与调控研究多是在植物组织和细胞中开展[46]。
例如,黄新等[47]利用mRNA差异显示技术研究茉莉酸甲酯对红豆杉细胞mRNA表达的差异。李娟等[48]考察了碳源、氮源、有机成分对HBsAg转基因人参细胞生长和HBsAg表达量的影响。刘峻等[49]研究了真菌诱导子影响人参毛状根总皂苷的合成量。以上研究,虽然距离真正的实现道地药材功能基因的表达和调控尚有一段距离,但为道地药材功能基因表达和调控研究积累了思路和方法。
4 诱导子作用机制及其应用
4.1 诱导子的作用机制
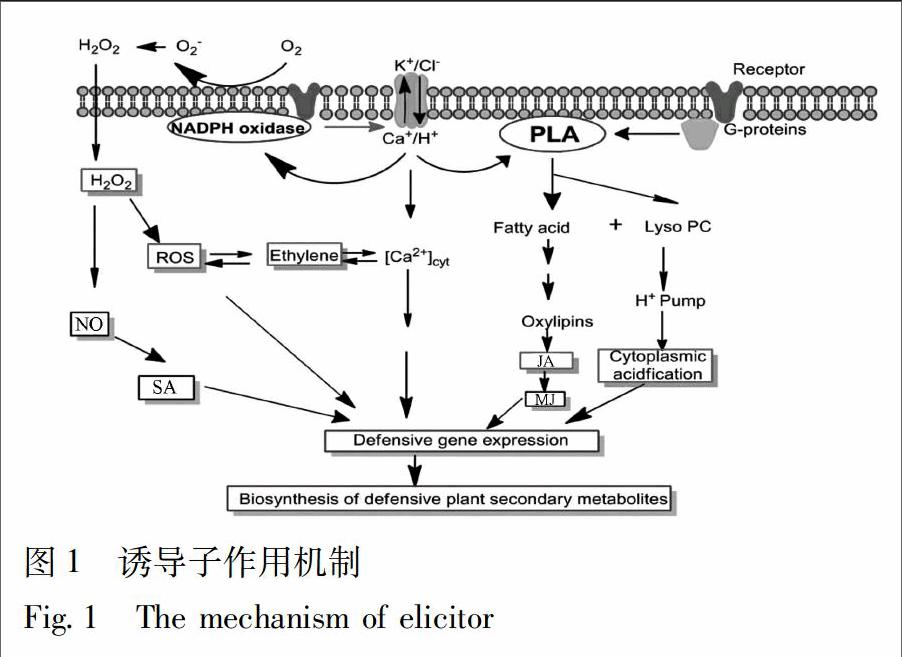
诱导子被植物细胞膜或细胞质中的接受体识别后,细胞膜便开始去极化,引起离子通道的开闭,如Ca2+从环境中流入细胞质,K+和Cl-流出,或者通过 G 蛋白偶联,激活第二信使系统,进一步的扩大这种效应,产生植物防御分子,如钙离子(Ca2+)、活性氧(ROS)、水杨酸(SA)、茉莉酸(JA)、一氧化氮(NO)、乙烯(ET)等,进而引起下游防卫基因的表达,改变相关酶的活性,并最终导致次级代谢产物的积累,见图1[50]。
4.1.1 细胞内信号分子和作用机制 植物细胞信号分子主要作用偶合各种胞内外刺激,包括生物或者非生物刺激,再通过细胞内信号传导系统使得刺激和细胞间信号级联放大,最终导致相关酶活性的改变,进而引起一系列生理生化反应。例如,诱导子刺激后,细胞质中钙离子(Ca2+) 浓度的升高、一氧化氮合成、活性氧迸发、茉莉酸合成途径激活等,诱导防御基因的开启,刺激代谢途径中关键酶的合成,最终促进植物次级代谢产物的合成[51]。
钙离子:诱导子刺激后,植物细胞在短时间内出现离子流,如Ca2+和H+从环境中流入细胞质,K+和Cl-流出等。这些Ca2+可以驱动钙离子受体钙调蛋白(CaM)的反应,一旦钙调蛋白激活后,可以进一步激活与钙调蛋白相关的蛋白激酶,从而引发蛋白的磷酸化,进而引起防御基因的表达[90]。例如,Ca2+含量迅速上升后,激活钙调蛋白,会使大量的NAD(P)H氧化酶被激活,NADPH氧化酶可以催化过氧化物的产生和积累,从而进一步引起并下游一系列的反应,刺激次级代谢物的积累[52]。Ca2+内流以后,可作为信号分子激活磷脂酸 C,进而使磷脂酰肌醇-4,5-二磷酸(PIP2)水解产生 2 个第二信使: IP3(三磷酸肌醇)和 DAG(甘油二酯)。其中,DAG 可以与蛋白激酶结合,激活蛋白激酶 C,从而触发蛋白磷酸化信号级联,引起细胞内代谢物的积累。IP3 可以与细胞内钙库(如内质网、线粒体、叶绿体、液泡等)上的钙离子通道受体结合,进而动员细胞内钙库中钙离子的释放。IP3-Ca2+信号途径被认为是诱导植物抗毒素产生的调节物质[53]。在长春花悬浮细胞过程中加入真菌粗提物作为诱导子时,IP3信号转导途径被激活,从而进一步使长春花细胞中生物碱的含量提高[54]。
活性氧:植物防御早期的另一个重要变化是活性氧的迸发现象。活性氧浓度升高可以作为第二信使引发胞内一系列抗性反应,如可以促进结构蛋白和木质素的合成使细胞壁增厚,细胞超敏死亡等,另外,活性氧介导的脂质氧化可以刺激茉莉酸及相关化合物的合成,诱导次级代谢防御基因和次生代谢物合成基因的表达,进而诱导次级代谢[55]。在红豆杉细胞培养过程中,经尖孢镰刀菌的细胞壁粗提物诱导后,红豆杉细胞中活性氧大量积累,最终会导致细胞中紫杉醇的含量显著上升,与对照相比提高了近4倍[56]。但是高浓度的活性氧,如 O2-,H2O2,OH-,1O2会损伤细胞内的成分。抗氧化酶体系,如超氧化物歧化酶 (SOD)、过氧化物酶 (POD)以及过氧化氢酶 (CAT)等,会被激活,从而减轻活性氧的毒性作用,增强植物的耐受性[57]。在丹参细胞培养过程中,Dong等[58]发现在丹参细胞培养基中加入水杨酸,酚类化合物的含量会显著增加,同时抗氧化物酶SOD,POD,CAT 的活性也会增加。
茉莉酸类:在许多的植物细胞培养体系中,诱导子都可以诱导内源的茉莉酸类物质的合成,其可作为细胞内和细胞间的信号分子,与转录因子相互作用,进而调节防御基因的表达,最终导致次生代谢物质的合成[59]。茉莉酸类也可以刺激次级代谢产物合成相关的基因的表达,从而促进一系列次级代谢产物的合成,包括萜类物质、黄酮类物质、生物碱和苯丙烷类物质[50]。在贯叶连翘细胞悬浮培养过程中,在培养基中加入黑曲霉细胞壁的粗提物作为诱导子时,茉莉酸的合成量迅速增加,同时细胞中金丝桃素的含量也迅速增加,而加入茉莉酸合成抑制剂后,细胞中金丝桃素的含量显著下降[57]。
一氧化氮和水楊酸:在自然界中,水杨酸(SA)是植物与病原菌相互作用的诱导物,它能够诱导发病机制相关 (PR) 基因的表达及和致病相关蛋白的产生,但是,水杨酸不是存在于所有植物防御代谢中[1]。研究发现,在部分植物的细胞中,水杨酸的迅速合成,可以诱导一些与代谢相关基因的表达,促使一些次生代谢产物的积累,例如,在茜草的培养过程中,SA可以增加细胞中蒽醌类物质的积累[60]。一氧化氮(NO)作为信号化合物,近年来才逐渐被认识。在植物体内,NO可以通过一氧化氮合成酶(NOS)、硝酸盐还原酶(NR)或者非酶促反应来合成[59]。转录组测序的结果表明,NO处理后,可以引起植物细胞内与压力和抗病性相关的基因的表达,说明了NO信号途径可能与细胞内的次生代谢相关[61]。另外,NO还可以作用于水杨酸(SA)等信号分子的上游,参与和调控植物细胞中SA等信号分子的生物合成[62]。
其他信号分子:除了上述信号分子外,乙烯和脱落酸都是植物的内源性激素,参与调节植物细胞内生长和发育的多种生理活动,如生长、衰老、植物应激反应等[50]。此外,乙烯、脱落酸等也在诱导子诱导的抗性反应中起重要作用,在不同浓度下它们从抑制和促进两方面影响次生代谢产物的积累[53]。例如,在加拿大红豆杉的培养过程中,乙烯可以促进低聚糖和茉莉酸甲酯对红豆杉细胞的诱导作用,增加紫杉醇的积累[63]。但是在丹参毛状根培养中,加入乙烯的抑制剂,反而会促进细胞中3种丹参酮的合成[64]。
4.1.2 转录因子 转录因子也称为反式作用因子,是指能够与真核基因启动子区域中顺式作用因子特异性结合的 DNA 结合蛋白。通过蛋白与基因之间以及蛋白与蛋白之间的相互作用,激活或抑制转录[65]。
诱导子刺激植物以后,往往是多种信号途径构成一个相互交错的工作网共同起作用,它们通过相互协调,刺激次生代谢物质的合成。其中,转录因子可以对多个信号途径进行整合,激活防御基因,使得宿主关闭一些基因表达而开启另一些基因表达的信号传递和调控,产生特定的次级代谢产物[66]。例如,真菌诱导子或者茉莉酸甲酯诱导后,可以引起细胞中AP2/ERF转录因子家族的转录因子的迅速积累,然后AP2/ERF转录因子识别的顺式元件GCCbox,从而激活乙烯响应基因的表达,进而激活下游异胡豆苷合酶的基因和生物碱合成相关的基因的表达,最终促进生物碱的积累[67]。
4.1.3 抗性基因的表达 诱导子与植物受体的相互识别后,第二信使的形成,会使抗性相关基因激活,从而相应的mRNA累积及相应酶的才会增加,最后促进次级代谢物质的产生。
目前研究得比较多并且己经得到克隆的抗性基因主要有:病程相关蛋白P(R)的基因如几丁质酶、β-1,3-葡聚糖酶等基因; 木质素合成相关酶的基因如PAL,PO等的基因; 富含轻脯氨酸糖蛋白和富含甘氨酸糖蛋白的基因; 钙离子依赖蛋白激酶的基因;谷胱甘肽S-转移酶基因等。研究发现,许多植物防御反应基因都是诱导表达的[68]。例如,在草莓的培养过程中,Landi等[69]考察了殼聚糖、苯并噻二唑和有机酸对草莓细胞中抗性相关基因表达量的影响,结果表明,3种诱导子都可以激活与钾离子通道、聚半乳糖醛酸酶、β-1,3-葡聚糖酶相关的抗性基因的表达,另外,苯并噻二唑也可以上调与钙离子依赖蛋白激酶、β-1,4-葡聚糖酶、抗坏血酸过氧化酶、谷胱甘肽-S-转移酶相关的抗性基因的表达量。
4.2 诱导子在组织培养中的应用
4.2.1 诱导子的定义与分类 从植物病理学方面来讲,诱导子是一种可以引起植物自身产生抗病反应以产生抗毒素(植保素)和过敏反应来保护自己的化学物质或生物因子;从植物组织培养方面来讲,诱导子是一种能促进植物细胞产生目标代谢物以及能引起某一组织内生理变化的化学物质或生物因子[70]。根据来源,诱导子可分为生物诱导子和非生物诱导子。生物诱导子主要包括真菌、细菌、病毒的灭活菌体,细胞粗提物以及某种成分、菌体分泌物以及当病原体或害虫侵染植物时植物的分泌物等。非生物诱导子主要包括金属离子、植物内源激素(茉莉酸、水杨酸、茉莉酸甲酯、脱落酸等)、化合物、理化因素(紫外光、温度、超声、水分、pH、盐强等)、气体(臭氧、一氧化氮、过氧化氢、二氧化碳、乙烯等)。
利用诱导子来提高次级代谢产物含量是目前在药用植物组织培养上常用的方法。其中最常用的是生物诱导子和非生物诱导子[70]。
4.2.2 生物诱导子在植物组织培养中的应用 生物诱导子中研究最多与最广泛的为真菌诱导子。1968年,Crutckshand等发现首个真菌诱导子Monilinia fructicola(褐腐病菌)菌丝体中的一种小分子多肽Monilicolin A能促进菜豆内表皮的形成与菜豆素的积累[71]。从此真菌诱导子受到越来越多的关注。
真菌诱导子主要包括真菌灭活菌体、真菌活菌、真菌菌体成分、真菌分泌物(培养液)等。在化学成分上主要包括多糖、低聚糖、多肽、蛋白、不饱和脂肪酸等。
真菌灭活菌体诱导子是指将真菌的菌丝体或孢子收获后,用蒸馏水清洗干净,放入烘箱烘干后,用研钵研成粉末,加入适量蒸馏水后放入灭菌锅中121 ℃灭菌25 min。菌丝体或孢子发生降解或部分降解。一般将灭活菌体溶液在一定时间加入植物培养物培养基,处理一定时间来诱导植物培养物的生长以及次级代谢产物的积累。真菌灭活菌体作为诱导子在植物组织培养中的应用,见表2。
真菌活菌作为诱导子是指将一定数目的真菌孢子加入植物组织培养物中。Awad等[86]认为,体外培养植物与栽培植物相比,体外培养通常是无菌环境,缺少与植物共生和非共生的土壤微生物,这可能造成体外培养植物细胞中次级代谢产物含量低的原因。因此,在Taverniera cuneifolia的培养过程中,考察了用5种真菌(黑曲霉、曲霉、青霉、串珠镰刀霉和冻土毛霉)和5种细菌(噬胺芽孢杆菌、发根农杆菌、致瘤农杆菌、蜡样芽孢杆菌和豆科根瘤菌)活菌为诱导子,对不定根中的甘草酸含量的积累,结果表明所有诱导子都可以提高不定根中甘草酸的含量,其中加入冻土毛霉后,甘草酸质量分数为4.90 mg·g-1,加入致瘤农杆菌后,甘草酸达到了6.37 mg·g-1,均高于空白组(1.46 mg·g-1)和茉莉酸甲酯(2.57 mg·g-1)处理组。
真菌菌体成分作为诱导子是指菌体中的某一成分作为诱导子加入植物组织培养物中。周立刚等考查了Fusarium oxysporium Dzf17菌低聚糖对盾叶薯蓣悬浮细胞的影响。首先考察了胞外多糖(EPS),水提菌丝体多糖(WPS)、氢氧化钠提取的菌丝体多糖(SPS)的添加时间以及多糖浓度对盾叶薯蓣悬浮细胞生长以及细胞中薯蓣皂苷元含量的影响。结果显示,WPS的效果最好,在细胞培养第25天时加入20 mg·L-1的WPS可以使细胞生物量、薯蓣皂苷元的含量和产量分别达到空白组的1.34,2.85,3.83倍[87]。接着周立刚等又考察了WPS中低聚糖DP4,DP7,DP10的的添加时间、多糖浓度以及添加次数对盾叶薯蓣悬浮细胞中薯蓣皂苷元含量的影响。结果显示,在细胞培养分别在第24,26天时连续加入6 mg·L-1 DP7,在第30天时收获,薯蓣皂苷元的含量和产量分别为空白组的9.19,12.38倍比单次加入刺激的效果好[88]。此外周立刚等还考察了分别来自于EPS,WPS,SPS的低聚糖EOS,WOS,SOS作用不同时间段对盾叶薯蓣悬浮细胞中防御相关酶(PAL,POD,PPO)活性的影响,结果显示EOS,WOS都能显著增加防御相关酶的活性,说明内生菌F. oxysporium Dzf17中的低聚糖可能激活和增强盾叶薯蓣悬浮细胞的防御机制[89]。真菌菌体成分作为诱导子在植物组织培养中的应用见表3。
真菌分泌物誘导子是指将真菌的发酵液收集后,用4层纱布进行过滤,1万×g 离心20 min后,用0.4 μm的滤膜进行过滤,用旋转蒸发仪把过滤后的发酵液浓缩到一定体积,放入灭菌锅中121 ℃灭菌25 min。一般将灭活菌体发酵液在一定时间加入植物培养物培养基,处理一定时间来诱导植物培养物的生长以及次级代谢产物的积累。真菌分泌物体作为诱导子在植物组织培养中的应用,见表4。
4.2.3 非生物诱导子在植物组织培养中的应用 水杨酸(SA)和茉莉酸类化合物 [茉莉酸(JA),茉莉酸甲酯(MJ)]由于可以通过诱导多种植物次级代谢产物合成途径中基因的表达来促进次级代谢产物含量的增加而广为人知。除此之外,由于他们可以在低浓度诱导远离他们合成部分的细胞反应,又被成为植物激素。其中,茉莉酸甲酯类应用最广泛,目前已经成功应用于人参、柴胡、百部、甘草、西洋参、匙羹藤、柴胡、紫锥菊、雷公藤、睡茄、刺五加、苦艾和积雪草等植物组织培养中来提高细胞中次级代谢物的含量。
在苦艾悬浮细胞培养中,添加1.0 mg·L-1的茉莉酸以及茉莉酸甲酯可以使其总酚,总黄酮的含量增加,另外,可以提高自由基清除能力[97]。加入10 mg·L-1的茉莉酸甲酯到人参不定根的培养液中,人参皂苷约32 mg·g-1是空白组的4.76倍[98]。
化学合成诱导子一般指的是对已知的诱导子进行改造,通过化学合成,在诱导子分子结构上加入一些官能团等,来提高诱导子的诱导能力,增加药用植物细胞中次级代谢物的积累。其中,最常见的是对茉莉酸甲酯的改造。例如,在三七悬浮细胞的培养过程中加入五氟丙基茉莉酸甲酯、2-羟乙基茉莉酸甲酯以及2-羟基乙氧基乙基,结果发现他们都能显著增加人参皂苷的含量[99]。Qian等[100]考察了2-羟乙基茉莉酸甲酯和三氟乙基茉莉酸甲酯对红豆杉细胞中次级代谢产物的诱导作用,结果发现,2-羟乙基茉莉酸甲酯和三氟乙基茉莉酸甲酯能显著提高红豆杉细胞中紫杉烷C的含量,质量分数分别达到44.3,39.7 mg·g-1,均高于对照组(茉莉酸甲酯处理组)的含量(质量分数分别为14.0,32.4 mg·g-1)。
除了常用的上述这些化合物外,还有一些物理因子如紫外光、温度、超声,金属离子,水分,pH,盐强,气体如臭氧、一氧化氮、过氧化氢、二氧化碳、乙烯等都能提高次级代谢产物的含量。
5 生物合成生产药用植物活性成分
由于生长受到自然环境的影响,许多药用植物生长缓慢或栽培机制复杂。中药材中的活性成分存在含量低、结构复杂、不稳定、很难化学合成或产率地下等缺点。利用生物技术生产活性成分有许多优点:包括生长周期短、生产标准化、质量稳定等,为中药资源保护提供了新的途径。随着生物合成技术在青蒿素、紫杉醇,丹参酮和人参皂苷生物合成研究中的应用,预测生物合成技术将成为中药可持续利用的重要途径之一。近年来利用生物合成来生产活性成分的例子见表5。
药用植物有效成分的生物合成是在基因组学的研究基础上,通过在细胞中构建有效成分的生物合成途径和代谢网络,从而实现药用有效成分的定向、高效的合成,以解决药用有效成分的生产及药物研发等一系列问题[52]。植物组织培养在生物合成中的应用包括功能基因的挖掘和基因功能体内验证。
基因功能基因的挖掘主要是通过转录组测序来实现的。Subramaniyam等[101]对MJ处理不同时间的人参不定根进行了转录组测序,发现30个转录因子,11个GT与人参皂苷合成有关。Nuruzzaman等[102]对MJ处理过的人参不定根以及未处理过的不定根进行测序,发现48个转录因子与人参皂苷的合成相关。Jayakodi等[103]对2种人参材料诱导出的不定根以及栽培人参进行了转录组测序,发现不定根中90%的基因与数据库得到一致,17%的基因是栽培品中所没有的,此外还发现了一些与人参皂苷合成相关的候选功能基因。
基因功能的体内验证主要是通过在植物组织培养器官体内进行基因沉默以及过表达来实现的。Park等[104]通过把CYP716A53v2基因在PCR 8.0中进行克隆,然后与pH2WG连接构建过表达载体,然后导入农杆菌中,侵染人参胚状体,经过筛选得到CYP716A53v2过表达根系;另一方面,把CYP716A53v2基因进行沉默,构建相应的沉默载体导入农杆菌中继而侵染人参胚状体,经过筛选得到CYP716A53v2沉默根系。研究发现,过表达根系中CYP716A53v2基因的表达以及原人参三醇类皂苷含量高于正常根系,沉默根系中CYP716A53v2基因的表达以及原人参三醇类皂苷含量低于正常根系,说明CYP716A53v2基因的功能是转化原人参二醇为原人参三醇。
[参考文献]
[1] Milen I, Georgiev A I, Pavlov T B. Hairy root type plant in vitro systems as sources of bioactive substances[J]. Appl Microbiol Biot,2007,74(6): 1175.
[2] Paek K Y, Murthy H N, Zhong J J. Production of biomass and bioactive compounds using bioreactor technology[M]. Netherlands:Springer, 2014.
[3] Gao M B,Zhang W,Li X T,et al.Expression profiling of genes involved in taxuyunnanine C biosynthesis in cell suspension cultures of Taxus chinensis by repeated elicitation with a newly synthesize jasmonate, in situ absorption and sucrose feeding[J].Chin J Biotechnol,2011,27(1):101.
[4] 趙寿经,侯春喜,徐立新,等. 抑制齐墩果烷型人参皂苷合成支路对达玛烷型人参皂苷生产能力的影响[J]. 吉林大学学报:工学版,2011,41(3):865.
[5] Balusamy S R D,Kim Y J,Rahimi S,et al.Transcript pattern of cytochrome P450,antioxidant and ginsenoside biosynthetic pathway genes under heavy metal stress in Panax ginseng Meyer[J].B Environ Contam Tox,2013,90(2):194.
[6] Moses T,Pollier J,Almagro L,et al.Combinatorial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16α hydroxylase from Bupleurum falcatum[J].Proc Natl Acad Sci USA,2014,111(4):1634.
[7] Zhang H C,Liu J M,Chen H M,et al.Up-regulation of licochalcone A biosynthesis and secretion by Tween 80 in hairy root cultures of Glycyrrhiza uralensis Fisch[J].Mol Biotechnol,2011,47(1):50.
[8] Kim J A,Kim Y S,Choi Y E.Triterpenoid production and phenotypic changes in hairy roots of Codonopsis lanceolata and the plants regenerated from them[J].Plant Biotechnol Rep,2011,5(3):255.
[9] Torkamani M R D,Jafari M,Abbaspour N,et al.Enhanced production of valerenic acid in hairy root culture of Valeriana officinalis by elicitation[J].Open Life Sci,2014,9(9):853.
[10] Sun J,Xu J S,Zhao L Z,et al.Induction of hairy roots and plantlet regeneration of Bupleurum chinense DC[J].Acta Pharmaceutica Sinica,2013,48(9):1491.
[11] Zhu C S,Miao G P,Guo J,et al.Establishment of Tripterygium wilfordii Hook. f. hairy root culture and optimization of its culture conditions for the production of triptolide and wilforine[J].J Microbiol Biotechnol,2014,24:823.
[12] Zhao J L,Zhou L G,Wu J Y.Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures[J].Appl Microbiol Biot,2010,87(1):137.
[13] Guo J,Zhou Y J,Hillwig M L,et al.CYP76AH1 catalyzes turnover of miltiradiene in tanshinones biosynthesis and enables heterologous production of ferruginol in yeasts[J].Proc Natl Acad Sci USA,2013,110(29):12108.
[14] Darvishi E,Kahrizi D,Bahraminejad S,et al.In vitro induction of α-pinene, pulegone, menthol, menthone and limonene in cell suspension culture of pennyroyal (Mentha pulegium)[J].Cell Mol Biol,2015,62(3):7.
[15] Hao G P,Du X H,Zhao F X,et al.Fungal endophytes-induced abscisic acid is required for flavonoid accumulation in suspension cells of Ginkgo biloba[J].Biotechnol Lett,2010,32(2):305.
[16] Qiao X L,Jiang S G,Lv X G,et al. Effects of phytohormones on plant regeneration and production of flavonoids in transgenic Saussurea involucrata hairy roots[J]. Biotechnology,2011,27(1):69.
[17] Li J,Wang J,Li J X,et al. Aspergillus niger enhance bioactive compounds biosynthesis as well as expression of functional genes in adventitious roots of Glycyrrhiza uralensis Fisch[J]. Appl Biochem Biotech,2016,178(3):576.
[18] Simic S G,Tusevski O,Maury S,et al. Fungal elicitor-mediated enhancement in phenylpropanoid and naphtodianthrone contents of Hypericum perforatum L. cell cultures[J]. Plant Cell Tiss Org,2015,122(1):213.
[19] Chen J R,Chen Y B,Ziemiańska M,et al. Co-expression of MtDREB1C and RcXET enhances stress tolerance of transgenic China rose (Rosa chinensis Jacq.)[J]. J Plant Growth Regul,2016,35(2):586.
[20] Park Y J,Thwe A A,Li X,et al. Triterpene and flavonoid biosynthesis and metabolic profiling of hairy roots, adventitious roots, and seedling roots of Astragalus membranaceus[J]. J Agr Food Chem,2015,63(40):8862.
[21] Wang Y H,He Z S,Sun Y X,et al. Study on the production of alkaloid by cell mass suspension culture of Fritillaria cirrhosa[J]. Chin Med Mat,2011,34(2):183.
[22] Van Moerkercke A,Steensma P,Schweizer F,et al. The bHLH transcription factor BIS1 controls the iridoid branch of the monoterpenoid indole alkaloid pathway in Catharanthus roseus[J]. Proc Natl Acad Sci USA,2015,112(26):8130.
[23] Qi X J,Chen R Y,Wang W. Cell suspension culture of Gentiana macrophylla (I)[J].Chin Tradit Herbal Drugs,2010,41(3):472.
[24] Zhao J L,Zhou L G,Wu J Y. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures[J].Appl Microbiol Biot,2010,87(1):137.
[25] Gandi S,Rao K,Chodisetti B,et al.Elicitation of andrographolide in the suspension cultures of Andrographis paniculata[J].Appl Biochem Biotech,2012,168(7):1729.
[26] 曹然,張明生,刘诗雅,等. 不同理化因子对三分三毛状根生长及其莨菪碱含量的影响[J]. 农业生物技术学报,2014,22(2):195.
[27] Sun J,Xu J S,Zhao L Z,et al.Induction of hairy roots and plantlet regeneration of Bupleurum chinense DC[J].Acta pharmaceutica Sinica,2013,48(9):1491.
[28] Verma P,Khan S A,Mathur A K,et al.Fungal endophytes enhanced the growth and production kinetics of Vinca minor hairy roots and cell suspensions grown in bioreactor[J]. Plant Cell Tiss Org,2014,118(2):257.
[29] Zhu C,Miao G,Guo J,et al. Establishment of Tripterygium wilfordii Hook. f. hairy root culture and optimization of its culture conditions for the production of triptolide and wilforine[J]. J Microbiol Biotechnol,2014,24:823.
[30] Lee Y S,Ju H K,Kim Y J,et al. Enhancement of anti-inflammatory activity of Aloe vera adventitious root extracts through the alteration of primary and secondary metabolites via salicylic acid elicitation[J]. PLoS ONE,2013,8(12):e82479.
[31] Kikowska M,Budzianowski J,Krawczyk A,et al. Accumulation of rosmarinic, chlorogenic and caffeic acids in in vitro cultures of Eryngium planum L[J]. Acta Physiol Plant,2012,34(6):2425.
[32] Sheng D F,Chen L. Effects of PEG-6000 stress on tanshinones accumulation in hairy roots of Salvia miltiorrhiza[J]. Chin Tradit Herbal Drugs,2013,44(9):1181.
[33] Wu HY,Baque M A,Lee E J,et al. Scale-up of adventitious root cultures of Echinacea angustifolia in a pilot-scale bioreactor for the production of biomass and caffeic acid derivatives[J].Plant Biotechnol Rep,2013,7(3):297.
[34] Sykowska-Baranek K,Pietrosiuk A,Naliwajski M R,et al.Effect of L-phenylalanine on PAL activity and production of naphthoquinone pigments in suspension cultures of Arnebia euchroma (Royle) Johnst[J].In Vitro Cell Dev-Pl,2012,48(5):555.
[35] Zheng C J,Wu X Y,Wang Z J. Optimization of inducement conditions for hairy roots of Morinda officinalis[J]. Agr Sci Technol,2014,10(1):77.
[36] Lee Y S,Ju H K,Kim Y J,et al.Enhancement of anti-inflammatory activity of Aloe vera adventitious root extracts through the alteration of primary and secondary metabolites via salicylic acid elicitation[J].PLoS ONE,2013,8(12):e82479.
[37] Silja P K,Satheeshkumar K. Establishment of adventitious root cultures from leaf explants of Plumbago rosea and enhanced plumbagin production through elicitation[J].Ind Crop Prod,2015,76(1):479.
[38] Wang Q J,Zheng L P,Sima Y H,et al. Methyl jasmonate stimulates 20-hydroxyecdysone production in cell suspension cultures of Achyranthes bidentata[J].Plant Omics,2013,6(2):116.
[39] Suthar S,Ramawat K G. Growth retardants stimulate guggulsterone production in the presence of fungal elicitor in fed-batch cultures of Commiphora wightii[J].Plant Biotechnol Rep,2010,4(1):9.
[40] Chavan J J, Gaikwad N B, Umdale S D, et al. Efficiency of direct and indirect shoot organogenesis, molecular profiling, secondary metabolite production and antioxidant activity of micropropagated Ceropegia santapaui[J]. Plant Growth Regul,2014,72(1): 1.
[41] Liu W, Li P J, Qi X M, et al. DNA changes in barley (Hordeum vulgare) seedlings induced by cadmium pollution using RAPD analysis[J]. Chemosphere, 2005,61(1): 158.
[42] Rong Z, Yin H. A method for genotoxicity detection using random amplified polymorphism DNA with Danio rerio[J].Ecotox Environ Safe,2004, 58(1): 96.
[43] Punja Z K,Feeney M, Schluter C,et al.Multiplication and germination of somatic embryos of American ginseng derived fromsuspension cultures and biochemical and molecular analyses of plantlets[J]. In Vitro Cell Dev-Pl, 2004,40: 329
[44] Sedov K A, Fomenkov A A, Solov′yova, et al. The level of genetic variability of cells in prolonged suspension culture of Arabidopsis thaliana[J]. Biol Bull, 2014,41:493.
[45] 黄璐琦, 郭兰萍, 胡娟, 等. 道地药材形成的分子机制及其遗传基础[J]. 中国中药杂志,2008,33(20):2303.
[46] 黄璐琦,刘昌孝. 分子生药学[M].北京:科学出版社,2015.
[47] 黄新,邱德有,黄璐琦.茉莉酸甲酯对中国红豆杉细胞基因表达的mRNA差异显示研究[J].分子植物育种,2006,4(5):627.
[48] 李娟,盛维瑾,刘大为,等.锌离子影响丹参金属硫蛋白MT2基因表达[J].分子植物育种,2007,5(3):389.
[49] 刘峻,丁家宜,周倩耘,等.真菌诱导子对人参毛状根皂苷生物合成和生长的影响[J].中国中药杂志,2004,29(4):302.
[50] Zhao J, Davis L C, Verpoorte R. Elicitor signal transduction leading to production of plant secondary metabolites[J]. Biotechnol Adv, 2005, 23: 283.
[51] 許道琦. 真菌诱导白桦三萜积累的营养生理和分子机制的初步研究[D]. 长春:东北林业大学,2013.
[52] Zhao Q, Hu Y Q, Guo W H, et al. Elicitor-induced indole alkaloid biosynthesis in Catharanthus roseus cell cultures is related to Ca2+-influx and the oxidative burst[J]. Plant Sci, 2001, 161: 423.
[53] 崔晋龙,付少彬,高芬,等. 真菌诱导植物次生代谢产物积累的信号机制及在药用植物中的应用[J].中草药,2012,43(8): 1647.
[54] 周敏. 内生真菌及其诱导子与长春花悬浮细胞生物碱合成代谢相关性研究[D]. 长沙:湖南农业大学, 2005.
[55] Li T T, Hu Y Y, Du X H, et al. Salicylic acid alleviates the adverse effects of salt stress in Torreya grandis cv. Merrillii seedlings by activating photosynthesis and enhancing antioxidant systems[J]. PLoS ONE, 2014, 9 (10):e109492.
[56] 张莲莲,谈锋.真菌诱导子在药用植物细胞培养中的作用机制和应用进展[J].中草药,2006,37(9):1426.
[57] Xu M J, Dong J F, Zhu M Y, et al. Nitric oxide mediates the fungal elicitor-induced hypericin production of Hypericum perforatum cell suspension cultures through a jasmonic-acid-dependent signal pathway[J]. Plant Physiol, 2005, 139: 991.
[58] Dong J, Wan G W, Liang Z S. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell cultures[J]. J Biotechnol, 2010, 148: 99.
[59] 黄超.小分子化合物的添加提高人参皂苷产量及其作用机制探索[D].上海:华东理工大学,2013.
[60] Bulgakov V P, Tchernoded G K, Mischenko N P, et al. Effect of salicylic acid, methyl jasmonate, ethephon and cantharidin on anthraquinone production by Rubia cordifolia callus cultures transformed with the rolB and rolC genes[J]. J Biotechnol, 2002, 97: 213.
[61] Aziz A, Poinssot B, Daire X, et al. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola[J].Mol Plant Microbe In, 2003, 16: 1118.
[62] Hu X, Neill S J, Cai W, et al. Nitric oxide mediates elicitor-induced saponin synthesis in cell cultures of Panax ginseng[J].Funct Plant Biol, 2003, 30: 901.
[63] Linden J C, Phisalaphong M. Oligosaccharides potentiate methyl jasmonate-induced production of paclitaxel in Taxus canadensis[J].Plant Sci, 2000, 158: 41.
[64] Zhang C H, Yan Q, Cheuk W K, et al. Enhancement of tanshinone production in Salvia miltiorrhiza hairy root culture by Ag+ elicitation and nutrient feeding[J]. Planta Med, 2004, 70: 147.
[65] 邢丙聰. 丹参毛状根中酚酸类和酮类成分合成途径中bHLH和WD40转录因子的调控[D]. 杭州:浙江理工大学,2015.
[66] Fujita M, Fujita Y, Noutoshi Y, et al. Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks[J]. Curr Opin Plant Biol, 2006, 9: 436.
[67] Vander F L, Zhang H, Menke F L H, et al. A Catharanthus roseus BPF-1 homologue interacts with an elicitor-responsive region of the secondary metabolite biosynthetic gene Str and is induced by elicitor via a JA-independent signal transduction pathway[J].Plant Mol Biol, 2000, 44:675.
[68] 王劲波. 中生菌素对水稻悬浮细胞主要防卫反应基因转录表达的影响[D].北京:中国农业科学院,2002.
[69] Landi L, Feliziani E, Romanazzi G. Expression of defense genes in strawberry fruits treated with different resistance inducers[J]. J Agr Food Chem, 2014, 62: 3047.
[70] 王和勇,罗恒,孙敏. 诱导子在药用植物细胞培养中的应用[J].中草药,2004,35(8):1.
[71] Cruickshank I A M, Perrin D R. The isolation and partial characterization of monilicolin A, a polypeptide with phaseollin-inducing activity from Monilinia fructicola[J]. Life Sci, 1968, 7(1):449.
[72] Bahabadi S E, Sharifi M, Chashmi N A, et al. Significant enhancement of lignan accumulation in hairy root cultures of Linum album using biotic elicitors[J]. Acta Physiol Plant, 2014, 36 (1):3325.
[73] Saikat D, Moumita G, Urmi D, et al. Signal transducer and oxidative stress mediated modulation of phenylpropanoid pathway to enhance rosmarinic acid biosynthesisin fungi elicited whole plant culture of Solenostemon scutellarioides[J]. Enzyme Microb Tech, 2014, 66(1): 1.
[74] Zhang S C, Yan Y, Wang B Q, et al. Selective responses of enzymes in the two parallel pathways of rosmarinic acid biosynthetic pathway to elicitors in Salvia miltiorrhiza hairy root cultures[J]. J Biosci Bioeng,2014, 117 (5): 645.
[75] Sudhamoy M, Adinpunya M. Accumulation of cell wall-bound phenolic metabolites and their upliftment in hairy root cultures of tomato (Lycopersicon esculentum Mill.)[J]. Biotechnol Lett, 2008, 30 (1):1253.
[76] Wang Y, Dai C C, Cao J L, et al. Comparison of the effects of fungal endophyte Gilmaniella sp. and its elicitor on Atractylodes lancea plantlets[J]. World J Microb Biot, 2012, 28 (1):575.
[77] Hao G P, Du X H, Zhao F, et al. Fungal endophytes-induced abscisic acid is required for flavonoid accumulation in suspension cells of Ginkgo biloba[J]. Biotechnol Lett, 2010, 32 (1):305.
[78] Wang J W, Zhang Z, Tan R X, Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp.[J].Biotechnol Lett, 2001, 23 (1): 857.
[79] Fan G Z, Zhai Q L, Li X C, et al. Compounds of Betula platyphylla cell suspension cultures in response to fungal elicitor[J]. Biotechnol Biotec Eq, 2013, 27 (1):3569.
[80] Ravi P G, Arvind C, Tukaram D N. Influence of biotic and abiotic elicitors on four major isomers of boswellic acid in callus culture of Boswellia serrata Roxb[J].Plant Omics, 2011, 4(4):169.
[81] Gao F K, Yong Y H, Dai C C. Effects of endophytic fungal elicitor on two kinds of terpenoids production and physiological indexes in Euphorbia pekinensis suspension cells[J].J Med Plants Res, 2011, 5 (18): 4418.
[82] Zhao J, Zhong L, Zou L, et al. Efficient promotion of the sprout growth and rutin production of tartary buckwheat by associated fungal endophytes[J].Cereal Res Commun, 2014, 42(3): 401.
[83] Ming Q L, Su C Y, Zheng C J, et al. Elicitors from the endophytic fungus Trichoderma atroviride promote Salvia miltiorrhiza hairy root growth and tanshinone biosynthesis[J].J Exp Bot, 2013, 64 (18): 5687.
[84] Zhao J L, Zheng B B, Li Y, et al. Enhancement of diepoxin ζ production by yeast extract and its fractions in liquid culture of Berkleasmium-like endophytic fungus Dzf12 from Dioscorea zingiberensis[J]. Molecules, 2011, 16(1): 847.
[85] Pereira P S, Fábio K T, Suzelei D C F, et al. Enhanced triterpene production in Tabernaemontana catharinensis cell suspension cultures in response to biotic elicitors[J].Quim Nova,2007, 30 (8):1849.
[86] Awad V, Kuvalekar A, Harsulkar A. Microbial elicitation in root cultures of Taverniera cuneifolia (Roth) Arn, for elevated glycyrrhizic acid production[J]. Ind Crop Prod, 2014, 54: 13.
[87] Li P Q, Mou Y, Shan T J, et al. Effects of polysaccharide elicitors from Endophytic Fusarium oxysporium Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis[J].Molecules,2011, 16(1): 9003.
[88] Li P Q, Mao Z L, Lou J F, et al. Enhancement of diosgenin production in Dioscorea zingiberensis cell cultures by oligosaccharides from its endophytic fungus Fusarium oxysporum Dzf17[J]. Molecules, 2011, 16(1), 10631.
[89] Li P Q, Luo H Y, Meng J J, et al. Effects of oligosaccharides from endophytic Fusarium oxysporum Dzf17 on activities of defense-related enzymes in Dioscorea zingiberensis suspension cell and seedling cultures[J]. Electronic J Biotechnol, 2014, 17 (1): 156.
[90] Mou Y, Zhou K Y, Xu D, et al. Enhancement of diosgenin production in plantlet and cell cultures of Dioscorea zingiberensis by palmarumycin C13 from the endophytic fungus, Berkleasmium sp. Dzf12[J]. Trop J Pharm Res, 2015, 14 (2): 241.
[91] Li Y C,Tao W Y. Effects of paclitaxel-producing fungal endophytes on growth and paclitaxel formation of Taxus cuspidata cells[J]. Plant Growth Regul, 2009, 58 (1): 97.
[92] MaY N, Han C, Chen J Y, et al. Fungal cellulase is an elicitor but its enzymatic activity is not required for its elicitor activity[J]. Mol Plant Pathol, 2015, 16(1): 14.
[93] Verma P, Khan S A, Mathur A K, et al. Fungal endophytes enhanced the growth and production kinetics of Vinca minor hairy roots and cell suspensions grown in bioreactor[J].Plant Cell Tiss Org, 2014, 118 (1): 257.
[94] Li Y C, Tao W Y. Paclitaxel-producing fungal endophyte stimulates the accumulation of taxoids in suspension cultures of Taxus cuspidate[J].Sci Hortic-amsterdam, 2009, 121 (1): 97.
[95] Chodisetti B, Rao K, Gandi S, et al. Improved gymnemic acid production in the suspension cultures of Gymnema sylvestre through biotic elicitation[J]. Plant Biotechnol Rep, 2013, 7(1):519.
[96] Verma P, Khan S A, Mathur A K, et al. Improved sanguinarine production via biotic and abiotic elicitations and precursor feeding in cell suspensions of latex-less variety of Papaver somniferum with their gene expression studies and upscaling in bioreactor[J]. Protoplasma, 2014, 251 (3):1359.
[97] Ali M, Abbasi B H, AliG S. Elicitation of antioxidant secondary metabolites with jasmonates and gibberellic acid in cell suspension cultures of Artemisia absinthium L.[J].Plant Cell Tiss Org, 2015, 120 (4):1099.
[98] Wang J,Gao W Y, Zuo B M, et al. Effect of methyl jasmonate on the ginsenoside contentof Panax ginseng adventitious root cultures and on the genes involved in triterpene biosynthesis[J]. Res Chem Intermediat, 2013, 39 (8):1973.
[99] Wang W, Zhao Z J, Xu Y F, et al. Efficient elicitation of ginsenoside biosynthesis in cell cultures of Panax notoginseng by using self-chemically-synthesized jasmonates[J].Biotechnol Bioproc E, 2005, 10 (5):162.
[100] Chen S L, Zhu X X, Li C F, et al. Genomics and synthetic biology of traditional Chinese medicine[J]. Acta Pharm Sin, 2012, 47: 1070.
[101] Subramaniyam S, Mathiyalagan R, Natarajan S, et al.Transcript expression profiling for adventitious roots of Panax ginseng Meyer[J]. Gene, 2014, 546:89.
[102] Nuruzzaman M, Cao H Z, Xiu H, et al. Transcriptomics-based identification of WRKY genes and characterization of a salt and hormone-responsive PgWRKY1 gene in Panax ginseng[J].Acta Biochim Biophys Sin, 2015, 22:1.
[103] Jayakodi M, LeeS C, Park H S, et al. Transcriptome profiling and comparative analysis of Panax ginseng adventitious roots[J].J Ginseng Res, 2014, 38: 278.
[104] Park S B, Chun J H, Ban Y W, et al. Alteration of Panax ginseng saponin composition by overexpression and RNA interference of the protopanaxadiol 6-hydroxylase gene (CYP716A53v2)[J].J Ginseng Res, 2016,40:47.
[105] Moses T, Pollier J, Almagro L, et al. Combinatorial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16α hydroxylase from Bupleurum falcatum[J].Proc Natl Acad Sci USA, 2014, 111 (4):1634.
[106] Huang Z W, Lin J C, Cheng Z X, et al. Production of oleanane-type sapogenin in transgenic rice via expression of β-amyrin synthase gene from Panax japonicus C. A. Mey[J].BMC Biotechnol, 2015, 15 (17):45.
[107] Zhao S C, Park C H, Li X H, et al. Accumulation of rutin and betulinic acid and expression of phenylpropanoid and triterpenoid biosynthetic genes in mulberry (Morus alba L.)[J].J Agric Food Chem, 2015, 63 (23): 8622.
[108] Biazzi E, Carelli M, Tava A, et al. CYP72A67 catalyzes a key oxidative step in Medicago truncatula hemolytic saponin biosynthesis[J]. Mol Plant, 2015, 8 (3): 1493.
[109] Dai L H, Liu C, Zhu Y M, et al. Functional characterization of cucurbitadienol synthase and triterpene glycosyltransferase involved in biosynthesis of mogrosides from Siraitia grosvenorii[J].Plant Cell Physiol, 2015, 56(6):1172.
[110] Huang Z W, Lin J C, Cheng Z X, et al. Production of dammarane-type sapogenins in rice by expressing thedammarenediol-Ⅱ synthase gene from Panax ginseng C.A. Mey[J]. Plant Sci, 2015, 239 (3):106.
[111] Zhao F L, Bai P, Liu T, et al. Optimization of a cytochrome P450 oxidation system for enhancing protopanaxadiol production in Saccharomyces cerevisiae[J].Biotechnol Bioeng, 2016, 9999: 1.
[112] Martin V J J, Pitera D J, Withers S T, et al. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids[J].Nat Biotechnol,2003, 21 (7): 796.
[113] Ignea C, Athanasakoglou A, Ioannou E et al. Carnosic acid biosynthesis elucidated by a synthetic biology platform[J]. Proc Natl Acad Sci USA, 2015, doi/10.1073/pnas.1523787113.
[責任编辑 吕冬梅]
