尿素对中肋骨条藻与米氏凯伦藻生长的影响
2017-06-22李志林周艳蕾王雪景石晓勇张传松
李志林,周艳蕾,王雪景, 石晓勇,3, 张传松,*
1 中国海洋大学化学化工学院,青岛 266100 2 中国海洋大学海洋化学理论与工程技术教育部重点实验室,青岛 266100 3 国家海洋局海洋减灾中心, 北京 100194
尿素对中肋骨条藻与米氏凯伦藻生长的影响
李志林1,2,周艳蕾1,2,王雪景1,2, 石晓勇1,2,3, 张传松1,2,*
1 中国海洋大学化学化工学院,青岛 266100 2 中国海洋大学海洋化学理论与工程技术教育部重点实验室,青岛 266100 3 国家海洋局海洋减灾中心, 北京 100194
采用实验室一次性培养,研究了尿素对我国东海赤潮优势藻中肋骨条藻(Skeletonemacostatum)和米氏凯伦藻(Kareniamikimotoi)生长的影响。结果表明,中肋骨条藻和米氏凯伦藻均能在不同比例尿素的条件下较好地生长。随着培养液中尿素比例的增大,中肋骨条藻细胞生长速率(0.91—0.82/d)逐渐减小,平台期最大生物量(2.0×105—1.2×105个/mL)也逐渐减小,而米氏凯伦藻细胞的生长速率(0.36—0.51/d)逐渐增大,最大生物量基本不变(约1.1×104个/mL)。在平台期中肋骨条藻培养液中氮盐浓度最低下降到2.5μmol/L左右维持不变,而米氏凯伦藻氮盐浓度最低下降到1.0μmol/L左右。在指数生长期,随着细胞的生长溶解有机氮(DON, Dissolved Organic Nitrogen)含量迅速增加,中肋骨条藻介质中DON的浓度达到最大值(5—6μmol/L),然后浓度基本不变。米氏凯伦藻介质中DON在指数生长阶段达到最大值(2—3μmol/L)后开始下降。中肋骨条藻单细胞颗粒氮的含量(约为10-6μmol,平台期约为10-7μmol)要远远小于米氏凯伦藻(指数期约为10-4μmol,平台期约为10-6μmol)。研究表明,两种藻对尿素的吸收利用存在明显差异,在较低的溶解无机氮和较高的溶解有机氮环境中,甲藻有更好的适应性,该研究对于解释我国长江口春季硅藻和甲藻赤潮的演替有借鉴的意义。
氮;尿素;赤潮;中肋骨条藻;米氏凯伦藻
赤潮是一种有害的海洋生态异常现象,赤潮的发生不仅会为近海水产养殖业带来巨大的经济损失,破坏海洋生态环境[1-2],还会危害人类的身体健康。赤潮的发生与很多因素有关,其中包括富营养化、水环境动力、生物环境、气相气候等多种因素,其中营养盐是其发生的必要物质基础[3- 5]。海水中溶解态氮营养盐主要包括溶解态无机氮(DIN)、溶解态有机氮(DON)。以往对引发赤潮的氮营养盐的研究主要集中在DIN,对DON的研究相对很少。直到30多年前Antia等人提出DON在海洋初级生产力中起着不容忽视的作用,才引起海洋界的高度重视。
实验研究发现,DON不仅可以做某些浮游生物种属的氮源,而且在浮游生物优势种的选择上也起着一定的作用[6]。DON含量的增加还会诱发有毒赤潮暴发频率的上升[7]。调查表明,在近岸海水中,DON在总氮中占有相当可观的含量,在胶州湾DON占总氮的66%[8],在珠江口外海DON超过总氮的80%[9],在美国的切萨皮克湾,DON在夏季占总氮含量的90%以上[10]。
尿素是DON的重要组成部分。在东海近岸海域尿素含量为(1.33±0.84)μmol/L,占DON的5.63%—15.73%[11],在黄渤海海域尿素含量为(0.84±0.20)μmol/L,占DON的6.80%—9.50%[12],杭州湾尿素含量为0.19—6.55μmol/L[13],大亚湾尿素含量为1.98μmol/L[14]。
近年来,越来越多的研究表明,尿素是某些浮游生物吸收的主要氮源。一些浮游生物还会优先吸收尿素,定鞭藻会优先利用DON中的尿素,许多有害的藻,如赤潮异湾藻、球形棕囊藻,角毛藻在培养中也更喜欢尿素[15]。陈园等人的研究表明,在硝态氮和尿素组成的混合氮源中当尿素比例大于50%时,球形棕囊藻的细胞密度要大于以硝态氮为单一氮源的实验组[16]。随着全球人口持续增长、工农业的发展以及尿素等氮肥的大量使用,越来越多的生活污水和农业灌溉水随着排污口或地表径流输入近岸水体,使得海水中尿素的浓度越来越高[17]。在经常使用尿素肥料的墨西哥Yaqui海湾,随着灌溉高峰期的来临,高于40%的氮流失于表层径流,施肥前后海湾中尿素浓度从13μmol/L增加到43μmol/L[18]。在美国的Chesapeake海湾,通过连续5a的监测发现在春末用尿素肥料或家禽肥料对小麦和玉米施肥时,尿素浓度经常超过10μmol/L,其浓度比该海湾表层海水平均浓度高50倍以上[10]。近年来,随着我国长江口及邻近海域中尿素等有机氮含量逐渐增加,该海域每年春季均会暴发以东海原甲藻为优势藻种的赤潮,且其暴发规模及甲藻的优势度呈逐年增加的趋势[19]。同时,该海域赤潮爆发在其生消过程中优势种呈现出“硅藻→甲藻→硅藻”的演替态势。
为了研究DON在赤潮发生发展及演替过程中的作用,本文选取东海赤潮优势种中肋骨条藻(硅藻)与米氏凯伦藻(甲藻),设计不同比例的尿素与硝氮做混合氮源,以期为揭示有机氮比例对浮游生物生长的影响提供科学基础,并为实际预防和治理赤潮的氮管理提供依据。
1 材料与方法
1.1 实验材料
实验藻种选用的中肋骨条藻(Skeletonemacostatum)与米氏凯伦藻(Kareniamikimotoi),均取自中国海洋大学海洋污染生态化学重点实验室。藻种在f/2培养液中培养,其中只有N、P浓度按实验要求添加,培养至指数期,然后重新接种到新的培养基中,依次循环培养3代,然后饥饿培养2d,接种,开始实验。
1.2 培养条件
实验藻种于室内光照培养箱中用1L的锥形瓶培养。培养温度(22±1)℃,光照强度为3500 lux,光暗比12H:12H。培养所用海水为人工海水,盐度30.5[20],人工调节pH为8.2。初始藻种接种密度约为104个/L。
1.3 实验设计
实验采用一次性培养。每个藻种设计5个实验处理组A—E,每个实验处理组做两组平行,总氮添加量为32μmol/L,磷(KH2PO4)为2μmol/L。每个实验组氮的添加状况如表1。
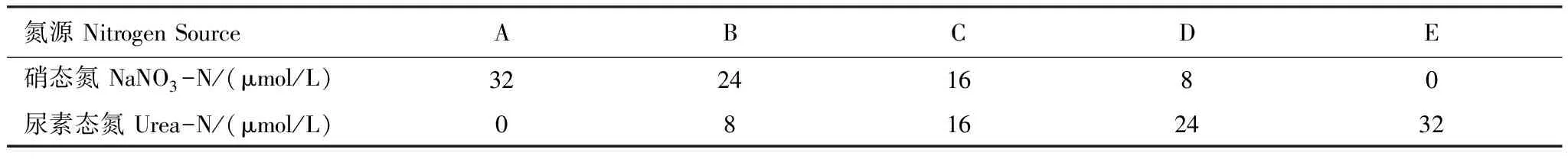
表1 5个实验处理组氮盐比例
1.4 实验测定指标
1.4.1 细胞计数
从接种第2天起,使用0.1mL的浮游生物计数框,在Olympus显微镜下进行细胞计数。每个样品取样3次,取其平均值。
1.4.2 比生长速率
比生长速率[21]根据下面公式拟合得到:
μ=(lnN2-lnN1)/(t2-t1)
式中,N2、N1分别为t2、t1时刻藻细胞的密度。
1.4.3 样品的测定
尿素的测定采用二乙酰一肟-盐酸氨基脲法[22],溶解无机氮的测定依据海洋调查规范之海水化学要素调查(GBT12763.4- 2007)分光光度法测定,其中氨氮的测定采用次溴酸钠氧化法,检出限为0.03μmol/L,硝酸盐的测定使用镉铜还原法,检出限为0.04μmol/L,亚硝酸盐的测定使用重氮偶氮比色法,检出限为0.02μmol/L。总溶解氮(DTN)采用碱性过硫酸钾消化,TechniconAAⅢ营养盐自动分析仪测定,测量精度为
2 实验结果
2.1 不同比例尿素对中肋骨条藻与米氏凯伦藻生长的影响
图1为中肋骨条藻和米氏凯伦藻在不同尿素比例下的生长曲线,表2为中肋骨条藻和米氏凯伦藻在指数生长期的比生长速率。
从中肋骨条藻生长曲线的图中可以看出,尿素是中肋骨条藻生长繁殖所需要的一种氮源,中肋骨条藻在以尿素为氮源的介质中也能较好的生长繁殖,但是其平台期最大生物量呈现随介质环境中尿素比例的增大而减小的趋势。具体来说,不同处理组在开始的前3天处于迟滞期,细胞密度没有明显的变化,第4天开始同时进入指数生长期,经过3—4d的指数生长,细胞密度进入相对稳定的平台期,各处理组平台期中肋骨条藻随尿素含量的增加其细胞密度呈逐渐降低的趋势。其中A处理组(100%硝氮)在平台期的细胞密度约为2.0×105个/mL,而E处理组(100%尿素)在平台期的细胞密度仅为1.2×105个/mL。同时,由表2可知,A组即硝氮含量100%时的比生长速率最大,为0.91/d,而E组即尿素含量100%时比生长速率最小,为0.82/d,中肋骨条藻的生长速率有随营养环境中尿素比例的增大而减小的趋势。
米氏凯伦藻的生长曲线与中肋骨条藻生长曲线显著不同,米氏凯伦藻在以硝酸盐为氮源和以尿素为氮源时其生长曲线没有明显的差异,平台期最大生物量随介质环境中尿素比例的增大基本不变。具体来说,和中肋骨条藻相比米氏凯伦藻迟滞期明显延长,在实验前7d其细胞密度均没有发生明显的变化,第9天米氏凯伦藻细胞密度开始明显的增多,进入细胞快速生长的指数生长期,大概第15天进入稳定生长的平台期。平台期细胞密度差别不大,均在1.1×104个/mL左右。同时,不同处理组米氏凯伦藻的比生长率在0.36—0.51/d之间,且在整体上呈现随混合氮源中尿素比例的增加其比生长速率不断增大的趋势。
对于中肋骨条藻,随着环境中尿素比例的增大,细胞生长速率逐渐减小,同时平台期的细胞密度也逐渐减小,说明中肋骨条藻在生长过程中更倾向于吸收无机氮,对无机氮的吸收利用要大于对有机氮的吸收利用。而对于米氏凯伦藻,随着环境中尿素比例的增加,细胞的生长速率逐渐增大,说明米氏凯伦藻对尿素有较好的亲和性。同时,平台期的细胞密度基本没有差别,也说明米氏凯伦藻在生长过程中能同时吸收利用有机氮和无机氮。因此,当海洋环境中DON/DIN增大时,以米氏凯伦藻为典型代表的甲藻在与以中肋骨条藻为典型代表的硅藻进行竞争时将处以有利的状况。
Heil研究表明硅藻类浮游植物更喜欢利用硝酸盐[23],而Sofomon研究发现,甲藻类浮游植物比较喜欢吸收利用有机氮中的尿素,海域中尿素等有机氮含量的上升与这种浮游植物的激增有着密切的关系[24]。同样,李京等人在2005年东海赤潮暴发过程中发现米氏凯伦藻对尿素有着较高的吸收速率,更喜欢以尿素为氮源[25]。近年来,随着我国近海海域中尿素等有机氮含量的增加[26],甲藻类赤潮发生的频率及比例明显增加[27- 29],本文的研究结果也能较好的说明这一点。
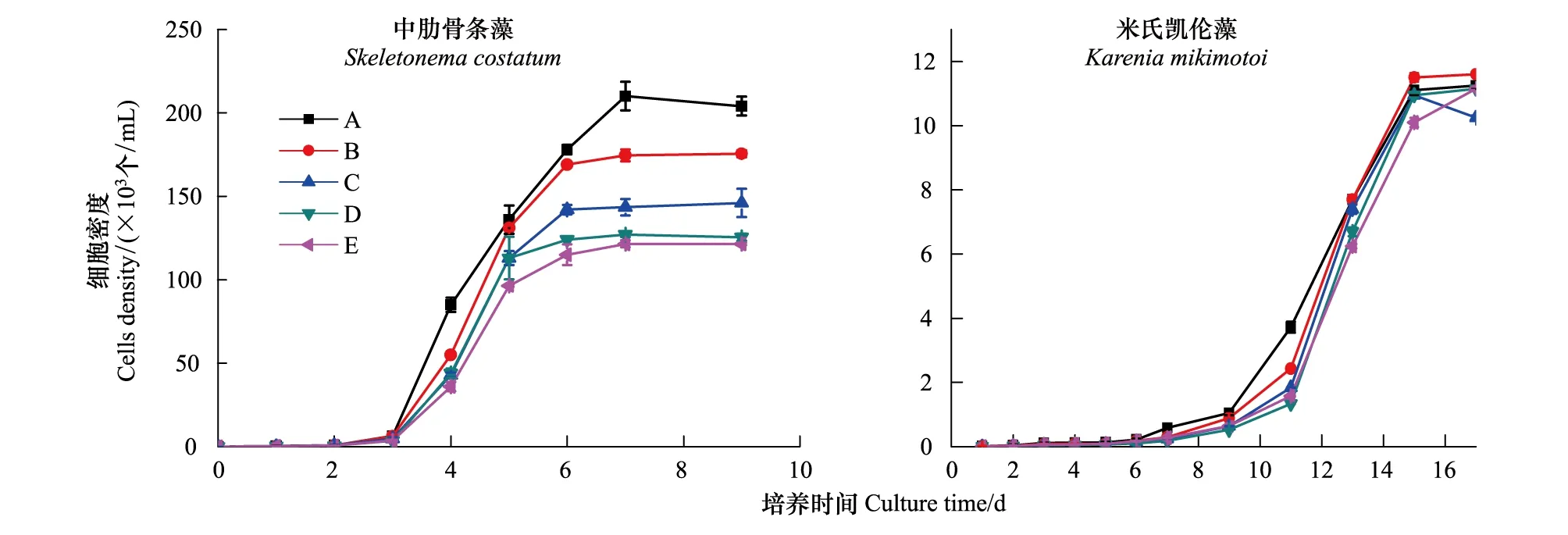
图1 不同比例尿素条件下中肋骨条藻和米氏凯伦藻的生长曲线Fig.1 Growth curves of Skeletonema costatum and Karenia mikimotoi under different ratio of urea
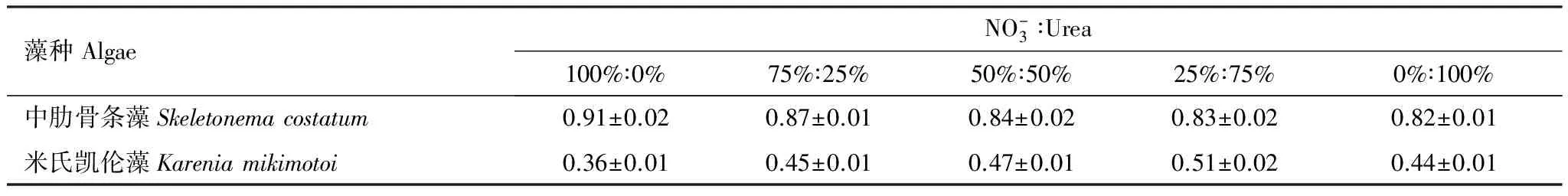
表2 不同尿素比例下两种藻的比生长速率/d-1
2.2 培养介质中氮盐的变化
图2为中肋骨条藻和米氏凯伦藻在培养过程中硝氮和尿素浓度的变化曲线。
从系列营养盐的变化曲线可以发现,在不同生长时期,中肋骨条藻和米氏凯伦藻的营养盐变化存在差异性。对于中肋骨条藻,在培养的前3d,即细胞生长的迟滞期,各个处理组添加的硝氮和尿素的浓度变化均较小,进入指数生长期后,两种氮盐的浓度都迅速降低,在培养的第7天,藻细胞进入生长的平台期,硝氮和尿素的浓度降到最低值,约为2.5μmol/L。对于米氏凯伦藻,在细胞生长的迟滞期(培养的前7d),细胞密度没有明显的增长,但是各个处理组添加的硝氮和尿素的浓度都有大幅度的降低,其中A组(硝氮100%)硝氮浓度从最初的32μmol/L下降到约25μmol/L(约20%),E组(尿素100%)尿素浓度从最初的32μmol/L下降到22.5μmol/L(约30%),进入指数生长期后,两种氮盐的浓度同样迅速下降,到培养的第14天时,细胞生长开始进入平台期,两种氮盐的浓度降到最低值,约为1μmol/L。
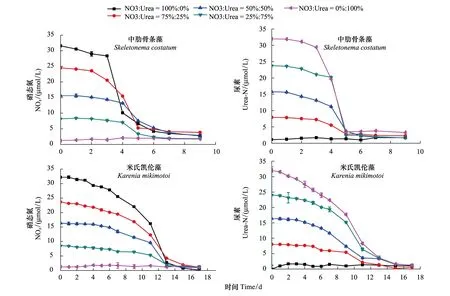
图2 两种藻在培养过程中营养盐的变化Fig.2 The change of the added nutrient of the two kinds of algae in the culture process
比较营养盐的变化可以看出,在藻细胞生长过程中,两种藻对尿素和硝氮吸收存在差异。中肋骨条藻生长曲线的变化特点和介质中营养盐浓度曲线的变化特点正好相反,在迟滞期,中肋骨条藻细胞数目增加缓慢,介质中氮盐的浓度变化不大,此时藻细胞吸收的营养盐也较少。进入指数生长期后,细胞密度急剧增加,介质中营养盐浓度下降迅速。进入平台期后,当介质环境中营养盐浓度较低时(2.5μmol/L),细胞密度也不再增大。由此可知,中肋骨条藻吸收的营养盐主要用于繁殖,即藻细胞数目的增加,当氮盐浓度较低不能满足藻细胞生长分裂的需求时,细胞密度也不再增大。米氏凯伦藻作为一种甲藻,可以高效的吸收介质中的营养盐,吸收的营养盐除了用于繁殖外,还有一部分储存起来[30],当介质中营养盐缺乏时仍然能够继续生长繁殖,在迟滞期,细胞密度增加缓慢,米氏凯伦藻也会吸收环境中的营养盐并在细胞中储存起来,从而导致培养初期氮盐较大幅度的降低。当培养至第14天时,介质中氮盐浓度已经降到最低值(小于1μmol/L),此时藻细胞仍然能利用储存起来的营养盐继续生长分裂,因此细胞密度还会持续增加,这说明米氏凯伦藻更能适应低浓度营养盐环境。
甲藻和硅藻生存策略上的明显差异导致它们不能同时成为赤潮事件中的优势种,而是演替性的出现[30],因此我国近岸海域常常暴发硅藻和甲藻演替性的赤潮,初期在营养盐丰富的条件下,迟滞期短、生长速率较快的中肋骨条藻等硅藻往往成为优势种,并发展为硅藻赤潮。后期,随着营养盐浓度的较低,常常演替为优势种为甲藻的赤潮[31- 32]
2.3 水体中DON的含量变化
图3为两种藻培养过程中溶解有机氮(扣除尿素)含量的变化曲线。从图中可以看出各个实验组中肋骨条藻培养液中DON的变化趋势与其生长曲线基本一致,在细胞生长迟滞期含量很少且基本不变,在细胞生长指数期随着细胞密度的增大水体中DON含量开始增加,在指数期结束时达到最大值(5—6μmol/L)后变化很小。米氏凯伦藻培养液中DON在生长迟滞期含量同样很低,在指数生长期随着藻细胞生长,含量开始迅速增加,在培养的第11天,细胞生长还未进入平台期时,DON的含量已经达到最大值约3.3μmol/L,随后出现下降的趋势。
以上变化表明,藻类在正常的生长过程中,自身的代谢会产生DON并释放到环境水体中,在细胞迟滞期,细胞数量少,生长速率低,代谢活动弱,因此几乎不产生或是产生少量的DON;而随着培养的进行,细胞生长进入指数期,此时细胞生长率高,细胞密度快速增大,代谢活动强,从而产生大量的DON。
中肋骨条藻培养液中DON的含量高于米氏凯伦藻DON的含量,且达到最大值后中肋骨条藻中DON含量出现平台期而米氏凯伦藻DON出现下降趋势,原因在于中肋骨条藻的细胞密度大于米氏凯伦藻的细胞密度,因此在生长过程中代谢产生的DON的含量就会高于米氏凯伦藻代谢产生的DON的含量,同时中肋骨条藻在生长过程中对有机氮的吸收利用能力较低,所以培养后期DON含量较高并在细胞生长的稳定期保持一定含量后变化不大。而米氏凯伦藻对有机氮的利用能力较高,并且会重新吸收自身产生的DON,因此在培养过程中DON含量较低且在达到最大值后会出现下降的趋势[33]。甲藻对有机氮能够有效的吸收和利用,这一特点也有助于在硅藻赤潮发生末期,优势种由硅藻向甲藻的演替。
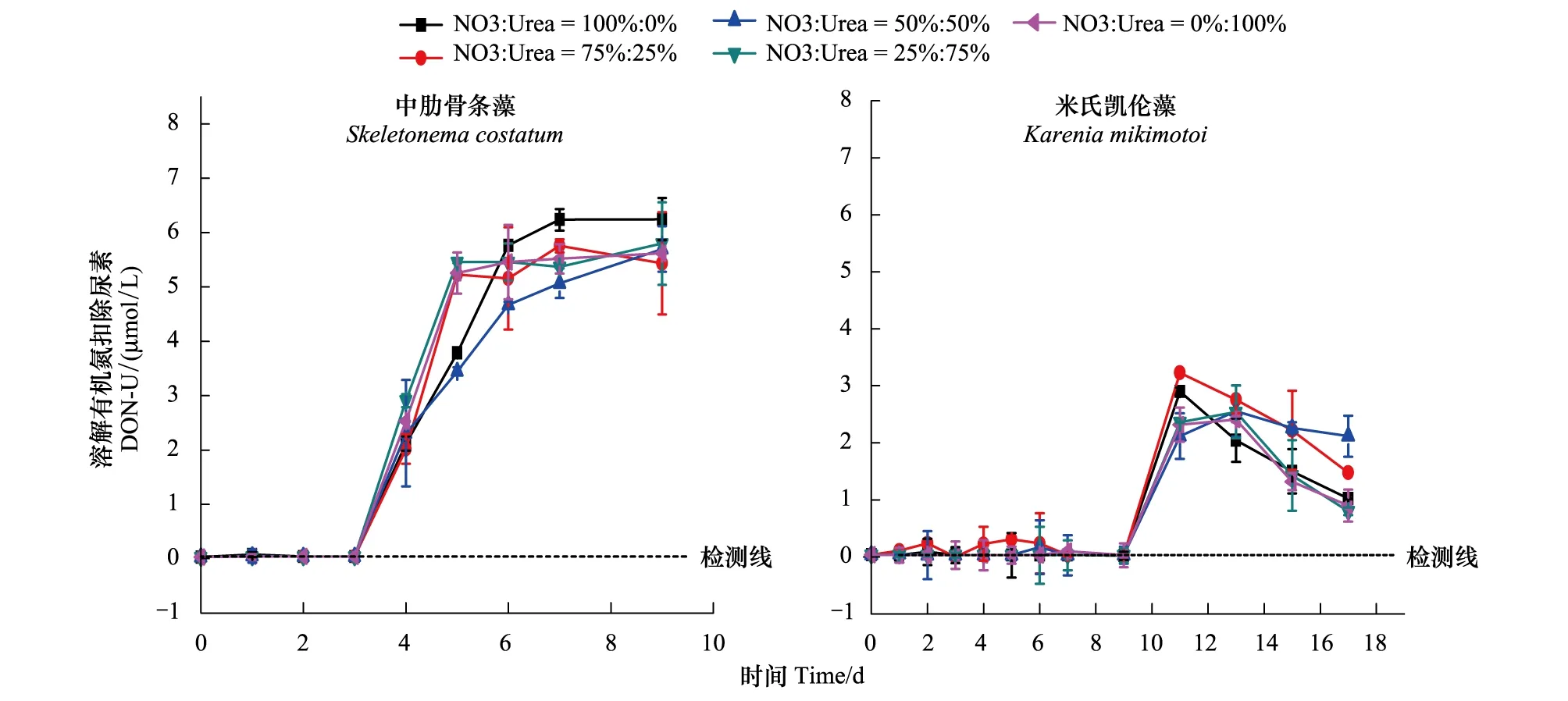
图3 两种藻在培养过程中DON的变化Fig.3 The variation of DON in culture of the algae
2.4 单细胞颗粒态氮的变化
图4为中肋骨条藻和米氏凯伦藻在培养过程中单个细胞颗粒态氮(PN,Particulate Nitrogen)含量的变化曲线。整体上来看,在实验培养的第1天,两种藻单细胞颗粒氮含量均呈现出增加的趋势,其原因是饥饿处理后的藻细胞在接触到丰富的营养盐时会迅速吸收到体内,合成细胞自身生长所必须的组分,如氨基酸、核酸、蛋白质等[3- 5],使得单细胞颗粒氮的含量增加明显。此后,在指数生长期,藻细胞对氮的吸收速率小于其分裂速率,两种藻单细胞颗粒氮的含量都迅速下降,当进入细胞生长的稳定期后,单个细胞中颗粒氮的含量基本稳定。
比较两图,可发现两种藻单细胞颗粒态氮的变化有所差异,具体来说:两种藻细胞内颗粒氮的含量存在差异,中肋骨条藻单细胞内颗粒氮的含量(指数期约为10-6μmol,平台期约为10-7μmol)要远远小于米氏凯伦藻单细胞内颗粒氮的含量(指数期约为10-4μmol,平台期约为10-6μmol),这种差异与两类藻细胞大小和生存策略有关。作为典型的r-策略者[33],中肋骨条藻藻分裂生长速度很快,细胞密度较大,细胞个体较小,因此每个细胞中颗粒氮的含量较小;而作为典型的k-策略者,米氏凯伦藻的生长速度较慢,细胞密度较小,细胞个体较大,因此每个细胞中颗粒氮的含量较大。此外,两种藻单细胞颗粒氮的含量随尿素比例的变化也有差异:中肋骨条藻随着氮源中尿素比例的升高,单细胞颗粒氮含量升高,其中平台期A组单细胞颗粒氮含量在2.9×10-7μmol左右,E组在5.6×10-7μmol左右。而对于米氏凯伦藻,在平台期单细胞颗粒氮含量基本在2.8—3.0×10-6μmol,随尿素含量的增加,变化不大。同时,实验中观察到中肋骨条藻在平台期E组细胞比较大且成串生长,而A组细胞比较小且分散生长,可能的原因是硝氮和尿素对中肋骨条藻生长的作用机制不同,硝氮主要用于促进细胞的分裂增殖,而尿素主要用于促进细胞形态大小的生长。对于米氏凯伦藻,各实验组细胞形态未观察到明显不同。
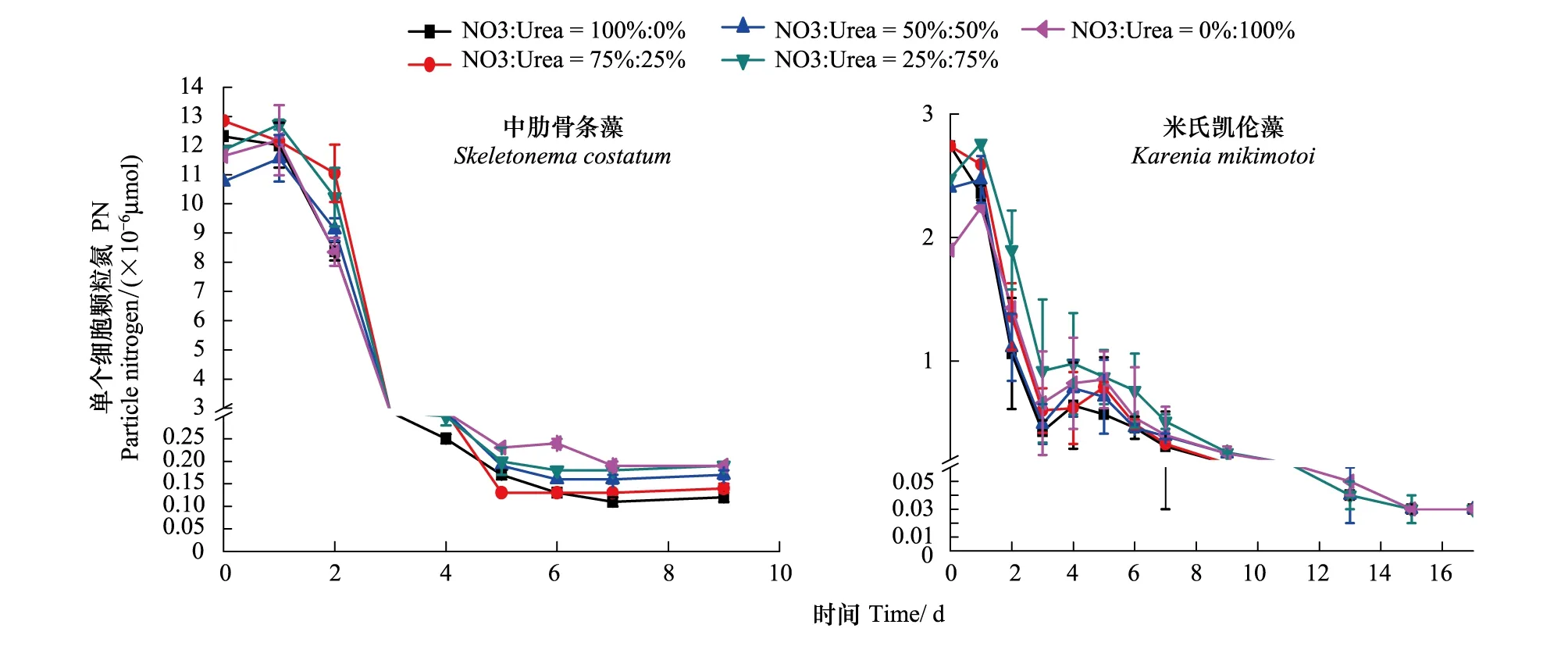
图4 两种藻在培养过程中单个细胞颗粒态氮含量的变化Fig.4 The change of particle nitrogen in average cell of the two kinds of typical algae
3 结论
(1)中肋骨条藻和米氏凯伦藻均能吸收环境中的尿素和硝酸盐,但其吸收利用程度有差异。中肋骨条藻对硝酸盐的吸收利用大于尿素,米氏凯伦藻对尿素有较好的亲和性,在生长过程中能同等的吸收利用硝酸盐和尿素。
(2)中肋骨条藻平台期氮盐浓度为2.5μmol/L左右,米氏凯伦藻平台期时氮盐浓度(1.0μmol/L),低于中肋骨条藻平台期的氮盐浓度,米氏凯伦藻更能适应低浓度营养盐环境。
(3)中肋骨条藻对代谢产生的DON 利用率很低,而米氏凯伦藻能重新吸收代谢产生的DON,这也有助于在硅藻赤潮发生末期,优势种由硅藻向甲藻的演替。
(4)中肋骨条藻细胞内颗粒氮的含量远远小于米氏凯伦藻,中肋骨条藻随着氮源中尿素比例的升高,单个细胞中颗粒氮含量升高,而米氏凯伦藻在平台期单个细胞颗粒氮含量随尿素含量的增加,变化不大。
[1] 孙军, 刘东艳, 王宗灵, 朱明远. 浮游动物摄食在赤潮生消过程中的作用. 生态学报, 2004, 24(7): 1514- 1522.
[2] Glibert P M, Anderson D M, Gentien P, Granéli E, Sellner K G. The global, complex phenomena of harmful algal blooms. Oceanography, 2005, 18(2): 136- 147.
[3] Amon R M W, Meon B. The biogeochemistry of dissolved organic matter and nutrients in two large Arctic estuaries and potential implications for our understanding of the Arctic Ocean system. Marine Chemistry, 2004, 92(1- 4): 311- 330.
[4] 袁美玲, 王朝晖, 李友富. N、P营养盐对海洋卡盾藻(Chattonellamarina)生长的影响. 生态学报, 2008, 28(1): 430- 435.
[5] 王金花, 唐洪杰, 杨茹君, 王修林. 氮磷营养盐对中肋骨条藻生长及硝酸还原酶活性的影响. 海洋科学, 2008, 32(12): 64- 68.
[6] 汪芳, 葛蔚, 柴超, 孟祥霞. 氮磷浓度对东海原甲藻优势种群吸附BDE28和BDE47的影响. 环境科学, 2013, 34(8): 2992- 3001.
[7] Carlsson P, Granéli E, Tester P, Boni L. Influences of riverine humic substances on bacteria, protozoa, phytoplankton, and copepods in a coastal plankton community. Marine Ecology Progress Series, 1995, 127(1/3): 213- 221.
[8] 赵夕旦, 祝陈坚, 举鹏, 史致丽. 胶州湾东部海水中氮的含量和分布. 海洋科学, 1998, 12(1): 40- 44.
[9] 王先伟, 温伟英, 刘翠梅. 珠江口及附近海域夏季氮的化学形式分布研究. 海洋科学, 2003, 27(4): 49- 53.
[10] Glibert P M, Magnien R, Lomas M W, Alexander J, Tan C L, Haramoto E, Trice M, Kana T M. Harmful algal blooms in the Chesapeake and Coastal bays of Maryland, USA: comparison of 1997, 1998, and 1999 events. Estuaries, 2001, 24(6): 875- 883.
[11] 王雪景, 石晓勇, 赵玉庭, 张传松. 2011年春季东海赤潮高发区尿素分布特征及影响因素. 海洋学报, 2015, 37(2): 65- 74.
[12] 李志林, 石晓勇, 张传松. 春季黄渤海海水中尿素分布特征及溶解态氮的组成. 环境科学, 2015, 36(11): 3999- 4004.
[13] 黄爽, 石晓勇, 张传松, 卓书辉. 2010年春季杭州湾邻近海域尿素含量及平面分布. 海洋环境科学, 2012, 31(1): 58- 61.
[14] 徐宁, 王萌, 孙凯峰, 胡章喜, 段舜山. 有机氮对大亚湾亚历山大藻种群生长的促进作用. 中国环境科学, 2012, 32(3): 504- 509.
[15] 胡章喜, 徐宁, 段舜山. 不同氮源对4种海洋微藻生长的影响. 生态环境学报, 2010, 19(10): 2452- 2457.
[16] 陈园, 徐宁, 段舜山. 有机氮比例和光强对赤潮藻球形棕囊藻生长和光合作用的影响. 生态环境学报, 2011, 20(3): 499- 504.
[17] 黄凯旋, 张云, 欧林坚, 吕颂辉, 吕淑果, 齐雨藻. 春季海南岛近岸海域尿素与浮游生物的脲酶活性. 生态学报, 2013, 33(15): 4575- 4582.
[18] Beman J M, Arrigo K R, Matson P A. Agricultural runoff fuels large phytoplankton blooms in vulnerable areas of the ocean. Nature, 2005, 434(7030): 211- 214.
[19] 王金辉, 黄秀清, 刘阿成, 张有份. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 2004, 23(1): 32- 39.
[20] Harrison P J, Waters R E, Taylor F J R. A broad spectrum artificial sea water medium for coastal and open ocean phytoplankton. Journal of Phycology, 1980, 16(1): 28- 35.
[21] 孙军, 宁修仁. 海洋浮游植物群落的比生长率. 地球科学进展, 2005, 20(9): 939- 945.
[22] 钱佐国, 孙明昆, 杨炼锋, 贺深. 测定海水中尿素的双乙酰单肟法研究. 黄渤海海洋, 1984, 2(1): 66- 71.
[23] Heil C A, Revilla M, Glibert P M, Murasko S. Nutrient quality drives differential phytoplankton community composition on the southwest Florida shelf. Limnology and Oceanography, 2007, 52(3): 1067- 1078.
[24] Solomon C M, Glibert P M. Urease activity in five phytoplankton species. Aquatic Microbial Ecology, 2008, 52(2): 149- 157.
[25] 李京. 东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究[D]. 青岛: 中国海洋大学, 2008.
[26] 胡章喜, 徐宁, 段舜山, 李爱芬, 张成武. 尿素对中国近海3种典型赤潮藻生长的影响. 环境科学学报, 2010, 30(6): 1265- 1270.
[27] 黄备, 邵君波, 魏娜, 王益鸣. 2014年春季东海大面积甲藻赤潮的生态特征. 生态环境学报, 2014, 23(9): 1457- 1462.
[28] 周伟华, 殷克东, 朱德第. 舟山海域春季浮游植物生物量及东海原甲藻赤潮频发机制初探. 应用生态学报, 2006, 17(5): 887- 893.
[29] 陈翰林, 吕颂辉, 张传松, 朱德弟. 2004年东海原甲藻赤潮爆发的现场调查和分析. 生态科学, 2006, 25(3): 226- 230.
[30] 吕颂辉, 李英. 我国东海4种赤潮藻的细胞氮磷营养储存能力对比. 过程工程学报, 2006, 6(3): 439- 444.
[31] 庞秋婷. 富营养化对东海两种赤潮优势藻种演替及黄海海区浒苔生长的影响[D]. 青岛: 中国海洋大学, 2013.
[32] 张璇, 石晓勇, 张传松, 韩秀荣. 长江口及邻近海域赤潮藻种演替过程中营养盐特征. 海洋环境科学, 2012, 31(6): 817- 820.
[33] 周名江, 于仁成. 有害赤潮的形成机制、危害效应与防治对策. 自然杂志, 2007, 29(2): 72- 77.
Effect of urea on the growth ofSkeletonemacostatumandKareniamikimotoi
LI Zhilin1,2,ZHOU Yanlei1,2,WANG Xuejing1,2,SHI Xiaoyong1,2,3,ZHANG Chuansong1,2,*
1CollegeofChemistryandChemicalEngineering,OceanUniversityofChina,Qingdao266100,China2LaboratoryofMarineChemistryTheoryandTechnology,MinistryofEducation,OceanUniversityofChina,Qingdao266100,China3NationalMarineHazardMitigationService,Beijing100194,China
With rich nutrients, suitable temperature, light, and weather conditions, large-scale harmful algal blooms (HABs) have frequently occurred in the East China Sea in recent years. According to datas from the State Bureau of Oceanic Administration of China from 2009 to 2014, there were 367 instances of HABs in the coastal waters of China and 195 instances in the East China Sea, accounting for 53%. It is generally believed that the occurrence of algal blooms is linked to eutrophication of coastal waters. Nutrients are the basic material for the growth and reproduction of organisms. As an important kind of DON(Dissolved Organic Nitrogen), urea has high biological activity. Many studies have reported that urea could serve as the sole nitrogen source for a variety of phytoplankton. As such, studying the effect of urea on the organisms that induce red tides is very important. To determine the different responses of diatoms and dinoflagelattes to DON, the effect of urea on the growth ofSkeletonemacostatumandKareniamikimotoi, which are the dominant algae in China, were determined using a batch of culture experiments in the laboratory. Results indicated that the two algae could grow under different ratios of urea to nitrate, but their kinetic parameters were different. With the increase of the proportion of urea in the mixed nitrogen source in the culture solution, the growth rate(0.91—0.82/d) ofSkeletonemacostatumand its maximum biomass(2.0×105—1.2×105cells/mL) in the plateau phase of the growth curve gradually decreased. However, the growth rate (0.36—0.51/d) ofKareniamikimotoigradually increased and its maximum biomass (1.1×104cells/mL) in the plateau phase remained relatively unchanged. In plateau phase, the concentration of nitrogen in theSkeletonemacostatumculture solution declined to 2.5μmol/L, and inKareniamikimotoiit declined to 1.0 μmol/L. In the exponential phase, the concentrations of DON increased with the growth of algae, and reached the maximum values (5—6 μmol/L), then remained constant in the cultural ofSkeletonemacostatum. ForKareniamikimotoi, the concentrations of DON declined after reaching maximum values (2—3 μmol/L) in the culture solution. The content of particulate organic nitrogen in each cell ofSkeletonemacostatum(approximately 10-6μmol in the exponential phase, 10-7μmol in the plateau phase) were much less than that ofKareniamikimotoi(10-4μmol in the exponential phase, 10-6μmol in the plateau phase). In an environment with lower inorganic nitrogen and higher DON concentration, dinoflagellates exhibit better adaptability than diatoms. This research can provide references on the interpretation of the succession between diatoms and dinoflagelattes in the Yangtze River Estuary in spring.
nitrogen;urea;HABs;Skeletonemacostatum;Kareniamikimotoi
国家自然科学基金资助项目(41376106)
2016- 01- 13; 网络出版日期:2016- 12- 19
10.5846/stxb201601130085
*通讯作者Corresponding author.E-mail: zcsong@ouc.edu.cn
李志林,周艳蕾,王雪景, 石晓勇, 张传松.尿素对中肋骨条藻与米氏凯伦藻生长的影响.生态学报,2017,37(9):3193- 3200.
Li Z L,Zhou Y L,Wang X J,Shi X Y,Zhang C S.Effect of urea on the growth ofSkeletonemacostatumandKareniamikimotoi.Acta Ecologica Sinica,2017,37(9):3193- 3200.
