皮蠹科昆虫化石研究进展
2017-06-05邓从双
邓从双,任 东,庞 虹*
(1. 生物多样性演化与保护广东普通高校重点实验室,中山大学生物博物馆,中山大学生命科学学院,广州 510275;2. 昆虫演化与环境变迁重点实验室,首都师范大学生命科学学院,北京 100048 )
皮蠹科昆虫化石研究进展
邓从双1,任 东2*,庞 虹1*
(1. 生物多样性演化与保护广东普通高校重点实验室,中山大学生物博物馆,中山大学生命科学学院,广州 510275;2. 昆虫演化与环境变迁重点实验室,首都师范大学生命科学学院,北京 100048 )
皮蠹科化石主要发现于新生代波罗的海和多米尼加琥珀,以及中生代和新生代欧洲、北美等地的湖相遗迹化石中,近期在白垩纪缅甸琥珀中发现了大量皮蠹科化石。本文回顾了世界皮蠹科昆虫化石研究进展,统计了已发表的化石属种,探讨了皮蠹科昆虫化石分布的地质年代及地层,简要介绍了皮蠹科昆虫的分类系统,并提出了目前皮蠹科化石研究中有待解决的问题。
鞘翅目;皮蠹科;化石;分类
注:本论文使用的数据来自国家动物数字博物馆数据库
皮蠹科Dermestidae隶属于鞘翅目长蠹总科,广泛分布于世界各地(Lawrence and S' lipiński, 2010;Háva, 2015)。皮蠹是一类重要的储藏物害虫,其食性杂、耐干燥、耐饥饿、传播广、繁殖力强,严重为害药材、奶粉、鱼类及肉类加工品、谷类及其加工品、生物标本等动植物材料,造成严重的资源浪费和经济损失,很多种类被我国列为检疫性害虫(张生芳等,2008)。皮蠹科成虫具有区别于其他科昆虫的显著特征:体型较小,长椭圆形,背面隆起,腹面扁平,被覆毛或鳞片,有时形成斑纹;头在前胸背板下几乎可完全伸缩,多数种类具单眼;触角4-11节,棒节通常1-3节,极少数种类多达5-8节;前胸背板基部约与鞘翅基部等宽;前足基节近球形,基节窝后方开放,后足基节非常发达,多具基节板。幼虫圆柱状或轻微弯曲,不呈C型,背面比腹面骨化程度高;头部下口式,咀嚼式口器;触角3节;腹面具穗状刚毛,腹部10节,腹部背板不具横向皱壁。因幼虫形态极为相似,难以区分,目前皮蠹科分类主要以成虫形态研究为主(Lawrence and S' lipiński, 2010;李枷霖等,2014)。
皮蠹科昆虫化石种类较多,尤其是近期在缅甸胡康河谷发现了数量丰富的皮蠹科琥珀化石。研究皮蠹化石可以揭示该类群的起源、演化、繁殖方式和生活习性,有助于推测当时的古地理、古气候和地球演变等重要问题。本文就当前世界皮蠹科昆虫化石的研究现状、存在问题展开综述。
1 皮蠹科昆虫化石研究简史
皮蠹科是鞘翅目中相对较小的一个科,全世界已知仅有1500余种,其中已描述的化石47种(Háva, 2004,2015;Lawrence and S' lipiński, 2010)(表1)。皮蠹科昆虫化石分布广泛,涉及到的地质时期跨度较大,从晚三叠世到中新世的地层均有发现,但产于晚三叠纪的化石的分类位置被Carpenter(1992)质疑。目前有确凿证据的最古老的皮蠹化石发现于中国内蒙古道虎沟地区中侏罗世九龙山组地层中(Dengetal., 2017)。皮蠹科昆虫化石研究记录始于1847年Heer发表的中新世早期化石Dermestespauper。1917年,Cockerell在缅甸琥珀中发现了皮蠹幼虫化石Dermesteslarvalis,将皮蠹科化石地质年代提前到早白垩世。同期,Wickham(1912,1913)和Scudder(1900)报道了美国科罗拉多州早渐新世地层中发现的4块皮蠹科昆虫化石,其中,Orphilusdubius为迄今为止唯一发现的Orphilinae亚科化石。1923年,Dunstan在澳大利亚昆士兰晚三叠世地层发现皮蠹科化石,并建立2属5种。1960年Pierce在美国加州Calico山脉发现中新世皮蠹幼虫Miocryptorhopalumkirkbyae。由于化石分布的局限性,20世纪中后期皮蠹科昆虫化石的研究较少。21世纪以来,随着多米尼亚、波罗的海和缅甸等地大量琥珀的开采,皮蠹科昆虫化石的分类研究逐渐增多。Zhantiev(2006)在乌克兰和波罗的海琥珀中发现始新世晚期皮蠹化石2属3种,并对皮蠹科昆虫的形态特征和系统分类等方面进行了深入研究。Háva(2004,2006,2007,2008,2012,2015)对皮蠹科化石昆虫的分类研究贡献非常突出,共描述、命名了9属23种,并对已发现的部分化石属种进行了总结,为皮蠹科昆虫化石分类奠定了较好的基础。目前皮蠹化石的研究主要集中在对中生代琥珀化石的分类研究(Háva and Prokop, 2006;Hávaetal., 2006, 2007;Zhantievetal., 2006; Kirejtshuketal., 2009, 2010;Rossetal., 2010;Herrmannetal., 2011;Perris and Háva, 2016;Caietal., 2017)。
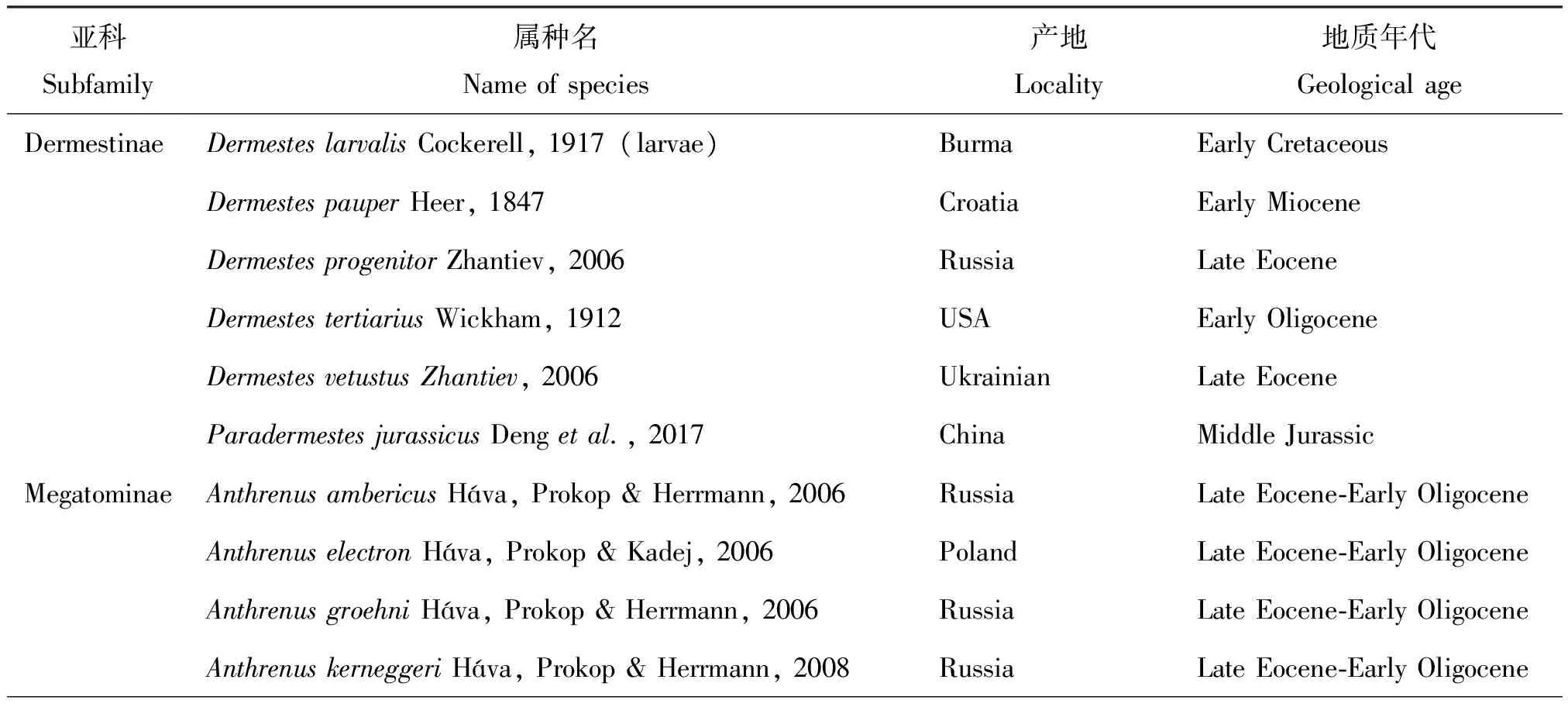
表1 世界已发现皮蠹科昆虫化石名录
续上表

亚科Subfamily属种名Nameofspecies产地Locality地质年代GeologicalageAmberodermabealiHava&Prokop,2004DominicaOligocene⁃MioceneCryptorhopalumambericumHáva&Prokop,2004DominicaOligocene⁃MioceneCryptorhopalumdominicanumHáva&Prokop,2004DominicaOligocene⁃MioceneCryptorhopalumelectronBeal,1972Mexico,DominicaOligocene⁃MioceneCryptorhopalumjantaricumHáva&Prokop,2004DominicaOligocene⁃MioceneGlobicornisrakoviciHáva,2008RussiaLateEocene⁃EarlyOligoceneGlobicornisambericusHáva,Prokop&Herrmann,2006Poland,RussiaLateEocene⁃EarlyOligoceneMegatomaatypicaDengetal,2017BurmaUpperCretaceousMegatomaelectraZhantiev,2006UkrainianLateEoceneMiocryptorhopalumkirkbyaePierce,1960(larvae)USAMioceneOrphilinaeOrphilusdubiusWickham,1912USAEarlyOligoceneTrinodinaeEvorineaambericaHáva,Prokop&Herrmann,2008RussiaLateEocene⁃EarlyOligoceneOisenodesazariKirejtshuk,Háva&Nel2010FranceLowermostEoceneOisenodesclavatusKirejtshuk,Háva&Nel2010FranceLowermostEoceneOisenodesgallicusKirejtshuk,Háva&Nel2010FranceLowermostEoceneOisenodesmetepisternalisKirejtshuk,Háva&Nel2010FranceLowermostEoceneOisenodesoisensisKirejtshuk,Háva&Nel2010FranceLowermostEoceneOisenodestransversusKirejtshuk,Háva&Nel2010FranceLowermostEoceneRinodesrufescensReitter&Háva,2003BalticamberMiddle⁃LateEoceneTrinodespuetziHáva&Prokop,2006RussiaLateEocene⁃EarlyOligoceneAttageninaeAttagenusaboriginalisWickham,1913USAEarlyOligoceneAttagenusambericusHava&Prokop,2004DominicaOligocene⁃MioceneAttagenusbalticusHáva,Prokop&Herrmann,2008RussiaLateEocene⁃EarlyOligoceneAttagenusburmiticusCai,Háva&Huang,2017BurmaUpperCretaceousAttagenusextinctusHeyden&Heyden,1865GermanyMiddleMioceneAttagenushoffeinsorumHáva,Prokop&Herrmann,2006Poland,RussiaLateEocene⁃EarlyOligoceneAttagenusobesusHáva,Prokop&Herrmann,2008RussiaLateEocene⁃EarlyOligoceneAttagenussecundusDengetal,2017BurmaUpperCretaceousAttagenussopitusScudder,1900USAEarlyOligoceneAttagenusturonianensisHáva&Peris,2016USALateCretaceousAttagenusyantarnyiHáva&Bukejs,2012RussiaUpperEoceneCretodermestespalpalisDengetal,2017BurmaUpperCretaceousGeneraReeveanaintermediaDunstan,1923AustraliaLateTriassicuncertainReeveanamajorDunstan,1923AustraliaLateTriassicReeveanaminorDunstan,1923AustraliaLateTriassicTryoniopsisgranulataDunstan,1923AustraliaLateTriassicTryoniopsispunctataDunstan,1923AustraliaLateTriassic
2 皮蠹科的分类系统
皮蠹科在鞘翅目中的系统位置存在较大的争议,其内部系统发育关系也尚未定论。Crowson(1960)首次提出长蠹系包括皮蠹总科和长蠹总科,其中,皮蠹总科分为伪郭公虫科Derodontidae、小丸甲科Nosodendridae、短跗甲科Jacobsoniidae和皮蠹科,长蠹总科分为长蠹科Bostrichidae和蛛甲科Ptinidae。Lawrence和Newton(1982)基于隐肾管的特殊类型、阳茎形状及幼虫下颚缺乏臼齿,将皮蠹科转移至长蠹总科。目前普遍认同皮蠹科、长蠹科、蛛甲科、小丸甲科和Endecatomidae共同组成长蠹总科Bostrichoidea(Lawrence and Newton, 1982;Ivie, 2002;S' lipińskietal., 2011)。依据整合分类信息系统(Integrated Taxonomic Information System, ITIS),长蠹总科包括皮蠹科、窃蠹科Anobiidae、长蠹科、短跗甲科Jacobsoniidae和小丸甲科。国内许多研究人员仍沿用皮蠹总科。Patrice等将皮蠹科分为6个亚科:皮蠹亚科、毛皮蠹亚科、姬皮蠹亚科、斑皮蠹亚科。Zhantiev(2010)将皮蠹科分为3个亚科:皮蠹亚科、光皮蠹亚科、圆皮蠹亚科Anthreninae,并基于皮蠹科成虫、幼虫及蛹的36个形态特征,构建了详细的系统发育树,其中将Trinodini、Trichelodini与Anthrenini归于一支。Háva(2014)修订了Zhantiev的分类体系,将圆皮蠹亚科分为毛皮蠹亚科和斑皮蠹亚科,其余两个亚科不变。Kiselyova和McHugh(2006)基于幼虫形态对皮蠹科进行了支序系统分析,结果表明除Orphilus属之外,皮蠹科的单系性得到支持,并推测其具有起源于劳亚大陆的早于中生代的祖先。但目前大多数皮蠹科系统发育研究未包含化石属种,为皮蠹科的系统分类和演化研究带来了较大的问题。随着各种分子技术手段推动皮蠹科分类鉴定发展的同时,有待更多的化石证据和现生类群介入到皮蠹科的系统发育研究中(李枷霖等,2014)。
3 皮蠹科昆虫化石的地区分布及时代分布特征
3.1 世界皮蠹科昆虫化石的地区分布
世界已知的皮蠹科昆虫化石分布于澳大利亚(Dunstan, 1923)、缅甸(Cockerell, 1917;Caietal., 2017;Dengetal., 2017)、中国(Dengetal., 2017)、克罗地亚(Heer, 1847)、多米尼加(Háva and Prokop, 2004;)、法国(Kirejtshuketal., 2010)、德国(Heyden and Heyden, 1865)、墨西哥(Beal, 1972)、波兰(Hávaetal., 2006a)、俄罗斯(Reitter and Háva,2003;Háva and Prokop, 2006;Hávaetal., 2006a, 2006b, 2008;Zhantiev, 2006;Háva & Bukejs, 2012)、乌克兰(Zhantiev, 2006)、美国(Wickham, 1912)(图1)。
3.2 世界皮蠹科昆虫化石不同地质时代的分布特征
3.2.1 三叠纪(Triassic)
皮蠹科昆虫化石记录贯穿中生代到新生代,具有较为完整的化石记录,对研究皮蠹科化石的起源和演化具有重要意义。皮蠹科昆虫化石最早地质年代始于晚三叠世,Dunstan(1923)在澳大利亚昆士兰州发现了皮蠹化石,简单的基于鞘翅结构将其归于该科,并建立了2化石属5种,但其分类地位有待商榷(Carpenter, 1992)。
3.2.2 侏罗纪(Jurassic)
侏罗纪皮蠹科化石目前仅记录1属1种,发现于内蒙古道虎沟九龙山组(Dengetal., 2017),是已知比较确定的最古老的皮蠹化石(图2),其外型特征与其他化石和现生类群均有所不同,体型硕大,呈宽椭圆形,单眼缺失,后足基节板仅中部发育,并由此建立了一族Paradermestini。
3.2.3 白垩纪(Cretaceous)
白垩纪地层的皮蠹科化石发现于缅甸胡康河谷琥珀及美国新泽西州琥珀中。其中Attageninae亚科3属,Megatominae亚科1属,Dermestinae亚科幼虫1属。大多数白垩纪皮蠹科化石兼具有和现生类群相似的外形特征(Caietal. 2017)。Peris and Háva(2016)通过对中生代皮蠹科琥珀化石的分析,推测皮蠹科特殊的生境是其保持谱系稳定性的重要原因。Poinar和Poinar(2016)在缅甸琥珀中发现了皮蠹幼虫的矛形刚毛,推测其具有和现生矛形刚毛相同的防卫功能,证明矛形刚毛作为皮蠹科幼虫的共有衍征,可追溯到白垩纪。此外,还有大量尚未系统分类的缅甸生物群中的皮蠹化石有待进一步研究。
3.2.4 新生代(Cenozoic)
从古近纪始新世开始,皮蠹科昆虫化石大量出现,主要采自波罗的海沿岸各国(始新世)和多米尼加琥珀(中新世),以及美国科罗拉州(渐新世和中新世)等地。到目前为止,共发表新生代的皮蠹科化石36种,绝大多数现生亚科都已在新生代出现。

图1 世界皮蠹科昆虫化石地区和时代分布Fig.1 The geographical and age distribution of fossil Dermestidae in the world 注:图注数据为发现的化石物种数。Note: Data shows number of fossil species.

Fig.2 Paradermestes jurassicus Deng et al., 2017注:照片和线条图引自Deng et al., 2017;比例尺=1 mm。Note: Photograph and line drawing updated from Deng et al., 2017; scale bars = 1 mm.
4 皮蠹科昆虫化石研究中存在的问题
4.1 化石标本保存不完善及其产地保护不佳
古生物化石是研究生命起源和演化的直接证据,是研究地质历史时期气候变化的主要依据,因此保护古生物化石及其产地具有重要意义。目前,仍然存在化石产地盗采滥挖损毁严重、管理保护机构不健全和受自然风化破坏明显等问题。一些平板化石和琥珀化石,保存的鉴别特征有限,甚至关键特征缺失,无法准确的判断其分类地位。
4.2 化石与现生类群分类准则不同
与化石标本往往保存不完整不同,现生标本通常数量较多且保存完整,两者的分类依据也存在较多差异,一些早期发表的化石种类描述过于简单或不准确,常导致部分化石类群的分类位置需要重新探讨并修订。
4.3 缺乏系统学研究
尽管皮蠹科化石昆虫研究历史已有近200年,由于皮蠹科化石分布面积较广、数量少但种类较多,目前对皮蠹科化石的研究工作大部分基于对化石标本的基本描述与分类,系统学研究工作有待在大量描述新种的基础上加强。
4.4 研究方法有待更新
目前对皮蠹科昆虫化石的研究主要依靠传统光学显微镜,采用先进的拍摄技术,如激光共聚焦显微镜、荧光显微镜、扫描电镜等,可细致全面地观察虫体,提高照片的清晰度,从而更精准的进行分类和系统学研究。
5 皮蠹科昆虫化石研究展望
中国皮蠹科化石研究起步较晚,仅对缅甸琥珀和道虎沟地层中的皮蠹科化石略有研究,在以下几个方面值得期待:(1)继续开展基础分类鉴定工作。中国北方化石资源丰富(任东等,1995,1997,2002;谭京晶等,2004),而目前仅发现1种皮蠹科化石,仍有大量化石种类未被研究,因此有必要继续收集皮蠹科昆虫化石标本,进行基础的分类学研究。(2)深入探讨皮蠹科昆虫的科内系统发育及其与长蠹总科姐妹类群的进化关系,建立完整的系统发育关系。(3)重视与皮蠹科化石研究与相同地质背景下其他因素的结合。皮蠹作为一种尸食性与腐食性昆虫,与同一地质时期中的其他生物具有密切联系。美国研究人员根据恐龙化石上的啃噬痕迹,推断皮蠹科昆虫啃食了1/8的侏罗纪和白垩纪恐龙尸体(Brittetal., 2008)。(4)制定法律法规保护化石产地,加强对皮蠹化石产地和皮蠹化石标本的管理。
References)
Britt BB, Scheetz RD, Dangerfield A. A suite of dermestid beetle traces on dinosaur bone from the upper jurassic morrison formation, wyoming, USA [J].Ichnos, 2008, 15 (2): 59-71.
Cai CY, Háva J, Huang DY. The earliest Attagenus species (Coleoptera: Dermestidae: Attageninae) from Upper Cretaceous Burmese amber [J].CretaceousResearch, 2017, 72: 95-99.
Deng CS, S' lipiński A, Ren D,etal. New Cretaceous Carpet beetles (Coleoptera: Dermestidae) fromBurmeseamber[J]. In Press.
Deng CS, S' lipiński A, Ren D,etal. The oldest Dermestidae beetle from the Middle Jurassic of China (Coleoptera: Dermestidae) [J].AnnalesZoologici(Warszawa), 2017, 67 (1): 109-112.
Grimaldi DA, Engel MS. Evolution of the Insects [M]. Cambridge: Cambridge University Press, 2005: 1-755.
枣棉间作条件下,不同灌水量对棉花产量存在显著影响(图3),适量供水和轻度水分胁迫下产量表现较好,充分供水次之,中度水分胁迫产量最低。M3W3处理产量较M3W1、M3W4分别高62. 5%和13. 9%,M3W2处理产量较M3W1、M3W4分别高55. 6%和9. 0%,M3W4处理产量较M3W1高42. 7%,M3W3处理产量与M3W2处理无显著差异。
Háva J, Bukejs A.Attagenusyantarnyisp. nov., a new species fromBalticamber(Coleoptera: Dermestidae) [J].BalticJournalofColeopterology, 2012, 12 (2): 155-158.
Háva J.Globicornisrakovicin. sp., a new fossil species (Coleoptera: Dermestidae: Megatomini) fromBalticamber[J].Alavesia, 2008, 2: 3-5.
Háva J, Kadej M. Description of a new species ofGlobicornisfrom China (Coleoptera: Dermestidae: Megatominae) with comparison to related species [J].FloridaEntomologist, 2014, 97 (3): 1081-1084.
Háva J, Prokop J, Andreas H. New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber [J].ActaSocietatisZoologicaeBohemicae, 2006, 281-287.
Háva J, Prokop J, Herrmann A. New fossil dermestid beetles (Coleoptera: Dermestidae) from Baltic amber [J].ActaSocietatisZoologicaeBohemicae, 2006, 69: 281-287.
Háva J, Prokop J, Herrmann A. New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber-III [J].ActaSocietatisZoologicaeBohemicae, 2007, 71: 151-157.
Háva J, Prokop J, Kadej M. New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber-II [J].StudiesandReportsofDistrictMuseumPrague-east,TaxonomicalSeries, 2006, 2: 65-68.
Háva J, Prokop J. New fossil dermestid-beetles (Coleoptera: Dermestidae) from the Dominican amber of the Greater Antilles, with an appendix listing known fossil species of this family [J].ActaSocietatisZoologicaeBohemicae, 2004, 68: 173-182.
Háva J, Prokop J.Trinodespuetzisp. nov., a new fossil species described from the Baltic Amber (Coleoptera: Dermestidae) [J].ActaSocietatisZoologicaeBohemicae, 2006, 69: 277-279
Háva J. World Catalogue of Insects Volume 13. Dermestidae (Coleoptera) [M]. Boston: Brill Leiden, 2015:419 .
Herrmann A, Háva J, Kadej M. A new species of Globicornis Latreille in Cuvier (Coleoptera: Dermestidae: Megatominae) from Switzerland [J].StudiesandReportsofDistrictMuseumPrague-EastTaxon, 2011, 7 (1-2): 141-145.
Háva J. World keys to the genera and subgenera of Dermestidae (Coleoptera), with descriptions, nomenclature and distributional records [J].ActaMuseiNationalisPragae,SeriesB,NaturalHistory, 2004, 60 (3-4): 149-164.
Ivie MA. Keys to families of beetles in America north of Mexico. In: Arnett RS, Thomas MC Jr., Skelley PE,etal., eds. American Beetles. Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea[C] .Boca Raton: CRC Press, 2002:1-835.
Kadej M, Háva J. First record of a fossilTrinodeslarva from Baltic amber (Coleoptera: Dermestidae: Trinodinae) [J].Genus, 2011, 22: 17-22.
Kirejtshuk AG, Azar D, Tafforeau P,etal. New beetles of Polyphaga (Coleoptera, Polyphaga) from Lower Cretaceous Lebanese amber [J].Denisia, 2009, 26: 119-130.
Kirejtshuk AG, Hava J, Nel A. New genus and species of subfamily Trinodinae (Coleoptera, Polyphaga, Dermestidae) from Lowermost Eocene French amber [J].ZoosystematicaRossica, 2010, 19 (1): 54-69.
Kiselyova T, Mchugh JV. A phylogenetic study of Dermestidae (Coleoptera) based on larval morphology [J].SystematicEntomology, 2006, 31 (3): 469-507.
Lawrence JF, S' lipiński A. Dermestidae Latreille, 1804. In: Leschen RAB, Beutel RG, Lawrence JF, eds. Handbook of Zoology. Volume IV Arthropoda: Insecta Part 38. Coleoptera, Beetles. Volume 2. Morphology and Systematics (Polyphaga partim) [C]. Berlin: W. DeGruyter, 2010:198-206.
Lawrence JF, S' lipiński A, Seago AE,etal. Phylogeny of the Coleoptera based on morphological characters of adults and larvae [J].AnnalesZoologici(Warszawa), 2011, 61: 1-217.
Li JL, Zheng SZ, Zhan GH,etal. Advances in the molecular biology of Dermestidae insects [J].GenomicsandAppliedBiology, 2014, 33 (5): 1123-1132. [李枷霖, 郑斯竹, 詹国辉, 等. 皮蠹科昆虫分子生物学研究进展[J]. 基因组学与应用生物学, 2014, 33 (5): 1123-1132]
Perris D, Hava J. New species from Late Cretaceous New Jersey amber and stasis in subfamily Attageninae (Insecta: Coleoptera: Dermestidae) [J].JournalofPaleontology2016, 90 (3): 491-498.
Poinar G, Hava J. New Dermestidae (Coleoptera) from Dominican amber [J].Palaeodiversity, 2015, 8: 1-11.
Poinar G, Poinar R. Ancient hastisetae of Cretaceous carrion beetles (Coleoptera: Dermestidae) in Myanmar amber [J].ArthropodStructureandDevelopment, 2016, 45 (6): 642-645.
Ross AJ, Mellish C, York P,etal. Burmese amber. In: Penney D, ed. Biodiversity of Fossils in Amber from the Major World Deposits [C]. Manchester: Siri Scientific Press, 2010: 208-235.
Scudde SH. Adephagous and Clavicorn Coleoptera from the Tertiary deposits at Florissant, Colorado, with description of a few other forms and a systematic list of the non-rhynchophorous tertiary Coleoptera of North America [J].UnitedStatesGeologicalSurvey,Monograph1900, 40: 1-148.
S' lipiński A, Leschen RAB, Lawrence JF. Order Coleoptera Linnaeus, 1758. In: Zhang ZQ, ed. Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness [J].Zootaxa, 2011, 3148: 203-208.
Tan JJ, Ren D, Li NN,etal. Current knowledge of Mesozoic Coleoptera fossils in China [J].ActaZootaxonomicaSinica, 2004, 4: 683-691. [谭京晶, 任东, 李宁宁, 等. 中国中生代鞘翅目化石研究现状[J]. 动物分类学报, 2004, 4, 683-691]
Wickham HF. A report on some recent collections of fossil Coleoptera from the Miocene shales of Florissant [J].BulletinfromtheLaboratoriesofNaturalHistoryoftheStateUniversityofIowa, 1912, 6 (3): 1-38.
Wickham HF. The Princeton collection of fossil beetles from Florissant [J].AnnalsoftheEntomologicalSocietyofAmerica, 1913, 6 (3): 359-366 , pls. 38-41.
Zhang SF, Chen HJ, Xue GH. Atlas of Beetles Associated with Stored Products [M]. Beijing: China Agriculture Science and Technique Press, 2008. [张生芳, 陈洪俊, 薛光华. 储藏物甲虫彩色图鉴[M]. 北京: 中国农业科学技术出版社, 2008]
Zhantiev RD. Ecology and classification of dermestid beetles (Coleoptera, Dermestidae) of the Palaearctic fauna [J].EntomologicalReview, 2009, 89 (2): 157-174.
Zhantiev RD. New species of Late Eocene dermestid beetles (Coleoptera, Dermestidae) from the Rovno and Baltic ambers [J].PaleontologicalJournal, 2006, 40 (5): 560-563.
Progress in the research of fossil Dermestidae (Coleoptera: Bostrichoidea)
DENG Cong-Shuang1, Ren Dong2*, PANG Hong1*
(1. Key Laboratory of Biodiversity Dynamics and Conservation of Guangdong Higher Education Institute, The Museum of Biology, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; 2. College of Life Science, Capital Normal University, Beijing 100048, China)
The fossil Dermestidae are mainly found in the Baltic and Dominica ambers of the Cenozoic, also from lacustrine deposits of North American and Europe. Several amber Dermestidae have recently been found in Burmese amber. Herein, the progress of research on fossil Dermestidae is reviewed. A list of fossil dermestids found worldwide is provided. Geographical and age distribution of fossil Dermestidae in the world are discussed. And phylogenetic research processes are summarized. Some unresolved problems are proposed aiming at development trends.
Coleoptera; Dermestidae; fossil; taxonomy
国家科技基础条件平台工作重点项目(2005DKA21402); 国家标本平台教学标本子平台(http://mnh.scu.edu.cn/)
邓从双,博士研究生,从事昆虫分类与系统进化研究
*通讯作者Author for correspondence,E-mail: rendong@mail.cnu.edu.cn; lsshpang@mail.sysu.edu.cn
Received: 2017-02-28;接受日期Accepted: 2017-03-09
Q961
A
1674-0858(2017)02-0284-07
邓从双,任 东,庞虹.皮蠹科昆虫化石研究进展[J].环境昆虫学报,2017,39(2):284-290.
