赤眼鳟形态特征及其两种同工酶的组织特异性分析
2017-05-30张涛陈建武张林周剑光何力
张涛 陈建武 张林 周剑光 何力

摘要:【目的】從形态特征和生化遗传层面上丰富赤眼鳟种质资源研究数据,同时为筛选出鉴定赤眼鳟种群的生化遗传标记打下基础。【方法】采用形态学测量观察赤眼鳟的形态特征,并以聚丙烯酰胺凝胶电泳检测其不同组织(心脏、眼睛晶状体、肌肉、肝脏和肾脏)中的乳酸脱氢酶(LDH)和苹果酸脱氢酶(MDH),分析两种同工酶表达的组织特异性。【结果】赤眼鳟的眼上缘可见一红斑,鳃耙短而疏,主要形态特征为背鳍D.iii-7,胸鳍Pi.14,臀鳍A.iii-7~8,腹鳍V.i-8,其齿式2·4·5/4·4·2或2·4·4/4·4·2。从赤眼鳟的心脏、眼睛晶状体、肌肉、肝脏和肾脏等5种组织中共检测到7条LDH酶带和6条MDH酶带,不同组织中的酶带数目和活性各不相同,呈明显的组织特异性,且同一组织不同个体间也存在明显差异。【结论】同工酶表达的组织特异性除了受遗传基因调控外,还与各组织的生化代谢活动相关,其中心脏LDH可作为鉴定赤眼鳟种质的生化遗传标记。
关键词:赤眼鳟;形态特征;乳酸脱氢酶(LDH);苹果酸脱氢酶(MDH);组织特异性
中图分类号:S965.199 文献标志码:A 文章编号:2095-1191(2017)12-2281-07
0引言
【研究意义】赤眼鳟(Squaliobarbus curriculus)俗称野草鱼、红眼鱼,原属于江河野生鱼类,除新疆和青藏高原外,我国各大河流湖泊均有发现,主要分布在长江水系和珠江水系,因具有生长快、抗逆性强、肉质细嫩、营养丰富等特点而深受人们喜爱,现已发展成为淡水养殖的主要对象。但近年来因过度捕捞、水资源污染、外来物种入侵及鱼类生境遭到破坏,导致赤眼鳟种质资源锐减(chen et al.,2009),因此研究和保护其种质资源势在必行。【前人研究进展】目前,有关赤眼鳟的研究报道主要集中在生物学特征(龙光华等,2005a;郭丽丽等,2009)、繁殖特性与人工繁殖(龙光华等,2005b,2005c;杨明生等,2005)、杂交育种(任丽珍等,2011;何美凤等,2015;李迪等,2016)及分子遗传特性评估(杨太有等,2008;邵芳等,2013;杨慧荣等,2013)等方面,并已证实赤眼鳟苗种包括仔鱼后期、稚鱼期和幼鱼期3个时期,其中,仔鱼后期卵黄囊消失,开始摄食;稚鱼期出现鳞片至全身批鳞;幼鱼期侧线明显,体形体色与成鱼相似。这些研究结果为赤眼鳟的开口摄食和苗种培育提供了技术参考和指导。同工酶作为一种生化遗传标记,在鱼类的亲缘关系研究、分类和物种鉴定及群体遗传结构分析等方面已得到广泛应用(Verspoor and Moyes,2005;张娟等,2011;Ardestani et al.,2014)。张瑞等(2014)研究表明,皱纹盘鲍(Haliotis discus hannai,♀)与黑足鲍(H.iris,♂)杂交F1代的同工酶主要来自母本皱纹盘鲍。杨凤香等(2015)通过比较白氏文昌鱼(Branchiostoma bel-cheri)、日本文昌鱼(B.japonicum)和佛罗里达文昌鱼(B.floridae)的4种同工酶[苹果酸脱氢酶(MDH)、超氧化物歧化酶(SOD)、酯酶(EST)、淀粉酶(α-AMY)]的酶谱,结果发现这4种同工酶的酶带数目、染色程度及迁移率在不同文昌鱼问的差异较明显,可作为鉴定文昌鱼种群的依据。张龙岗等(2017)研究发现,乌鳢(Ophicephalus argus)和杂交鳢(O.argus ♂×O.maeulata早)的MDH、乙醇脱氢酶(ADH)存在明显的品种差异性,即这两种酶可作为鉴别乌鳢和杂交鳢的新指标。【本研究切入点】至今,尚无涉及赤眼鳟生化遗传特性方面的研究报道。【拟解决的关键问题】采用形态学测量观察赤眼鳟的形态特征,并以聚丙烯酰胺凝胶电泳检测其不同组织中的乳酸脱氢酶(LDH)和苹果酸脱氢酶(MDH),拟从形态特征和生化遗传层面上丰富赤眼鳟种质资源研究数据,同时为筛选出鉴定赤眼鳟种群的生化遗传标记打下基础。
1材料与方法
1.1试验材料
供试赤眼鳟样本均为池塘养殖群体,于2017年3月采自湖北宜昌,共30尾,体重范围6.3~33.1 g/尾,体长范围7.7~12.9 cm/尾。
1.2形态测定
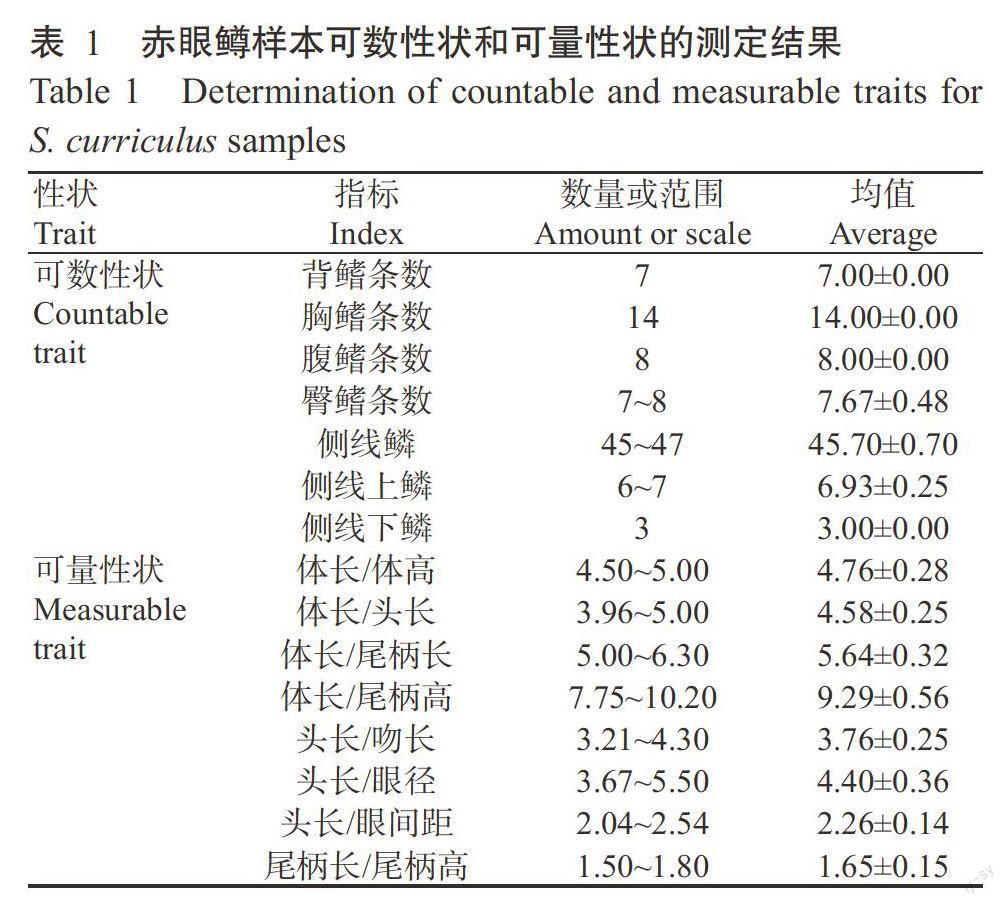
按照GB/T 18654.3-2008《养殖鱼类种质检验》中的相关规定,用游标卡尺(精确度0.1 mm)对样品鱼进行形态测定。其中,可数性状包括背鳍、臀鳍、胸鳍和腹鳍的鳍式、侧线鳞式及左侧第一鳃弓外侧鳃耙数,可量性状包括体长、体高、头长、吻长、眼径、眼间距、尾柄长和尾柄高。
1.3组织酶液制备
参照贺刚等(2012)的方法制备组织酶液。用-剪刀剪除赤眼鳟鳃部后置于水中放血,冰浴条件下趁鱼体尚存活时取心脏、眼睛晶状体、肌肉、肝脏和肾脏等5种组织样品,各组织样品经预冷生理盐水冲洗干净后-80℃保存备用;或直接称重后置于洗净预冷的匀浆器中,按1:3(g/mL)的比例加人预冷双蒸水,冰浴条件下反复研磨至浆状,4℃下12000 r/min离心30 min,重复3次,收集上清液,分装后-80℃保存备用。
1.4聚丙烯酰胺凝胶垂直板电泳及染色
同工酶分析采用聚丙烯酰胺凝胶垂直板电泳,其分离胶浓度7.5%,浓缩胶浓度3.0%,电极缓冲液为pH 8.3的Tris甘氨酸。电泳采用稳压(220 V)方式,电泳时间5 h。电泳结束后取出凝胶板,室温下避光染色,其中,LDH染色参照杨凤香等(2015)的方法,MDH染色参照孟彦等(2009)的方法。待显现清晰条带后以去离子水漂洗凝胶板2~3次,然后将凝胶板平放在自制灯箱上采用尼康数码相机拍照。
1.5统计分析
所测得可量性状数据采用SPSS 20.0进行统计分析。
2结果与分析
2.1赤眼鳟的形态特征
赤眼鳟鱼体延长,略呈圆筒形,腹部圆,后段较侧扁,头呈圆锥形,吻圆钝;口端位,稍向上翘;口裂宽。上颌有两对细小的须;眼大,靠近吻端,眼上缘可见一红斑;鼻孔位于眼前上方。鱼鳞较大,圆形,侧线平直,向后延伸至尾柄正中。背鳍无硬刺,起点与腹鳍相对或略前,背鳍起点至吻端的距离小于至尾鳍基部的距离;胸鳍末端可达胸鳍起点至腹鳍基部距离的3/5处;臀鳍靠后,起点至腹鳍基部的距离大于至尾鳍基鱼的距离;尾鳍分叉深(图1)。赤眼鳟的主要形态特征为背鳍D.iii-7,胸鳍Ri-14,臀鳍A.iii-7~8,腹鳍V.i-8。
赤眼鳟通体银白,但背部颜色较深。体侧每一鳞片后部边缘有黑斑,组成体色纵裂条纹。背鳍、尾鳍深灰色,尾鳍有黑色边缘,其他各鳍灰白色(图1)。鳔2室,前室膨大,后室向尾部逐渐变细,无鳔管。腹膜黑色。鳃耙很短,顶端尖,排列稀疏,鳃丝长。下咽齿3行,齿端呈钩状,齿式2·4·5/4·4·2或2·4·4/4·4·2。赤眼鳟的头长大于体高,吻长大于眼径,且尾柄长约是尾柄高的1.50~1.80倍(表1)。
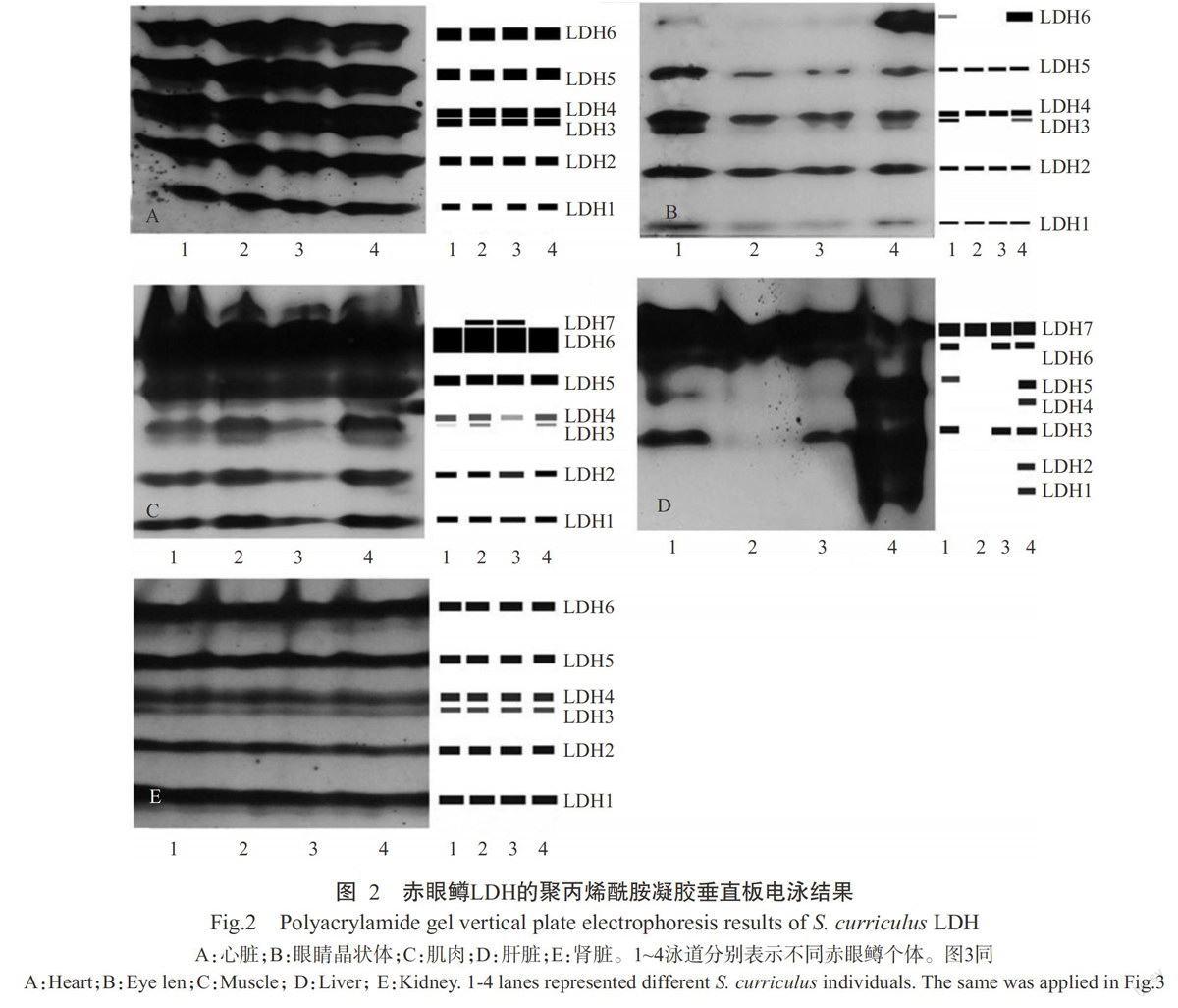
2.2赤眼鳟LDH的表达情况
由图2可看出,从赤眼鳟的心脏、眼睛晶状体、肌肉、肝脏和肾脏等5种组织中共检测到7条LDH酶带,将分子量最小的酶带命名为LDH1,分子量最大的酶带命名为LDH7,不同组织中的酶带数目和活性各不相同,呈明显的组织特异性。其中,从心脏中共检测到6条酶带,以LDH5活性最强,LDH1活性最弱,4尾不同赤眼鳟个体的酶带数目和活性程度基本相同;从眼睛晶状体中共检测到6条酶带,4尾不同赤眼鳟个体的酶带数目和活性程度各不相同,LDH1、LDH2、LDH4和LDH5为共有酶带,1号鱼和4号鱼的酶带数目完全相同,均有LDH3和LDH6;从肌肉中共检测到7条酶带,LDH1、LDH2、LDH4、LDH5和LDH6为共有酶带,LDH6为所有酶带中活性最强的酶带,除3号鱼外其他个体均有LDH3,只是酶带活性有所差异;从肝脏中也检测到7条酶带,以4号鱼的酶带数目最多(7条),2号鱼仅见1条酶带,LDH7为所有酶带中活性最强的酶带;从4尾不同赤眼鳟个体的肾脏中均能检测到6条酶带,且以LDH3的活性最弱。
2.3赤眼鳟MDH的表达情况
MDH分上清液型(S-MDH)和线粒体型(M-MDH)两种类型,分别由两个基因位点控制,为二聚体。由图3可看出,从赤眼鳟的5种组织中共检测到6条MDH酶带,将分子量最小的酶带命名为MDH1,分子量最大的酶带命名为MDH6,其数目和活性也存在明显的组织特异性。其中,从心脏中共检测到3条酶带,MDH3为共有酶带,MDH2活性最强,MDH1活性较弱,1号鱼和2号鱼的带型相同,3号鱼和4号鱼的带型相同;在4尾样本鱼的眼睛晶状体中均未检測到酶带;从肌肉中共检测到6条酶带,MDH3为共有酶带,以2号鱼的酶带数目最多(5条)、3号鱼的酶带数目最少(仅1条);从肝脏中共检测到5条酶带,1号鱼未检测到酶带,2号鱼检测到1条酶带(活性较弱),3号鱼的酶带数目最多(4条),4尾样本鱼中未发现共有酶带;从肾脏中共检测到3条酶带,MDH2为共有酶带。
2.4赤眼鳟生化遗传标记
除了心脏和肾脏外,其余组织的LDH和MDH均呈多态性,不适宜作为鉴定赤眼鳟种质的生化遗传标记。同时考虑到肾脏成分相对复杂,易造成污染,故不建议采用肾脏作为生化遗传标记。心脏组成主要是心肌,其成分相对简单,制备同工酶样本时不易造成提取成分污染。由图4可看出,10尾赤眼鳟样本的心脏LDH均呈单态,即不存在个体差异,且各酶带分离清晰,因此建议采用心脏LDH作为鉴定赤眼鳟种质的生化遗传标记。
3讨论
3.1形态学方法在鱼类种质鉴定中的应用
由于不同鱼类具有不同的形态学特征,观察其外部形态特征,或通过形态测量获取可数性状和可量性状数据,可直观、便捷地进行种类鉴定(丁严冬等,2015)。除传统形态学数据外,近年来还有学者将框架数据运用到鱼类种质鉴定中(宓国强等,2010;许淼洋等,2013)。通过与前人的相关研究结果进行对比分析,发现本研究选取的赤眼鳟与珠江水系赤眼鳟的部分形态特征存在明显差异。本研究中,赤眼鳟的齿式2·4·5/4·4·2或2·4·4/4·4·2,体长为体高的4.50-5.00倍,尾柄长为尾柄高的1.50~1.80倍;而珠江水系赤眼鳟的齿式2·4·5/5·4·2或2·4·4/4·4·2或2·4·5/4·3·2,体长为体高的3.80~4.40倍,尾柄长为尾柄高的1.20~1.40倍(广西壮族自治区水产研究所和中国科学院动物研究所,2006)。造成这种差异的原因除了与样本量及规格差异有关外,可能还与测量方法有关,如可量性状参数的起始点界定,常因研究者的不同而产生差异。此外,本研究挑选的赤眼鳟样本为池塘养殖群体,其饵料来源相对野生群体更丰富,但是否会造成部分可量性状差异,尚需进一步探究。
3.2同工酶表达的组织特异性
本研究结果表明,LDH和MDH两种同工酶在赤眼鳟组织中的表达均具有组织特异性,即某一同工酶在同一个体不同组织器官中的表达类型和程度皆有不同。从赤眼鳟的心脏中共检测到5条LDH酶带,在其肾脏中共检测到6条酶带,]tLDH在心脏中的表达活性整体上强于肾脏。同工酶在组织器官中的表达情况受遗传基因调控,因此部分同工酶基因只在特定的组织或器官中表达。一般认为,脊椎动物同工酶LDH在胚胎发育和细胞分化中具有明显的分化调控模式,表现出高度的发育和组织特异性(张娟等,2011)。从本研究结果来看,赤眼鳟心脏和肾脏中的LDH均为单态,可考虑作为鉴定赤眼鳟种质的生化遗传标记,但在实际操作中采集赤眼鳟肾脏组织时易掺入其他组织而影响研究结果,而心脏组成主要是心肌,成分相对简单,制备同工酶样本时不易造成提取成分污染,故建议采用心脏LDH作为鉴定赤眼鳟种质的生化遗传标记。
3.3同工酶表达的活性强弱
同工酶的表达与组织器官的代谢活动也密切相关。LDH是糖酵解过程中的一种重要酶系,能促使乳酸与丙酮酸问发生可逆的转化(余敏等,2006);MDH是参与三羧酸循环的酶系,在葡萄糖异生和糖酵解过程中发挥重要作用(林文燕等,2010)。本研究从赤眼鳟肝脏中检测出7条LDH酶带,眼睛晶状体中检测到6条酶带,且LDH在肝脏中的表达活性比在眼睛晶状体中的强;在MDH方面,从赤眼鳟肝脏中检测出5条酶带,而在眼睛晶状体中未检测到对应的酶带。肝脏是机体重要的代谢功能器官,具有解毒、储存糖原、促进蛋白合成、分泌胆汁促消化等重要生理作用,因此同工酶在肝脏中的普遍表达与其行使复杂、重要的生理功能密切相关。
3.4同工酶表达的个体差异
同工酶在同一物种不同个体相同组织中的表达差异可能与其生长发育阶段、健康状态、地理环境(生境)等均有关联。薛俊增(2002)研究发现,病蟹和健康蟹的同工酶分离图谱存在明显差异,如健康蟹的心脏和肌肉LDH均有2条谱带,而病蟹只有1条谱带。贾守菊等(2004)研究表明,中华绒螯蟹不同发育阶段ADH的酶带数目及其活性均呈减弱趋势,而EST在抱卵蟹时的酶带数最多、活性最强。余敏等(2006)研究发现,云南高背鲫鱼心脏EST-5在10尾样本鱼中仅部分表达,存在明显的个体差异。黄孝湘等(2012)推测,造成不同水域克氏原螯虾同种组织LDH酶带数目差异的原因是水质问题。本研究结果表明,除了心脏和肾脏外,其余组织的LDH和MDH均呈多态性,如2号鱼肌肉中检测到7条LDH酶带,而3号鱼肌肉中仅检测到6条酶带,其差异可能与不同个体的大小或健康状况有关,但具体原因有待进一步探究。
4结论
同工酶表达的组织特异性除了受遗传基因调控外,还与各组织的生化代谢活动相关,其中心脏LDH可作为鉴定赤眼鳟种质的生化遗传标记。
