群体感应抑制及其植物病害防控应用研究进展
2017-05-30周静煌严准高春生余永廷李智敏曾粮斌陈佳郭利桃严理程毅
周静煌 严准 高春生 余永廷 李智敏 曾粮斌 陈佳 郭利桃 严理 程毅
摘要:许多植物病原菌致病机制依赖于病原微生物群体感应系统的调控,基于群体感应抑制的原理筛选高效的群体感应抑制剂有望成为解决植物病害防治问题的有效途径。文章介绍了群体感应抑制的研究策略,主要包括阻断群体感应信号分子的产生、促进信号分子降解及抑制信号分子与受体结合;综述了近年来主要的群体抑制剂种类及其在植物病害防治上的应用情况,指出群体感应抑制剂并不妨碍病原菌的正常生长代谢,而是抑制群体感应系统,减弱病原菌的毒力,降低其致病性。提出今后应加强群体感应抑制剂筛选和基于群体感应系统淬灭机制的植物病害防治研究,為植物病害防治提供绿色安全的防控措施。
关键词:植物病害防治;群体感应;信号分子;群体感应抑制剂
中图分类号:S432 文献标志码:A 文章编号:2095-1191(2017)12-2197-07
0引言
植物病害防治一直备受关注,一些新的防治方法也不断出现,目前最主要的防治方法包括植物进出口检疫、抗病品种培育、农业栽培控制、化学药剂防治、生物防治和生防机理治疗等。生产上最常用的方法是化学防治,该法强调彻底灭杀病原物,然而,由于化学药剂的长期大量使用,导致很多病原菌产生了耐药性(Roy et al.,2011),对今后的植物病害防治是巨大挑战,因此,研究更安全的病害防治策略尤为必要。许多植物病原菌致病性的形成受到群体感应(Quorum sensing,QS)系统的影响,致病菌定殖到宿主体内后,在Qs机制调控下激活与致病相关基因的表达而产生各种毒力因子,导致宿主发病(程古月等,2012)。此外,由于QS调节,病原菌侵染宿主后能形成生物被膜抵抗宿主的免疫,并产生对药剂的抗性。因此,微生物QS系统的调控通路有望成为病害防治的新靶点(韦柯,2007)。
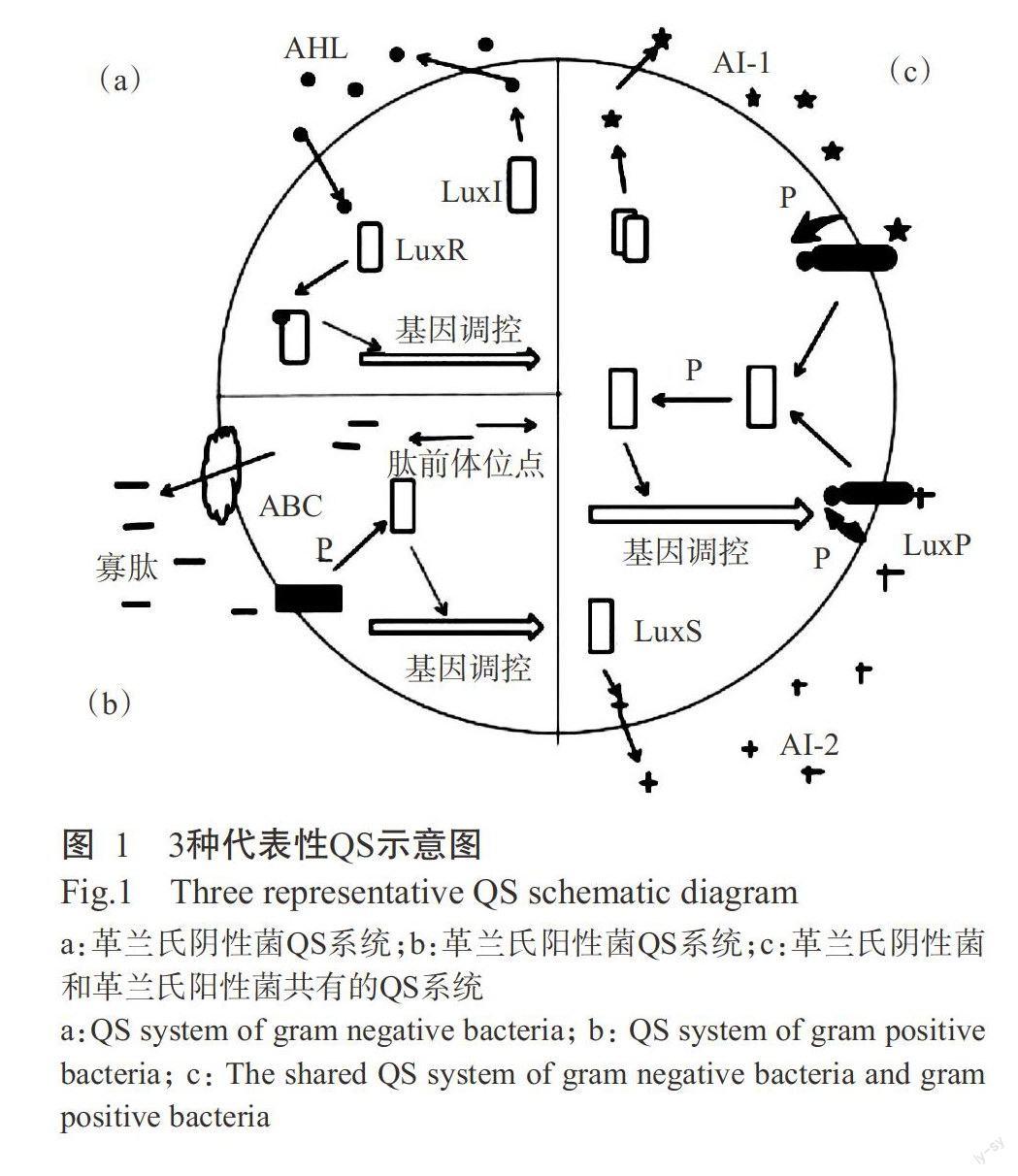
Fuqua等(1994)发现许多微生物的生理生化特征会随着群体密度增加而发生改变,能够分泌毒素、形成生物膜、产生抗生素、生成孢子、发射荧光等,以适应环境的变化,是微生物QS系统的自我调节(程古月等,2012)。在特定环境下,微生物产生并向环境中释放一种被称为自体诱导物(Auto-inducer,AI)的细胞外信号分子,随着个体密度的增大,信号分子也会成比例增加,当信号分子积累到一定浓度时,会诱导微生物产生独特的、多样的群体行为,这一现象称为QS(韦柯,2007)。微生物Qs所分泌的信号分子有3种,故将OS系统分为3类(图1):(a)革兰氏阴性菌的酰基高丝氨酸内酯(N-acvl-homoserine lac-tones,AHLs)型系统。LuxI是可催化合成AI的胞内蛋白酶,其催化带有酰基载体蛋白的酰基侧链与S-腺苷蛋氨酸上的高丝氨酸结合成酰化高丝氨酸内酯化合物(Acyl-homoserine lactone,AHL)。(b)自体诱导肽(Auto-inducter peptide,AIPs)系统,存在于革兰氏阳性菌中。AIP是一种寡肽,虽是小分子多肽,但需经过ABC运转系统(ATP-binding-cassette)或其他膜蛋白通道作用才能穿过细胞壁并行使功能(韦柯,2007)。(c)自诱导物Ⅱ类分子(AI-2s)系统。AI-2是革兰氏阳性菌和革兰氏阴性菌共有的信号分子,是调节细菌行为通用语言。AI-2的产生依赖于LuxS蛋白(Bassler et al.,1993),如失活LuxS基因则不产生AI-2(XavierK and Bassler,2003)。因此,可从植物病原微生物Qs的信号分子着手,减少由QS机制调节的致病性的产生,达到防治植物病害的效果。筛选高效的群体感应抑制剂(Quorum sensing inhibi-tors,QSIs)有望成为防治植物病害的一条有效途径。
正是由于病原菌QS能调节其致病性的产生,作为一种新型的病害防治策略,QS调控系统越来越受到国内外学者重视。近年来,国内外对QSIs的研究发展迅速,并取得了一些进展。了解并有效地利用QS机制来控制病原菌生物膜的形成、毒力因子的产生、致病基因的表达、病原菌问相互抑制或竞争(梁新焱和阮海华,2015)等无疑为抵御病菌感染、植物病害和细菌耐药性研究提供了一条新途径。目前,AHLs降解酶已开始广泛应用于植物病害防治上;同时,已有研究表明抗生素的使用可有效钝化抑制QS系统,从而达到缓解软腐病发生的作用(葛玮珍,2013)。随着上述研究的深入,传统化学药剂防治植物病害的瓶颈将有望被打破。
1 QS抑制的主要策略
基于上述QS调控原理,干扰病原微生物的QS系统的途径有3种。WAHL系统为例,包括:(1)合成信号分子AHL底物的类似物来阻断信号分子的形成;(2)促进AHL的降解;(3)抑制AHL与受体结合。
1.1阻断信号分子产生
AHL的生物合成通常涉及一系列使用S-腺苷蛋氨酸(S-Adenosyl methionine,SAM)作为氨基供体生成高丝氨酸内酯环的一部分反应(Miller and Bassler,2001),酰基载体蛋白(Acyl cartier protein,ACP)作为前体n-AHL的侧链分子。基于以上知识,合成了SAM类似物如丁酰-S腺苷甲硫氨酸、全酰基载体蛋白、西尼毒素和L/D-S-腺苷高半胱氨酸等,研究显示这些物质具有抗Os的活性(Scutera et al.,2014)。但目前仅在人体医学上有应用,极少有进行植物病害防治的报道。
1.2阻止信号分子与受体蛋白结合
通过拮抗剂分子抑制信号分子的接收是另一个防控QS介导的植物病害机制,这些拮抗剂分子与信号分子竞争,干扰信号分子与受体结合。二酮哌嗪(Diketopiperazine,DKP)作为AHL类似物可与受体蛋白结合,从而阻止信号分子与受体蛋白结合,抑制QS系统所调节的基因的表达,使病原菌的致病性降低(Kalia and Purohit,2011)。由海洋红藻分泌的卤化呋喃也是与信号分子竞争并降解受体蛋白的AHL信号分子类似物,卤化呋喃能有效抑制胡萝卜软腐果胶杆菌(Pectobacterium carotovorum)致病因子的表达,降低其对寄主的侵染能力(Hentzer et al.,2003)。
1.3信号分子的降解
在QS机制中AHL分子达到特定浓度是引发病原菌表达致病因子的关键。因此,如果能降解AHLs,使其无法达到浓度阈值,将很大程度削弱植物病原菌的致病性。虽然不同AHLs具有不同碳链长度和不同的酰基侧链,但均以高丝氨酸为主体(韦柯,2007),具有相似的调控基因表达的机制。根据AHLs的结构特点,理论上有4个作用位点可分解AHLs,分别是内酯酶(AHL-lactonase)位点、脱氢酶(AHL-decarboxvlase)位点、酰基转移酶(AHL-acyl-ase)位点和脱氨酶(AHL-deaminase)位点(张丽群等,2010)。目前已在大量原核生物和少数真核生物中发现QS信号分子的降解酶。研究表明,群体猝灭(Quorum quenching,QQ)酶水解AHL中酰胺键或内酯环中的任何一个。降解AHLs很大程度上受AHL酰基转移酶和AHL内酯酶等的调节(Lin et al.,2003)。在病原菌或寄主植物中表达AHL降解酶可破坏病原菌的QS系统,从而达到防治植物病害的效果。
1.4基于QS的植物病害防治策略
根据植物对病原菌QS系统的反应,可采用以下几种策略防治由病原菌QS造成的植物病害。(1)植物本身能夠合成QSIs从而降低病原菌的侵染力。如从弗吉尼亚栎的叶子和假紫斑大戟气生部分提取出能有效抑制根癌土壤杆菌(Agrobacterium tumefa-ciens)QS的活性物质(Adonizio et al.,2006)。虽然已从许多植物中提取出QSIs活性物质,却很少应用于植物病害防治,但随着研究的深入,一旦从植物中提取出具有信号分子抑制活性的物质,即可通过转基因技术达到防治植物病害的目的。(2)合成QS信号分子降解酶,使由QS调控的致病基因无法正常表达。植物自身很少产生信号分子降解酶,但目前已从一些微生物中发现AHLs降解酶,其中,芽胞杆菌产生的内酯酶aiiA应用最广,将aiiA蛋白转入植物中,发现植物对欧文氏菌具有较好的抗性(Dong et al.,2001;Reimmann et al.,2002)。(3)根据天然QSis的结构人工合成QSIs,并使其在植物体内大量表达,抑制侵染植物的病原菌的致病力。(4)利用信号分子在植物体内的微量表达刺激植物免疫系统发生反应。宋水山(2010)用几种细菌信号分子AHL处理几种欧文氏菌常见寄主,发现处理后植物体内与自身防御反应相关的酶的活性提高;拟南芥能较好地防御由软腐欧文氏菌引起的软腐病。若将细菌AHL合成酶基因转入植物,植物可合成QS信号分子,在病原菌尚未产生QS前就能控制信号分子AHL的作用,提高了植物的抗性。如表达胡萝卜软腐欧文氏菌的N-3-氧代己酰基高丝氨酸内酯合成酶基因expI的转基因烟草对欧文氏菌的抗病性提高(宋水山等,2005)。
2 QSIs种类及应用
QSIs以QS分泌的信号分子为靶标,不会影响病原菌的正常活动,极大地减少了病原菌耐药性的产生。根据QSis分子的性质,可将其分为非肽类小分子化合物、肽类化合物(主要是AIPs同系物)和蛋白质(主要为QS猝灭酶)(程古月等,2012)。
2.1非肽类小分子化合物
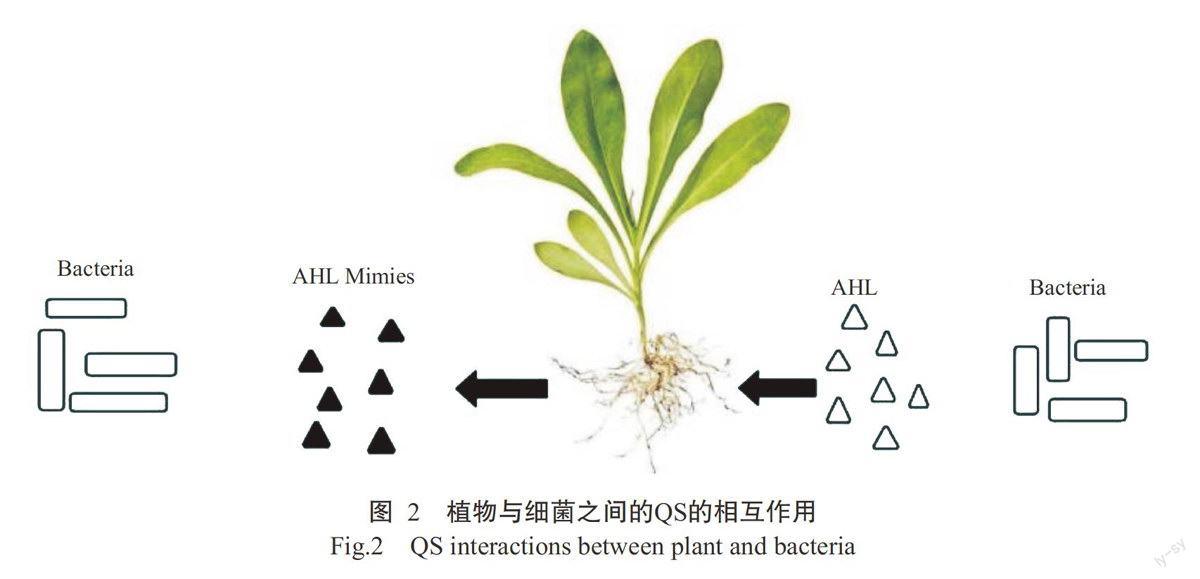
2.1.1天然来源OSis 植物可产生QSI活性以降低侵染菌的致病能力(图2),病原菌释放AHL分子导致生物被膜及冠瘿形成。另一方面,这些植物会对细菌产生的信号分子作出反应,通过释放AHL模拟化合物用于防御,进而影响细菌的QS系统(Kalia and Purohit,2011)。目前已有许多文献报道了从自然植物资源中可分离获得QSIs,由于这些植物可被人类消耗,因此具有QS抑制活性的化合物被认为具有安全,不会对人类细胞造成毒害作用。最先发现具有抑制QS活性的天然产物是由海洋红藻(Delisea pulchra)产生的卤代呋喃酮,研究发现卤代呋喃酮在几个革兰氏阴性菌中通过干扰信号分子AHL与受体蛋白LuxR的结合来干涉AHL调控QS(Chong-Lek et al.,2013)。
除海洋红藻外,从一些高等植物中也分离获得QSI活性物质,但目前从植物中提取出的QSls大多用于医学上,很少用来防治植物病害。因此,了解植物产生的QSIs活性物质及其作用机理,将植物本身产生的QSIs应用于植物病害防治将是一个重要的研究方向。
自然界中一些拮抗菌能通过营养竞争作用、产生抗生素、诱导植物本身的防御系统及重寄生作用来干扰病原菌QS,因此,拮抗菌分泌的抗生素和次级代谢产物可成为一种重要的天然QSIs。研究发现,铜绿假单胞菌分泌的抗生素及其次级代谢产物吩嗪类化合物能有效防治小麦全蚀病,对水稻枯萎病病原菌、辣椒疫菌和黄瓜炭疽病菌也具有抑制效果(孔丽萍,2008)。
2.1.2人工合成QSIs 虽然许多天然化合物具有QSIs活性,但其产量并不足以满足大范围的植物病害防治,因此人们根据天然溴化呋喃酮的结构对取代基和侧链长度稍加改动,设计了具有QSIs活性的化学分子库(程古月等,2012)。如Manny等(1997)合成的如呋喃酮56可阻断AHL分子与受体蛋白结合,并已取得明显的结果。另外,C5O2Br2和CsHOzl3r能通过加速LuxR的周转代谢来抑制QS;C9H10O2Br2能抑制基于AHLs和AI-2s的OS系统。目前,大部分人工合成的抑制剂主要对哺乳动物致病菌的QS有效,对植物病原菌的QS抑制研究甚少,因此有待进一步探究,以确认人工合成QSIs对植物病害的防治效果。
除上述信号分子结构类似物外,人们根据天然的抑制剂分子结构从一些小分子化合物中筛选出一系列能有效抑制病原菌QS的单体化合物,如吡咯酮类化合物。Kaufxnann等(2005)发现吡咯二酮化合物能抑制生物被膜的形成,干扰假单胞菌进行群体交流。陈卫民等(2009)的研究结果显示,吡咯酮化合物是比呋喃酮更有潜力的QSIs的先导化合物。
2.2肽类化合物
在QSIs中,最常见的肽类化合物是AIPs同系物。AIP是一种小分子多肽,是革兰氏阳性菌的自诱导分子。Mayville等(1999)发现AIP分子的尾部发生改变只能影响细菌本身二元信号转导(agr)系统的活化,但依然能抑制跨细菌种属的QS。基于这一原理,人们设计了一些尾部结构不同的AIPs同系物作为QSIs。如尾部丙氨酸被修饰的几种AIPs类似物可有效抑制跨细菌种属的QS系统(Mayville et al.,1999)。一个只含有硫内酯环结构的All肽既能感染病原菌群体内的QS,也能抑制群体问的QS(Lyon et al.,2000)。通过对AIP分子三维构效关系的研究,Lyon等(2000)发现TrAIP-Ⅱ可破坏信号传导,抑制细菌QS,从而降低细菌的毒力。除了环状的AIP肽外,一种线状的RNAⅢ抑制肽(RIP)能减弱病原菌的毒力(Balaban et al.,2005)。
2.3 QQ酶
一些细菌能产生一种降解AHLs的酶——AHL内酯酶,其能水解酰基高丝氨酸内酯环(Scutera et al.,2014)。Dong等(2001)研究表明,一种芽孢杆菌能产生酰基高丝氨酸内酯酶基因aiiA基因,其编码的蛋白质酶能降解胡箩卜欧文氏菌产生的AHL,极大程度上减弱了该病菌的致病性。铜绿假单胞菌(Pseudomonas aeruginosa)PAO1中 B.cereus菌株A24 aiiA的表达会弱化一些QS控制的功能,如AHL积累和毒力因子的表达及分泌(Reimmann et al.,2002)。Mei等(2010)也研究发现,表达AHL内酯酶基因aidH的Pectobacterium carotovorum内AHL信号分子产量极大减少,其在一些主要寄主上的致病力也显著下降。近年来,我国也开展了AHL信号降解酶的研究,发现植物青枯菌(Ralstonia solanacearum)菌株中的aac基因编码的蛋白能有效降解细菌QS信号分子(张争等,2008)。转入aiiA基因的西甜瓜细菌性果斑病菌(Acidovorax citrulli)菌株信号分子的产生显著降低,果斑病症状明显减轻(陈涛等,2008);该基因的转化植株也能防御P.carotovorum引起的软腐病(柴鑫莉等,2007;马宏等,2009)。
在发现QQ内酯酶aiiA后不久,Leadbetter和Greenberg(2000)分离出一种能利用AHL作为能量和氮源的Vanovorax paradoxus菌株,降解该AHL需要AHL酰基转移酶。AHL酰基转移酶随后也在P.aeruginosa、Ralstonia strains、Comamonas sp.、She-wanella sp.和Ochrobactrum sp.等中被鑒定出来(Su-sanne,2015)。因为AHLs有不同的酰基链长度,其为AHL酰化酶提供易变性(Lin et al.,2003)。不同酰化酶对不同长度AHL的作用效果也不同。真氧产碱杆菌中的AHL酰化酶AiiD对超过8个碳的长链AHLs更有效(Ren et al.,2002)。相反,链霉菌M664中的AHL酰化酶在链长度少于8个碳时具有高效性(Park et al,2005)。许多AHL酰化酶等位基因有降解长链AHLs的偏好。然而,Uroz等(2007)从毛单胞菌中发现有不同活性的酰化酶,能降解酰基链长度4~16碳的AHLs。
一系列的研究已显示红球菌(Rhodococcus)有QQ能力。红平红球菌(R.erythropolis)W2拥有AHL内酯酶和AHL酰化酶活性。在植物内,红平红球菌能明显减少马铃薯块茎中果胶杆菌属Carotovorumsp.的致病性(Kiran et al.,2008)。近年来发现有的真菌也能降解AHLs酶,如Phialocephala、Ascomyce-tes和Melinimyces已被证实拥有作用于C6HSL和30C6HSL内酯环的内酯酶活性(Kalia and Purohit,2011)。
3展望
作为一种新型的植物病害防治策略,基于QS系统调控的植物病害防治途径不影响病原物的正常生长,只是调控病原菌致病因子的正常表达,使病原菌不能侵染寄主或降低致病力,从而有效控制植物病害,且不易使病原菌产生抗性(Scutera et al.,2014)。理论上,QSis与传统的化学防治试剂相比具有以下优点:(1)QSIs并不妨碍病原菌的正常生长代谢,而是抑制QS系统,减弱病原菌的毒力,降低其致病性。这样既可有效抑制植物病害,又能缓解病原菌产生耐药性,而且能减少对有益菌的杀伤。(2)由于作用机制不同,QSis可与其他农用抗生素联用,以增强抗菌疗效,减少化学试剂的用量,在有效治疗相关病害的同时,减少环境污染及防治费用(孙琦,2014)。
自发现可利用病原微生物QS以防治植物微生物病害以来,科学家们对OS抑制的研究从未停歇。大量研究表明,向植物中转AQS信号降解酶基因培育出的转基因植物能较好地防治植物病害,具有很好的应用前景。如转基因植物对果胶杆菌引起的软腐病已表现出明显的抗性。除了研究转基因植物,向植物体内注入QSIs,在病原菌侵染宿主前即抑制信号分子的作用,也能有效防治植物病害。植物本身就能产生QSis,但合成量较少。越来越多人工合成的QSis应用于实际生产,初步解决了自然资源中QSis合成量很少的不足。
在今后的研究中,利用现有的理论基础开展QSis筛选和基于QS系统淬灭机制的植物病害防治有望成为植物病害防治工作的重要方向之一,为此应加强以下方面的工作:(1)拓宽QSIs研究范围,使其在革兰氏阳性菌和革兰氏阴性菌中均能广泛应用。目前的QSis主要针对革兰氏阴性菌的AHLs,而针对AT-1、AI-2及最新发现的信号分子AI-3(陈伟,2011)的抑制剂很少被报道。(2)加大QQ酶的研究力度。在多种QSIs中,QQ酶的毒性最小,被认为是最安全有效的酶类(范兴辉等,2017)。(3)AHLs的QQ酶在植物病害防治中存在一些需要解决的问题,如淬灭酶在植物体内的稳定性、催化效率、底物特异性、在应用过程中是否存在副作用及预防措施等。(4)植物能监测到QS信号分子的存在,并作出相应的应激反应(Kalia and Purohit,2011),但目前对有关植物产生的各种QS信号分子类似物的化学结构和性质了解较少,关于AHL信号在植物体内的信号转导途径及信号分子受体及其与受体的特异性结合的研究也不常见。为全面了解植物体内受AHL影响而发生变化的生理生化特性及相关基因的表达,还需开展更深入的研究。(5)QS现象广泛存在于各种微生物中,但目前只了解部分模式细菌群体的QS及其抑制方法,对于一些重要致病细菌及真菌QS系统的研究并不深人,因此应加强这方面的研究,如真菌致病性与QS的联系、QS信号的区分、QSis的筛选等,为大量植物细菌或真菌病害提供新型绿色安全的防控措施。
