矮秆小粒水稻潇湘矮的形态学与分子遗传学分析
2017-05-19吕育松谢耘丰圣忠华邬亚文唐绍清胡培松魏祥进
吕育松谢耘丰圣忠华邬亚文唐绍清胡培松魏祥进,*
(1中国水稻研究所国家水稻改良中心/水稻生物学国家重点实验室,杭州310006;2杭州师范大学生命与环境科学学院,杭州310036;#共同第一作者;*通讯联系人,E-mail:weixiangjin@caas.cn)
矮秆小粒水稻潇湘矮的形态学与分子遗传学分析
吕育松1,#谢耘丰1,2,#圣忠华1邬亚文1唐绍清1胡培松1魏祥进1,*
(1中国水稻研究所国家水稻改良中心/水稻生物学国家重点实验室,杭州310006;2杭州师范大学生命与环境科学学院,杭州310036;#共同第一作者;*通讯联系人,E-mail:weixiangjin@caas.cn)
【目的】研究揭示潇湘矮矮秆小粒的遗传机制,为潇湘矮的育种利用提供理论基础。【方法】利用水稻育种过程中自然突变而得到的稳定遗传的矮秆小粒水稻材料潇湘矮,对其进行农艺性状考查、赤霉素(GA3)和油菜素内酯(BR)敏感性分析、遗传学分析,并利用潇湘矮与其近等基因系NIL(NIP)衍生的F2群体对控制矮秆小粒的基因xxa进行图位克隆,最终利用转基因互补试验验证候选基因。【结果】潇湘矮除表现为矮秆小粒外,其穗长变短,穗型紧凑,千粒重极显著下降。不同浓度梯度的赤霉素(GA3)和油菜素内酯(BR)处理,发现潇湘矮对GA3部分敏感,而对BR不敏感。遗传分析发现其符合孟德尔3∶1分离规律。图位克隆将xxa基因定位于第5染色体InDel标记F81和F82之间约70 kb区间的物理距离内。该区间包含8个开放阅读框(ORF)。其中,第5个ORF(LOC_Os05g26890)被注释为水稻株高基因D1。序列分析发现,潇湘矮的D1基因在第5和12外显子分别有1个碱基的无义替换和3个碱基的缺失,其中第12外显子3个碱基的缺失导致1个赖氨酸的缺失。转基因互补显示D1可以恢复潇湘矮的表型。【结论】潇湘矮控制株高的途径可能与GA3代谢有关;潇湘矮矮秆小粒表型符合单隐性核基因控制的遗传规律;潇湘矮矮秆小粒性状是由D1基因突变所致。
水稻;矮秆小粒;图位克隆;机械化制种
水稻矮秆突变体对于研究水稻株高的调控机制具有重要的意义。到目前为止已经有60多个的水稻矮秆突变体被鉴定[5]。很多矮秆突变体除了表现为矮秆之外,还表现出一些其他表型特征,如小粒、多分蘖、窄叶、卷叶等[5]。水稻矮秆、半矮秆基因主要与植物激素赤霉素(GA)、油菜素内酯(BR)、独脚金内酯素(SLs)等的合成与信号传导途径有关[6-9]。其中,“绿色革命”中使用的半矮秆栽培品种,就是由于赤霉素的合成或信号传导途径基因改变,如sd1属于赤霉素的生物合成途径缺陷突变体,gid1、d1等属于赤霉素信号传导缺陷突变体[8-9]。SD1参与赤霉素的生物合成,该基因编码赤霉素合成途径中关键酶GA20氧化酶,该基因的突变导致水稻不同程度矮化[10-11]。GA代谢途径中,受体蛋白GID1通过降解DELLA蛋白介导水稻中GA的信号传导。gid1突变体表现株高严重矮化、不育等性状[12]。Dwarf 1(D1)参与了依赖G蛋白的GA信号传导,是水稻中第一个鉴定出的GTP结合蛋白(G蛋白)α亚基。d1突变体多表现为水稻株高矮化,茎秆增粗;叶片短且宽,谷粒小而圆,节间密集等。随着研究的深入,功能也逐步清晰。异三聚体G蛋白存在于多种真核生物中,由G蛋白α亚基(D1)、β亚基和γ亚基3个亚基构成。在通常状态下,这3个亚基结合在一起,直到一个配基被G蛋白偶联受体(GPCR)检测到,一起参与GA应答以及信号传导[13-14]。此外,水稻矮秆突变体d2、d11、d61等被证实为BR生物合成或信号传导相关基因发生突变所致[15-17];而d27、d10、d14、d3、d53则被证实为与SLs生物合成或信号传导相关基因发生突变所致[9,18-19]。水稻株高除受一些主基因控制之外,也受大量微效数量基因控制,关于水稻株高相关的QTL定位也有大量研究报道[20-22]。因此,水稻株高调控具有极其复杂的遗传机制,鉴定、解析新的矮秆、半矮秆资源仍然是分子生物学和遗传育种学的关注热点。
本研究以矮秆小粒水稻潇湘矮为研究对象,从形态学、生理学等角度分析了其矮秆小粒特征,并对其矮秆小粒基因进行了图位克隆和功能验证,为潇湘矮在水稻育种中的应用潜力提供理论依据。
1 材料与方法
1.1 试验材料
矮秆小粒水稻潇湘矮(Xiaoxiang’ai,XXA)为水稻育种中自然突变产生,具有稳定的矮化、小粒表型[23]。潇湘矮与半矮秆品种日本晴、02428、热研1号和Y01衍生的F1及F2群体用来对矮秆小粒基因进行遗传分析。从日本晴与潇湘矮的F2群体中选高秆单株连续与潇湘矮回交3代并自交,选择出株高类似日本晴的BC3F2单株为潇湘矮的高秆近等基因系NIL(NIP)。矮秆小粒基因定位群体为潇湘矮与NIL(NIP)杂交的F2群体。所有材料均于正季种植于中国水稻研究所杭州富阳试验基地,种植与管理方法同大田生产。
1.2 外源赤霉素(GA3)和油菜素内酯(BR)处理
NIL(NIP)和潇湘矮种子用1%次氯酸钠消毒,浸种、催芽各24 h后,选取芽长一致的20粒种子
于含梯度浓度GA3(0、1×10-10、1×10-9、1×10-8、1×10-7、1×10-6、1×10-5、1×10-4mol/L)和BL(0、1×10-10、1×10-9、1×10-8、1×10-7、1×10-6mol/L)的50%MS固体培养基培养,30℃下持续光照培养1或2周(GA3为1周,BR处理2周)后,测量每株幼苗第2叶叶鞘长度。每个浓度梯度设置3个重复。
1.3 基因定位与候选基因分析
由《社会管理和公共服务综合标准化试点细则(试行)》② 2013年7月,标准委、教育部等25部委印发《社会管理和公共服务综合标准化试点细则(试行)》,细则中第二条对“社会管理和公共服务综合标准化试点”的定义作了详细解释。引申得出,社会管理和公共服务综合标准化是采用综合标准化方法,开展以制定标准、组织实施标准、对标准实施进行监督为主要内容,以实现管理规范、服务质量良好、公众满意度高为目标,促进提高社会管理科学化水平、推动基本公共服务均等化、加强保障和改善民生的活动。
从潇湘矮和NIL(NIP)衍生的F2群体中挑选出矮秆小粒的隐性单株,提取其DNA。首先分析潇湘矮与NIL(NIP)之间的多态性SSR标记,之后利用22株F2矮秆小粒的隐性单株调查每个多态性标记与潇湘矮矮秆小粒性状之间的连锁关系,对突变体基因进行初步定位。最后以786株F2矮秆小粒单株为材料,用新开发的6对SSR或InDel多态性标记对矮秆小粒基因进行精细定位(表1)。利用水稻基因组注释数据库(Rice Genome Annotation Project)对精细定位区间进行基因预测,并通过对候选基因进行序列分析,最终确定潇湘矮矮秆小粒候选基因。
1.4 载体构建与转基因
以PCR引物lv-1(序列:TTTGGATCC ATG GGCTCATCCTGTAGCAG,TTTGTCGACTCAAG TTCCTTCCCTGGAGC)从NIL(NIP)中扩增潇湘矮矮秆小粒候选基因完整基因组序列,利用限制线内切酶BamHⅠ和SalⅠ将其克隆到双元表达载体pCUbi1390载体上。利用农秆菌介导法将候选基因(启动子为玉米Ubi启动子)转入潇湘矮中。水稻愈伤组织诱导、分化,转化、筛选等方法参见文献[24]。

表1 本研究中基因定位所用引物Table 1.Primers for mapping in the study.
1.5 RNA提取与荧光定量PCR
分别以3周大小的潇湘矮以及互补转基因家系幼苗为材料,利用提取RNA试剂盒(北京鼎国昌盛生物技术有限公司)提取地上部分总RNA,用Primer premier 5设计D1定量PCR引物,前引物为ATTG ATGGCAGGTTGGATTA,后引物为GTTTCCTGA ATGGCTGGGTC;内参Ubquitin定量PCR引物,前引物为F(CTTGGTCGTGTCCCGT),后引物为R(TCCATGCTGCTCTACCAC)。实验仪器为高通量实时荧光定量PCR系统LightCycler®480Ⅱ,通过特定的荧光定量PCR产物的溶解曲线的单峰来判定引物的特异性,适合进行定量PCR。
2 结果与分析
2.1 潇湘矮表型以及形态学分析
潇湘矮株高从苗期开始就明显矮于其近等基因系,成熟后其株高约为NIL(NIP)的60%,同时穗长变短,穗型紧凑,粒型为小圆粒型(图1-A~E)。另外,潇湘矮的结实率、千粒重、粒长相比其近等基因系也显著下降,其中千粒重约为NIL(NIP)的45%,而有效穗数、粒宽等与NIL(NIP)差异不明显(图1-F~J)。比较植株各节间长度发现,相对于近等基因系,XXA植株主茎的第1、2和3节间及穗长都显著短于NIL(NIP)(图2-A、B)。
2.2 潇湘矮对GA3、BR敏感性分析
水稻株高的调控多与植物激素如赤霉素、油菜素内酯等的合成、代谢有关[5]。我们对潇湘矮及其NIL(NIP)设置了GA3和BR的梯度浓度处理(图3)。结果显示,施加外源BR后,潇湘矮与NIL(NIP)的第2叶叶鞘长度并没有明显改变(图3-B),说明调控潇湘矮株高的基因可能与BR合成无关。不同于BR处理,潇湘矮和NIL(NIP)的株高对GA3具有敏感性,随着GA3浓度的逐渐上升,它们第2叶叶鞘长度逐渐增加,虽然潇湘矮第2叶叶鞘的长度始终短于NIL(NIP),但是它们对GA3的响应速率基本一致(图3-A)。
2.3 潇湘矮矮秆小粒的遗传学分析及基因定位
我们将潇湘矮与半矮秆表型的日本晴、热研1号、0248、Y01杂交获得F1组合,结果显示所有F1组合植株都表现为半矮秆正常株型,其衍生的4个F2群体中正常半矮秆与类似潇湘矮的矮秆小粒株型分离,比例符合3∶1(表2)。因此,我们推测相对于日本晴等半矮秆水稻品种,潇湘矮发生了单基因突变,其矮秆小粒表型符合单隐性核基因控制的遗传规律,并将该突变体基因暂命名为xxa。同时,通过连续回交及杂交,我们构建了株型表现为正常半矮秆植株的近等基因系NIL(NIP)。对从潇湘矮和NIL(NIP)衍生的F2群体中挑选出的22株矮化小粒突变体单株进行连锁分析,发现矮秆小粒性状与第5染色体长臂上SSR标记RM182402、RM18457存在连锁。利用从F2中选出的96株矮化小粒单株将突变体基因初步定位于RM18420和RM18450间约2 cM的区间内(图4-A)。通过加密标记,并从F2中共挑选786株矮化小粒单株,突变体基因xxa最终被定位在第5染色体的InDel标记F81和F82间约70 kb区间的物理距离内(图4-A)。利用国际水稻基因组注释数据库(Rice Genome Annotation Project)分析发现该区间包含8个开放阅读框(ORF,图4-A)。其中,第5个ORF被注释为株高基因D1(LOC_Os05g26890)。D1编码G蛋白的α亚基,参与依赖G蛋白的GA信号传导,该基因突变后导致植株矮化粒型变小[13-14]。对潇湘矮和NIL(NIP)的D1基因序列比对发现,在潇湘矮中D1的第5和12外显子分别有1个碱基的替换和3个

图1 潇湘矮(XXA)及其近等基因系NIL(NIP)的表型分析Fig.1.Phenotype analysis of XXA and NIL(NIP).

图2 潇湘矮与近等基因系NIL(NIP)主茎穗长、节间长度Fig.2.The internode and panicle length comparison in XXA and NIL(NIP).

图3 潇湘矮与NIL(NIP)幼苗期第2叶叶鞘对外源赤霉素(A)和油菜素内酯(B)的响应Fig.3.Elongation of the second leaf sheath in response to gibberellin(A)and brassinolide(B)in NIL(NIP)and XXA plants.

表2 日本晴、热研1号、0248、Y01和潇湘矮杂交组合F2株高的分离情况Table 2.Segregation of combination Nipponbare/XXA,Reyan 1/XXA,0248/XXA and Y01/XXA.

图4 矮秆小粒基因的精细定位与候选基因分析Fig.4.Fine mapping and positional cloning of the candidate gene.
碱基的缺失,其中第5外显子1个碱基的替换属于无义突变,并没有导致氨基酸的任何改变,而第12外显子3个碱基的缺失导致1个赖氨酸的缺失(图4-B)。因此,我们推测潇湘矮矮秆小粒性状是D1基因突变所致。
2.4 转基因互补验证
为了进一步验证潇湘矮矮秆小粒性状是D1基因突变所致,我们扩增NIL(NIP)基因组中的D1基因序列,构建互补载体pCUbi1390-D1,通过农秆菌介导法导入潇湘矮中,让NIL(NIP)的D1基因在Ubi启动子的驱动下在潇湘矮中表达。对转基因T0阳性家系(D1/XXA)中D1基因表达水平分析发现,D1转基因和NIL(NIP)表达水平显著高于潇湘矮。表型分析发现,相对于潇湘矮,转基因阳性家系D1/XXA-1表现为株高变高,穗型变大,穗长及第Ⅰ、Ⅱ、Ⅲ节间长度显著变长,粒长显著增加,粒宽无明显变化(图5)。说明潇湘矮矮秆小粒性状是D1基因突变所致。
3 讨论
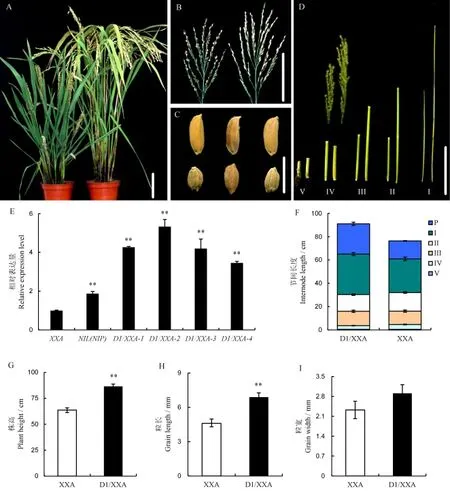
图5 转基因功能互补Fig.5.Phenotype analysis of complementary plants(D1/XXA).
本研究通过对育种过程中自然突变的矮秆小粒材料潇湘矮进行了形态学以及遗传学分析,发现了潇湘矮矮秆小粒性状符合单隐性基因控制的遗传规律。通过构建近等基因系及定位群体,对矮秆小粒基因进行了图位克隆。结果表明潇湘矮矮秆小粒性状是D1基因突变所致。水稻中最早报道的矮秆小粒突变体d1正是Dwarf 1(D1)基因突变所致。迄今,水稻报道的d1突变体有20多种,主要由大片段缺失、单碱基的突变、移码突变导致翻译提前终止等[5,25]。本研究中新发现的潇湘矮为d1的新等位突变体(第12个外显子处有3个碱基缺失导致一个1个赖氨酸的缺失)。生物信息学分析表明,潇湘矮突变位点正好位于G蛋白α亚基(G_alpha)功能域,可能影响G蛋白偶联受体的信号传导,最终导致矮秆圆小粒表型。D1的功能丧失除了导致水稻的矮化外,还伴有其他性状,如:茎秆增粗;叶片短且宽,叶色暗绿;穗形直立,着粒紧密,谷粒小而圆;节间密集,有时第2节间不伸长等[25-28]。我们发现潇湘矮株高明显矮于其近等基因系NIL(NIP),粒型小圆,穗型变小,且第2节间长度明显短于NIL(NIP)(图2)。前人研究表明d1矮秆突变体对外源赤霉素刺激不敏感,即外源赤霉素不能诱导其株高恢复正常[25]。本研究却显示潇湘矮对GA3表现出一定程度的敏感性,施加外源GA3可以部分恢复潇湘矮的株高(图3),该结果表明除D1外,可能还存在其株高相关基因(如半矮秆基因等)共同在调控潇湘矮的株高等表型。前人的研究推测控制潇湘矮矮秆小粒性状的可能有2对隐性基因,其中1对为已知的半矮秆基因sd1,且存在一个未知的半矮秆基因[23]。本研究中日本晴和潇湘矮的背景亲本均为半矮秆材料,它们均含有半矮秆基因sd1。因此,在半矮秆遗传背景下得出结论潇湘矮的矮秆性状主要由d1控制。而前者提到的控制潇湘矮矮秆小圆粒的未知基因很有可能就是d1。此外,最新研究发现D1可能参与水稻干旱胁迫调控途径,D1新的等位突变体rga1对干旱胁迫的敏感性降低,干旱处理14 d后,叶片仍为暗绿色且直立,而野生型叶片黄化并萎蔫[29]。因此,D1的突变体rga1在培育抗旱水稻的分子育种中具有重要应用价值。
本研究中稳定遗传矮秆小粒水稻潇湘矮,其籽粒很小,千粒重只有12 g左右,因此,我们可以将其导入不育系中,杂交制种时小粒不育系与恢复系混播、混种、混收,最后通过机械筛选去除大粒恢复系种子留下小粒杂交种子,而小粒性状为隐性核基因控制,杂种F1所结种子粒型正常,不影响产量。因此利用潇湘矮的矮秆小粒基因可以实现水稻杂交制种全程机械化,具有重要的实践意义。
[1]Khush G S.Green revolution:the way forward.Nat Rev Genet,2001,2(10):815-822.
[2]Hu C H.Evaluation of breeding semidwarf rice by induced mutation and hybridization.Euphytica,1973,22: 562-574.
[3]Sasaki A,Ashikari M,Ueguchi-Tanaka M,Itoh H, Nishimura A,Swapan D,Ishiyama K,Saito T,Kobayashi M,Khush G S,Kitano H,Matsuoka M.Green revolution: A mutant gibberellin-synthesis gene in rice.Nature,2002, 416:701-702.
[4]Peng J,Richards D E,Hartley N M,Murphy G P,Devos K M,Flintham J E,Beales J,Fish L J,Worland A J, Pelica F,Sudhakar D,Christou P,Snape J W,Gale M D, Harberd N P.“Green revolution”genes encode mutant gibberellin response modulators.Nature,1999,400: 256-261.
[5]张云辉,张所兵,林静,汪迎节,方先文.水稻株高基因克隆及功能分析的研究进展.中国农学通报,2014, 30:1-7. Zhang Y H,Zhang S B,Fang X W,Lin J,Wang Y J, Fang X W.Research progress on cloning and functional analysis of plant height genes in rice(Oryza sativa L.). Chin Agric Sci Bull,2014,30:1-7.(in Chinese with English abstract)
[6]Fujioka S,Yokota T.Biosynthesis and metabolism of brassinosteroids.Annu Rev Plant Biol,2003,54:137-164.
[7]ItohH,Ueguchi-TanakaM,Sato Y,AshikariM, Matsuoka M.The gibberellin signaling pathway is regulatedbytheappearanceanddisappearanceof SLENDER RICE1 in nuclei.Plant Cell,2002,14:57-70.
[8]Hirano K1,Asano K,Tsuji H,Kawamura M,Mori H, KitanoH,Ueguchi-TanakaM,MatsuokaM. Characterization of the molecular mechanism underlying gibberellin perception complex formation in rice.Plant Cell,2010,22:2680-2696.
[9]Smith S M,Waters M T.Strigolactones:Destructiondependent perception?Curr Biol,2012,22:924-927.
[10]Monna L1,Kitazawa N,Yoshino R,Suzuki J,Masuda H, Maehara Y,Tanji M,Sato M,Nasu S,Minobe Y. Positional cloning of rice semidwarfing gene,sd-1:Rice“green revolution gene”encodes a mutant enzyme involved in gibberellin synthesis.DNA Res,2002,9: 11-17.
[11]SpielmeyerW,EllisMH,ChandlerPM. Semidwarf(sd-1),“green revolution”rice,contains a defective gibberellin 20-oxidase gene.Proc Natl Acad Sci USA,2002,99:9043-9048.
[12]Ueguchi-Tanaka M,Ashikari M,Nakajima M,Itoh H, Katoh E,Kobayashi M,Chow T Y,Hsing Y I,Kitano H, YamaguchiI,MatsuokaM.GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin.Nature,2005,437:693-698.
[13]Izawa Y,Takayanagi Y,Inaba N,Abe Y,Minami M, Fujisawa Y,Kato H,Ohki S,Kitano H,Iwasaki Y. Function and expression pattern of the alpha subunit of the heterotrimeric G protein in rice.Plant Cell Physiol, 2010,51:271-281.
[14]Kamei T,Matozaki T,Sakisaka T.Coendocytosis of cadherin and c-Met coupled to disruption of cell-cell adhesion in MDCK cells-regulation by Rho,Rac and Rab small G proteins.Oncogene,1999,18:6776-6784.
[15]Wu Y,Fu Y,Zhao S,Gu P,Zhu Z,Sun C,Tan L. CLUSTERED PRIMARY BRANCH 1,a new allele of DWARF11,controls panicle architecture and seed size in rice.Plant Biotechnol J,2016,14:377-386.
[16]Tong H,Liu L,Jin Y,Du L,Yin Y,Qian Q,Zhu L,Chu C.DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice.Plant Cell, 2012,24:2562-2577.
[17]Shi Z.Characterization and cloning of SMALL GRAIN 4, a novel DWARF11 allele that affects brassinosteroid biosynthesis in rice.Chin Sci Bull,2015,60:905-915.
[18]Zhou F,Lin Q,Zhu L,Ren Y,Zhou K,Shabek N,Wu F, Mao H,Dong W,Gan L,Ma W,Gao H,Chen J,Yang C, Wang D,Tan J,Zhang X,Guo X,Wang J,Jiang L,Liu X, Chen W,Chu J,Yan C,Ueno K,Ito S,Asami T,Cheng Z, Wang J,Lei C,Zhai H,Wu C,Wang H,Zheng N,Wan J. D14-SCF(D3)-dependent degradation of D53 regulates strigolactonesignalling.Nature,2013,504(7480): 406-410.
[19]Jiang L,Liu X,Xiong G,Liu H,Chen F,Wang L,Meng X,Liu G,Yu H,Yuan Y,Yi W,Zhao L,Ma H,He Y, Wu Z,Melcher K,Qian Q,Xu H E,Wang Y,Li J. DWARF 53 acts as a repressor of strigolactone signalling in rice.Nature,2013,504:401-405.
[20]Huang N,Courtois B,Khush G S.Association of quantitative trait loci for plant height with major dwarfing genes in rice.Heredity,1996,77:130-137.
[21]Moncada P,Martínez C P,Borrero J.Quantitative trait loci for yield and yield components in an Oryza sativa× Oryza rufipogon BC2F2population evaluated in an upland environment.Theor Appl Genet,2002,102:41-52.
[22]罗炬,邵高能,魏祥进,陈明亮,唐绍清,焦桂爱,谢黎虹,胡培松.一个控制水稻株高QTL qPH3的遗传分析.中国水稻科学,2012,26(4):417-422. Luo J,Shao G N,Wei X J,Chen M L,Tang S Q,Jiao Q A,Xie L H,Hu P S.Genetic analysis of a QTL qPH3 for plant height in rice.Chin J Rice Sci,2012,26(4):417-422. (in Chinese with English abstract)
[23]余应弘,吴云天,曾翔,袁隆平.水稻矮秆小粒突变体潇湘矮的特征特性及其遗传鉴定.杂交水稻,2007, 22(6):67-70. Yu Y H,Wu Y T,Zeng X,Yuan L P.Characterization and genetic studies on dwarf rice mutant Xiaoxiangai with small grains.Hybrid Rice,2007,22(6):67-70.(in Chinese with English abstract)
[24]郑贵朝,胡事君.提高水稻愈伤组织植株再生能力几种方法的评价.杂交水稻,2005,20(2):54-57. Zheng C G,Hu S J.Evaluation of some culture methods for enhancing plant regeneration ability of rice callus. Hybrid Rice,2005,20(2):54-57.(in Chinese with English abstract)
[25]Ashikari M,Wu J,Yano M,Sasaki T,Yoshimura A.Rice gibberellin-insensitivedwarfmutantgeneDwarf1 encodes the alpha-subunit of GTP-binding protein.Proc Natl Acad Sci USA,1999,96:10284-10289.
[26]Ueguchi-Tanaka M,Fujisawa Y,Kobayashi M.Rice dwarf mutant d1,which is defective in the alpha subunit of the heterotrimeric G protein,affects gibberellin signal transduction.Proc Natl Acad Sci USA,2000,97: 11638-11643.
[27]Hartweck L M,Olszewski N E.Rice GIBBERELLIN INSENSITIVE DWARF1 is a gibberellin receptor that illuminates and raises questions about GA signaling. Plant Cell,2006,18:278-282.
[28]Choe S,Dilkes B P,Gregory B D,Ross A S,Yuan H, Noguchi T,Fujioka S,Takatsuto S,Tanaka A,Yoshida S, Tax F E,Feldmann K A.The Arabidopsis dwarf 1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis.Plant Physiol,1999,119:897-907.
[29]Ferrero-Serrano A,Assmann S M.The alpha-subunit of the rice heterotrimericG protein,RGA1,regulates drought tolerance during the vegetative phase in the dwarfricemutantd1.JExpBot,2016,67(11): 3433-3443.
Morphological and Molecular Genetic Analysis of a Dwarf and Small Grain Rice Variety Xiaoxiang’ai
LÜ Yusong1,#,XIE Yunfeng1,2,#,SHENG Zhonghua1,WU Yawen1,TANG Shaoqing1,HU Peisong1, WEI Xiangjin1,*
(1State Key Laboratory of Rice Biology/Key Laboratory of Rice Biology and Breeding of Ministry of Agriculture,China National Rice Research Institute, Hangzhou 310006,China;2College of Life and Environmental Sciences,Hangzhou Normal University,Hangzhou 310036,China;#These authors contributed equally to this work;*Corresponding author,E-mail:weixiangjin@caas.cn)
【Objective】To uncover the genetic mechanism of a dwarf and small grain variety Xiaoxiang’ai(XXA),and lay an important theoretical foundation in rice dwarfing breeding.【Method】Rice variety XXA characterized as a dwarf and small grain phenotype was derived from natural mutation under breeding process.The agronomic traits of XXA were investigated.Its sensibility to phytohormone was analyzed under GA3and BR treatments at various concentrations. Genetic analysis of XXA with different varieties and fine mapping of xxa gene from the F2population derived from the cross between near-isogenic lines NIL(NIP)and XXA were done.Furthermore,complementary assay was performed to confirm the candidate gene.【Result】The XXA shows short spikes and compact panicles,with significant decrease in 1000-grain weight under GA3and BR treatments,we found that XXA is partially sensitive to GA3,whereas insensitive to BR.The segregation behavior in each of the derived F2populations between XXA and either varieties was consistent with the Mendelian monogenic ratio of 3∶1.The xxa gene was finally narrowed down to a 70 kb physical region between the InDel markers F81 and F82 on chromosome 5.Within this region,eight open reading frames(ORFs)were predicted, of which,ORF5(LOC_Os05g26890)has been annotated as a Dwarf1 gene(D1).Furthermore,sequence analysis of all the region revealed that there are only two mutation sites including one 1-bp nonsense mutation in the 5th exon and a 3-bp deletion that resulted in a lysine deletion in the 12th exon of D1.The complementary assay showed that the abnormal phenotype of XXA can be recovered.【Conclusion】The dwarf and small grain phenotype is probably associated with GA3metabolism.The abnormal phenotype is mainly controlled by a single recessive gene.The D1 indisputably corresponds to mutation phenotype of XXA.
rice;dwarf and small grain;gene mapping;mechanized propagation
S511.024献标识码:A
:1001-7216(2017)03-0238-09
2016-11-24;修改稿收到日期:2017-02-06。
国家自然科学基金资助项目(31471472);国家重点研发计划资助项目(2016YFD0101801);浙江省自然科学基金资助项目(LY14C30009)。
